
潞江坝干热河谷区5 种常见作物田的昆虫群落特征*
2023-08-03 06:37:34李勇辉周天江陈文华唐国文
云南农业大学学报(自然科学) 2023年3期
李勇辉,周天江,陈文华,唐国文 ,柳 青
(1.云南农业大学 植物保护学院,云南生物资源保护与利用国家重点实验室,云南 昆明 650201;2.保山学院 资源环境学院,云南省高校滇西昆虫资源保护与利用重点实验室,云南 保山 678000)
农田生态系统是一种人为干预和控制下形成的人工生态系统[1],其服务功能是以最大限度地提供粮食等优质农产品为核心[2]。随着农业耕作模式向集约化和规模化的转变,在农业管理过程中大量使用化学农药,造成农田害虫抗药性增强、天敌昆虫的非靶标作用和生物多样性丧失等问题越来越严重[3-4],进一步降低了农田生态系统的自我调节能力,加剧了植食性害虫的频繁暴发,严重影响农产品的产量和质量[5]。
近年来,通过改变农作物的种植模式来改善农田生物多样性和农田生态系统保益控害的服务功能[6],实现对作物害虫的生态调控,已逐渐成为国内外学者研究的热点[7-8]。已有研究证实:作物多样性种植可减少农田害虫的数量、减轻其危害程度[9],如间作番茄显著降低了花椰菜田中菜蚜(Lipaphis erysimi)的个体数量,且控制效果随番茄间作比例的上升而增强[10];玉米间作大豆能明显提高农田节肢动物群落的物种数、物种丰富度指数、均匀度指数、多样性指数和天敌物种数[11-12];与单作玉米田相比,邻作模式下玉米田节肢动物的物种丰富度指数、多样性指数和均匀度指数均明显增加[13]。
云南省保山市潞江坝是中国典型的亚热带干热河谷之一,具有光照充足、全年无霜、干湿季分明、降水少和蒸发量大等气候特点[14],也是保山市重要的热区农作物种植区,主要种植咖啡、芒果、火龙果、龙眼、荔枝、香蕉和蔬菜等多种热带经济作物。随着近年来种植面积的不断扩大,各种农田害虫发生种类及危害程度呈逐年增加的趋势[15]。目前,利用农田生物多样性控制虫害的研究已在小麦、烟草和水稻等多种作物上取得成效,而在其他作物上的研究还较少[16-17],尤其是对干热河谷地区农作物害虫及天敌群落的研究还比较缺乏。因此,本研究选取潞江坝干热河谷区5 种常见作物田进行害虫及天敌群落调查,以明确该地区不同作物田害虫及天敌群落的结构和特征,为干热河谷区农田害虫防控实践提供参考依据。
1 材料与方法
1.1 调查样地概况
调查样地位于云南省保山市隆阳区潞江镇顿东村(N24°54′46″,E98°52′06″,海拔760 m),年平均气温25 ℃,年降水量1 000 mm。结合当地作物种植情况,选取5 种常见作物田进行调查:黄瓜地,面积约0.30 hm2,种植品种为津优11 号,周围邻作菜豆;番茄地,面积约0.40 hm2,种植品种为德澳特7845,周围邻作黄瓜;菜豆地,面积约1.20 hm2,种植品种为泰国架豆,周围邻作少量咖啡;芒果地,面积为0.80 hm2,种植品种为鹰嘴芒,树高约5 m,周围零星种植保山小粒咖啡(树高约1 m),样地管理粗放,分布多种杂草及其他小型灌木;青枣地,面积约0.35 hm2,种植品种为台南,树高约2 m,样地内有较多杂草及小型灌木,周边分布有少量芒果(树高约5 m)。5 个样地间隔均大于100 m,调查期间各样地均按照当地常规措施管理,农药使用频率约为每周1 次。
1.2 研究方法
1.2.1 调查方法
调查期为2020 年3—11 月,每隔15 d 采用目测观察法、扫网法和诱集法调查1 次。
目测观察法:采用五点取样法,每点随机选择一定数量的作物或果树(蔬菜:1 m2范围内,果树:相邻2 株),仔细观察并记录植株以及地面上的昆虫种类和数量。
扫网法:在每个样地的5 个取样点进行扫网采集,每2 个调查点之间随机扫网10 次,共计40 网,记录入网的昆虫种类和数量。
诱集法:每个样地按五点取样法在2 个对角调查点各悬挂黄板(30 cm×20 cm) 1 块,悬挂高度比作物高15 cm,另外2 个对角调查点各放置黄盘(圆形,直径15 cm×高2 cm) 1 个,中心调查点放置1 个装有引诱剂500 mL 的诱集瓶(圆柱形,直径10 cm×高15 cm),引诱剂为糖、醋、56%白酒和水的混合物,各成分的质量比为3∶4∶1∶2[13]。每次诱集24 h,收集并记录诱集到的昆虫种类和数量。
在田间不能鉴定的昆虫标本保存于75% 乙醇溶液中,带回实验室鉴定。鳞翅目和蜻蜓目昆虫标本用乙酸乙酯毒瓶毒死后装入三角纸袋。
1.2.2 数据分析方法
用Margalef 丰富度指数、Simpson 优势集中性指数、Shannon-Wiener 多样性指数、Pielou 均匀度指数和chao1 指数分析害虫和天敌的群落多样性[18-20]。其中chao1 指数也是生态学中度量物种丰富度的指标,可根据观测到的结果推算理论丰富度,且该丰富度更接近真实值[20]。以上所有指数均在R 4.2.0 软件中计算。
采用R 语言软件中的iNEXT 软件包绘制5 种作物田的物种稀疏及预测曲线,分析其抽样充分性。群落稳定性采用Sa/Sb值 (Sa为天敌个体数,Sb为害虫个体数)和ds/dm值 (ds为多样性指数标准差,dm为多样性指数平均值)进行描述[21]。
群落相似性采用 Jaccard 指数(Cj)计算[22]:Cj=c/(a+b-c)。式中,c为两样地共有的物种数;a和b分别为样地 A 和样地 B 的物种数。当 0≤Cj<0.25 时,群落极不相似;当 0.25≤Cj<0.50时,群落中等不相似;当 0.50≤Cj<0.75 时,群落中等相似;当 0.75≤Cj≤1.00 时,群落极相似。利用R 4.2.0 软件中的iNEXT 和vegan 包,采用95%置信区间的200 次Bootstrap 重抽样方法进行计算[23]。各样地群落特征指数差异采用Duncan’s 新复极差法进行显著性检验,显著性水平为P<0.05。使用Excel 2019 和R 4.2.0 软件中的Venn-Diagram 绘图。
2 结果与分析
2.1 5 种常见作物田的昆虫群落组成
调查共采集农田昆虫6 307 头,隶属12 目64 科113 种,其中害虫亚群落72 种,占物种总数的63.72%,隶属8 目42 科;天敌亚群落41种,占物种总数的36.28%,隶属8 目22 科。由表1 可知:芒果地的昆虫物种数和个体数最多,有12 目47 科66 种2 334 头,分别占昆虫总物种数的57.89%和总个体数的37.01%;其次为青枣地,有9 目39 科50 种1 344 头,占总物种数的43.86%和总个体数的21.31%;黄瓜地、番茄地和菜豆地的昆虫物种数和个体数均较少,其中黄瓜地的物种数最少(20 种),占总物种数的17.54%;菜豆地昆虫个体数最少(796 头),占总个体数的12.62%。

表1 5 种常见作物田的昆虫群落组成Tab.1 Insect community composition in five common crop fields
5 种作物田中,害虫亚群落物种数以芒果地和青枣地最丰富,分别为41 和36 种,各占害虫物种总数的56.94%和50.00%,其他3 种作物田的害虫物种数为菜豆地(24 种)>番茄地(15 种)>黄瓜地(12 种);害虫个体数为芒果地(1 837 头)>青枣地(1 186 头)>黄瓜地(858 头)>番茄地(854 头)>菜豆地(739 头);害虫chao1 指数为芒果地(76.00)>青枣地(49.20)>菜豆地(45.00)>黄瓜地(22.00)>番茄地(18.75)。5 种作物田的害虫优势种较相似,主要有黑腹果蝇 (Drosophila melanogaster)和棉露尾甲(Haptonchus luteolus)。
5 种作物田中,天敌亚群落物种数为芒果地(25 种)>青 枣地(14 种)>菜豆 地(11 种)=番 茄地(11 种)>黄瓜地(8 种);天敌个体数为芒果地(497 头)>青枣地(158 头)>番茄地(84 头)>菜豆地(57 头)>黄瓜地(37 头)。天敌chao1 指数为芒果地(36.14)>青枣地(15.67)>黄瓜地(14.00)>番茄地(13.00)>菜豆地(12.20)。5 种样地的天敌优势种主要包括圆腹异突颜寄蝇(Ectophasia rotundiventris)、日本蠼螋(Labidura japonica)、鳃佩雷寄蝇(Peleteria semiglabra)、黄猄蚁(Oecophylla smaragdina)和蠼膄(Forficula auricularia)。
由图1 可知:5 种作物田共有害虫4 种,各作物田特有的害虫物种数由高到低依次为青枣地(15 种)>芒果地(13 种)>菜豆地(7 种)>黄瓜地(4 种)>番茄地(3 种);5 种作物田共有天敌3 种,各作物田特有的天敌物种数由高到低依次为芒果地(13 种)>青枣地(5 种)=菜豆地(5 种)>番茄地(3 种)>黄瓜地(2 种)。

图1 5 种常见作物田的害虫(a)和天敌(b)物种集合韦恩图Fig.1 Venn diagram of the pests (a) and natural enemies (b) in five common crop fields
2.2 5 种常见作物田的昆虫群落特征
2.2.1 抽样充分性
抽样性分析显示:随着昆虫个体数的增加,芒果地的物种数也逐渐增加,表明该地区的昆虫群落抽样不充分;而黄瓜地、番茄地、菜豆地和青枣地的物种数随着昆虫个体数的增加而逐渐趋于稳定,说明这4 种作物田的昆虫群落抽样较充分(图2)。

图2 5 种常见作物田的昆虫群落稀疏—外推曲线Fig.2 Sparse extrapolation curves of the insect communities in five common crop fields
2.2.2 多样性
由表2 可知:昆虫群落的物种丰富度指数和多样性指数为芒果地>青枣地>菜豆地>番茄地>黄瓜地,优势集中性指数和均匀度指数为芒果地>青枣地>黄瓜地>番茄地>菜豆地,其中芒果地的昆虫群落物种丰富度指数、多样性指数、优势集中性指数和均匀度指数均显著高于其他作物田(P<0.05)。

表2 5 种常见作物田的昆虫群落多样性特征Tab.2 Characteristics of insect communities diversity in five common crop fields
害虫亚群落多样性分析(图3)显示:物种丰富度指数为芒果地>青枣地>菜豆地>番茄地>黄瓜地,多样性指数为芒果地>青枣地>菜豆地>黄瓜地>番茄地,且芒果地和青枣地均显著高于其他3 种作物田(P<0.05);优势集中性指数为青枣地>芒果地>黄瓜地>番茄地>菜豆地,其中青枣地显著高于其他作物田(P<0.05);均匀度指数为芒果地>青枣地>黄瓜地>番茄地>菜豆地,其中芒果地显著高于其他4 种作物田(P<0.05)。

图3 5 种常见作物田的害虫和天敌亚群落特征指数Fig.3 Characteristic indices of sub-community of pests and natural enemies in five common crop fields
天敌亚群落多样性分析(图3)显示:物种丰富度指数为芒果地>青枣地>菜豆地=番茄地>黄瓜地,其中芒果地显著高于其他作物田(P<0.05);多样性指数和均匀度指数表现为黄瓜地>菜豆地>番茄地>青枣地>芒果地,且青枣地和芒果地显著低于其他作物田(P<0.05);优势集中性指数表现为黄瓜地>番茄地>菜豆地>青枣地>芒果地,其中黄瓜地显著高于其他作物田(P<0.05)。
2.3 5 种常见作物田的昆虫群落稳定性
由表3 可知:5 种常见作物田的昆虫群落Sa/Sb值为芒果地>青枣地>番茄地>菜豆地>黄瓜地;ds/dm值为菜豆地>番茄地>黄瓜地>青枣地>芒果地。可见,在人为干扰较小的芒果地和青枣地有较多的天敌个体数,且多样性指数的变异系数较小,说明群落内天敌占比较大,食物网关系的复杂性及相互制约程度较大,群落的稳定性也相对较强。

表3 5 种常见作物田的昆虫群落稳定性分析Tab.3 Stability analysis of insect community in five common crop fields
2.4 5 种常见作物田的昆虫群落物种相似性
由表4 可知:5 种常见作物田的昆虫群落物种相似度系数均小于0.5,表现为中等不相似或极不相似。其中,黄瓜地和菜豆地的相似度最低(0.170 2),表现为极不相似;黄瓜地和番茄地的相似度最高(0.352 9),芒果地和青枣地的相似度为0.303 4,表现为中等不相似;其余各样地间的相似度小于0.25,表现为极不相似。
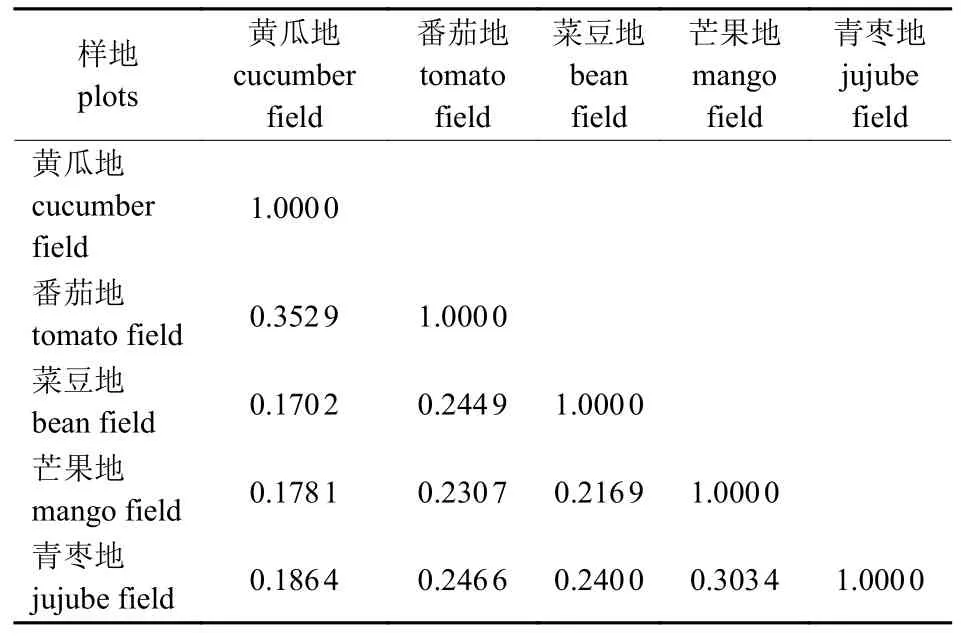
表4 5 种常见作物田的昆虫群落相似性分析Tab.4 Similarity analysis of insect communities in five common crop fields
3 讨论
农业实践的集约化经营极大降低了农田节肢动物的多样性[24-26],导致农田虫害的发生越来越严重[27-28]。干热河谷区具有十分独特的气候条件,植被覆盖度低,水热矛盾突出,加之农业活动频繁,生态环境较为脆弱,各种病虫害极易暴发成灾,对农田作物的影响较大[14]。因此,研究干热河谷区作物田的害虫和天敌群落特征对特殊生境条件下农田害虫防控具有借鉴意义。
本研究对云南省保山市潞江坝干热河谷区5 种常见作物田的昆虫群落进行调查,共采集昆虫6 307 头,隶属12 目64 科113 种,反映出该地区昆虫群落丰富,其中害虫8 目42 科72 种,天敌8 目22 科41 种。5 种样地中,以芒果地的昆虫物种数和个体数最多,青枣地次之,黄瓜地、番茄地和菜豆地的昆虫物种数和个体数均较少;害虫亚群落和天敌亚群落组成与总昆虫群落表现出相似规律,物种数、个体数和chao1 指数均以芒果地和青枣地最高,这与SPERBER 等[29]对巴西可可农林复合生态系统树种丰富度和密度对膜翅目寄生蜂多样性的影响研究结果较为吻合。本研究中芒果地和青枣地管理粗糙,样地中植被覆盖度高,适宜各种昆虫的生存和繁衍,而黄瓜地、番茄地和菜豆地管理精细,植被类型非常单一,不利于昆虫生存。此外,本研究还发现5 种样地的害虫优势种较相似,天敌优势种则存在一定差异,出现这种情况的原因可能与调查区域种植的作物类型有关,5 种样地种植作物以蔬菜和水果等多肉植物为主,加之该地区光热资源丰富,一年四季均可种植,且作物种类相对固定,使各样地害虫优势种表现出一定相似性,而天敌优势种由于受各样地害虫种类的不同而表现出差异性。
物种多样性可以反映群落基本特征,是判断群落发展阶段的重要尺度[30-33]。本研究中,5 种样地昆虫总群落及害虫亚群落物种丰富度指数、多样性指数和均匀度指数均为芒果地最高,青枣地次之,其余3 种样地则较低,其原因可能是黄瓜地、番茄地和菜豆地受人为因素的影响较大,而芒果地和青枣地管理较为粗放,人为干扰较小,且样地中有多种杂草及其他灌木,不仅增加了昆虫的物种数,也为昆虫个体数的发生提供了条件。天敌亚群落多样性与害虫亚群落完全不同,除物种丰富度指数外,芒果地和青枣地的多样性指数、优势集中性指数和均匀度指数均显著低于其他3 种样地,这一结果与陈芳等[34]对新疆六师农垦区不同生境条件下节肢动物群落多样性的研究结果类似。已有研究认为:农田作物物种种类及丰富度可以显著影响田间昆虫发生的种类和数量,但对害虫的影响更为直接,对天敌的影响甚微,天敌发生情况更多受害虫物种丰富度的影响[35]。本研究中,芒果地和青枣地天敌的物种丰富度较高而其他3 个指标较低,可能受各样地害虫发生情况及该区域特殊气候条件等多种因素的影响[36]。此外,本研究中5 种样地的昆虫总群落优势集中性指数和均匀度指数表现出一致的变化规律,这与前人对云南玉溪烟田害虫和天敌资源[37]以及对安徽菜子湖湿地鞘翅目昆虫区系和多样性研究[38]的结果相似,分析其原因可能与5 种样地的昆虫群落组成有关。5 种样地具有不同的昆虫物种组成,但表现出相似的害虫和天敌优势种种类,且发生数量较大,在一定程度上影响了各样地昆虫群落的均匀度。
Sa/Sb值是反映昆虫群落稳定性的重要指标,比值越大,稳定性越强[13];而ds/dm则相反,比值越小,群落稳定性越好[20]。本研究中,Sa/Sb值表现为芒果地最大,青枣地次之,其他3 种样地最小;ds/dm值则表现为芒果地和青枣地最小,其他 3 种样地最大,表明芒果地和青枣地的稳定性和抗干扰能力较其他3 种样地强。这与前人对不同种植环境夏玉米田节肢动物群落稳定性的研究[39]以及桃园生草对桃树节肢动物群落稳定性的研究[20]结果一致。群落结构相似性分析表明:5 种常见作物田的昆虫群落物种相似度系数均小于0.5,表现为中等不相似或极不相似,这可能与研究区种植的作物种类有关。5 种样地种植的作物是该地区常见的类型,但因种类和管理方式不同,直接导致各样地昆虫群落的差异。
4 结论
潞江坝干热河谷区气候条件复杂,其农田昆虫群落特征特殊,对该区域农田害虫的有效防控应注重保持农田植被较高的稳定性,选择抗虫的蔬菜和水果品种。结合本研究结果,建议尽可能采取间作和混作等多样化种植模式,提高农田作物多样性,实现害虫的绿色防控。
猜你喜欢
今日农业(2022年7期)2022-07-06 20:28:51
基层中医药(2021年4期)2021-07-22 07:15:32
——管氏肿腿蜂的应用技术
江苏林业科技(2020年3期)2020-08-31 01:00:08
现代园艺(2017年21期)2018-01-03 06:42:15
作文与考试·初中版(2017年23期)2017-08-14 10:43:50
文学少年(小学版)(2016年7期)2016-05-14 13:13:29
中国果业信息(2016年3期)2016-01-29 23:19:02
小天使·六年级语数英综合(2014年10期)2014-10-20 07:03:37
食品科学(2013年13期)2013-03-11 18:24:41
长江蔬菜(2013年23期)2013-03-11 15:09:39
