
盐渍土磷有效性研究进展与展望①
2023-08-02 02:53咸敬甜陈小兵张晓龙
土壤 2023年3期
咸敬甜,陈小兵,王 上,张晓龙,徐 刚
盐渍土磷有效性研究进展与展望①
咸敬甜1,2,陈小兵2*,王 上3,张晓龙1,徐 刚4
(1 烟台大学环境与材料工程学院,山东烟台 264005;2 中国科学院海岸带环境过程与生态修复重点实验室(烟台海岸带研究所),山东烟台 264003;3 德国基尔大学土壤植物微生物系,德国基尔 24118;4曲阜师范大学地理与旅游学院,山东日照 276826)
盐渍土约占据全球土地面积的3%,是具有重要战略意义的后备土地资源。然而,盐分胁迫不仅降低土壤水的渗透势,产生离子毒害作用,而且抑制植物营养的均衡吸收,对植物大量营养元素磷(P)的影响尤为严重,使其有效性大大降低,严重影响作物产量。因此,尽管有效磷缺乏是各土壤类型广泛存在的问题,但盐渍土中磷有效性低的问题更加突出。盐分胁迫和有效磷缺乏是两种主要的非生物胁迫,尤其对于钙质土壤,植物经常同时暴露在这两种胁迫之下,但现阶段关于盐分与磷素的作用效应缺乏清晰的界定和系统的总结。因此,本文首先汇总了盐分降低磷有效性的作用机制,其后综述了盐分与磷素的相互、综合、交互作用效应,最后总结了目前盐渍土磷有效性的提升策略并对其研究前景进行了展望,以期为未来盐渍土非生物胁迫的减轻、磷元素的优化管理提供理论支持。
盐渍土;磷有效性;作用机制;作用效应;提升策略
在全球范围内,由于人口激增、生态系统破坏和退化、气候变化等带来的粮食安全、资源短缺问题已迫在眉睫,继而盐渍土作为潜在的农用土地资源而备受关注。当前,全球盐渍土的总面积为1.1×109hm2,其中86% 可以用于农业生产,为全球提供11% 的作物产量[1],且全世界超过6% 的土地和20% 的灌溉面积正面临盐分威胁。我国盐渍土总面积达3.69× 107hm2[2]。盐渍化是半干旱和干旱地区的一个自然过程,但也受人类活动的影响,显著影响土地肥力[3]。除干旱和养分(尤其是磷)的限制外,土壤盐分胁迫是对作物生产力最具破坏性的非生物限制因素之一[4-5]。随着全球人口的增长,根据联合国粮食及农业组织(FAO)的数据,到2050年,要求粮食产量至少翻一番[6]。因此,盐渍土的改良及利用得到了高度重视。
磷对植物营养和土壤肥力都起着十分重要的作用,是光合作用、呼吸作用以及核酸和膜合成的关键元素[7-8]。但在大多数自然生态系统中全磷含量相对较低,一般在0.05% ~ 0.5%(/)左右,且其中只有0.1% 的生物有效磷被植物吸收利用[9]。土壤肥力退化,尤其是磷素耗竭,是限制植物生长发育的主要因素,据统计全世界约30% ~ 40% 可耕地上的作物产量受磷的限制[10]。磷缺乏是限制全球许多低投入农业系统作物产量的主要因素,包括许多石灰性和碱性土壤[11-12]。为提高农业生产力,大量磷肥施入土壤,但在施用当年作物仅能吸收磷肥施用量的15% ~ 30%[13]。施入土壤中的磷迅速转化成植物难以利用的形态,一般磷缺乏是指有效磷缺乏。据统计,在1998—2009年间粮食产量仅增加了10%,而化肥投入增加了近19%,增加的化肥并没有增加相应的粮食产量[14]。另外,磷肥主要来源于不可再生的磷矿石,按照目前每年近12 Mt P2O5的开采速度[15],全球商业磷酸盐储量将在50 ~ 100年内耗尽[6],而当利用磷矿石生产磷肥时,40% ~ 60% 的磷酸盐会流失和浪费[6],且目前还没有磷酸盐的替代品。因此,减少磷肥使用量,提高作物对磷的吸收和利用效率,是实现磷矿资源可持续利用、降低过度施用磷肥对环境的不利影响的关键[16]。现有研究显示,土壤中磷的有效性主要取决于土壤中黏土矿物含量、金属(Ca、Fe、Al)氧化物含量、pH、有机质含量等[17-20]。此外,土壤盐分胁迫也会降低磷的有效性[21-22]。
盐分胁迫和有效磷缺乏是两种主要的非生物胁迫,严重限制了全球植物生产、粮食安全和农业发展[23-25],对区域和全球经济产生了巨大影响,尤其是在以高pH、低磷溶解度为特征的钙质土壤中,植物常同时暴露在这两种非生物胁迫之下[26-27]。盐渍土中电解质浓度高,盐分离子富集,增强了磷的吸附过程,使磷的有效性降低,但因土壤质地不同而有所不同[28]。
近年来盐渍土中磷有效性的研究得以广泛开展,但现阶段对于盐分和磷素的相互、综合、交互作用效应及盐渍土磷素有效性提升策略缺乏系统的总结和探索。由此,本文汇总分析了盐分降低磷有效性的作用机制,盐分与磷素的各种作用效应,以期为盐渍土磷素有效性提升、盐渍土优化磷素管理及作物生产力提高提供较为全面的理论支持。
1 土壤盐渍化与磷素研究相关文献分析
以Web of Science核心数据集2000年1月1日— 2022年3月1日已发表的文献为统计分析对象,以“soil salinization”为关键词进行所有字段检索,获得相关文章共计2 626篇,将其全部导入VOSviewer进行文献可视化分析(图1A)。由于关键词数量众多,对关键词的出现频次进行了限定,筛选出频次高于50次的关键词,最后得到278个关键词,且分为6个聚类簇。选取其中4个总关联强度高的聚类簇分别标号:①聚类簇主要是“碳(carbon)”“有机物(organic- matter)”“氮(nitrogen)”“磷(phosphorus)”“动力学(dynamics)”等关键词。说明主要营养元素及物质的动力学过程备受关注。这些过程又依赖于根际微生物的生命活动,因此,根际微生物及酶活性也是现阶段研究的热点。但过度补充矿质养分(化肥)又会造成土壤次生盐渍化等问题,因而其负面影响的修复也是研究的重要方向,其中黄河三角洲地区(Yellow River Delta)作为典型案例是重点研究对象。②聚类簇主要是“土壤盐度(soil salinity)”“植被(vegetation)”“模型(model)”“遥感(remote sensing)”“水分(moisture)”等关键词。说明土壤盐度对植物的影响效应是研究的主体。由于水盐的监测都依赖于一定的模型和技术方法,因此,利用遥感、光谱学及模型等来绘制精确的数字土壤水盐运移地图也成为常用的研究手段,其中中国(China)也是一个出现频次较高的关键词,说明中国是重要的研究对象或主力;③聚类簇主要是“盐渍化(salinization)”“土(soil)”“土壤盐渍化(soil salinization)”“水(water)”“地下水(groundwater)”“灌溉(irrigation)”“管理(management)”等关键词。表明盐渍化的形成原因是研究重点,且其相关过程蒸发、降水及区域、水系研究也很热门,由此,引申的各种灌溉和高效农业水利用方式也是热点研究之一;④聚类簇主要是“盐度(salinity)”“生长(growth)”“植物(plants)”“盐分胁迫(salt stress)”“盐分耐受性(salt tolerance)”等关键词。表明研究植物的耐受机制等对植物生长的影响是研究重点,且研究植物主要集中于大米、玉米和西红柿等,盐分胁迫条件下的应激反应如渗透调节、抗氧化、光合作用等过程也备受关注。标签视图(图1B)表明,现阶段盐渍土的主要研究方向是各元素、物质和生物的响应过程、水盐模拟等。
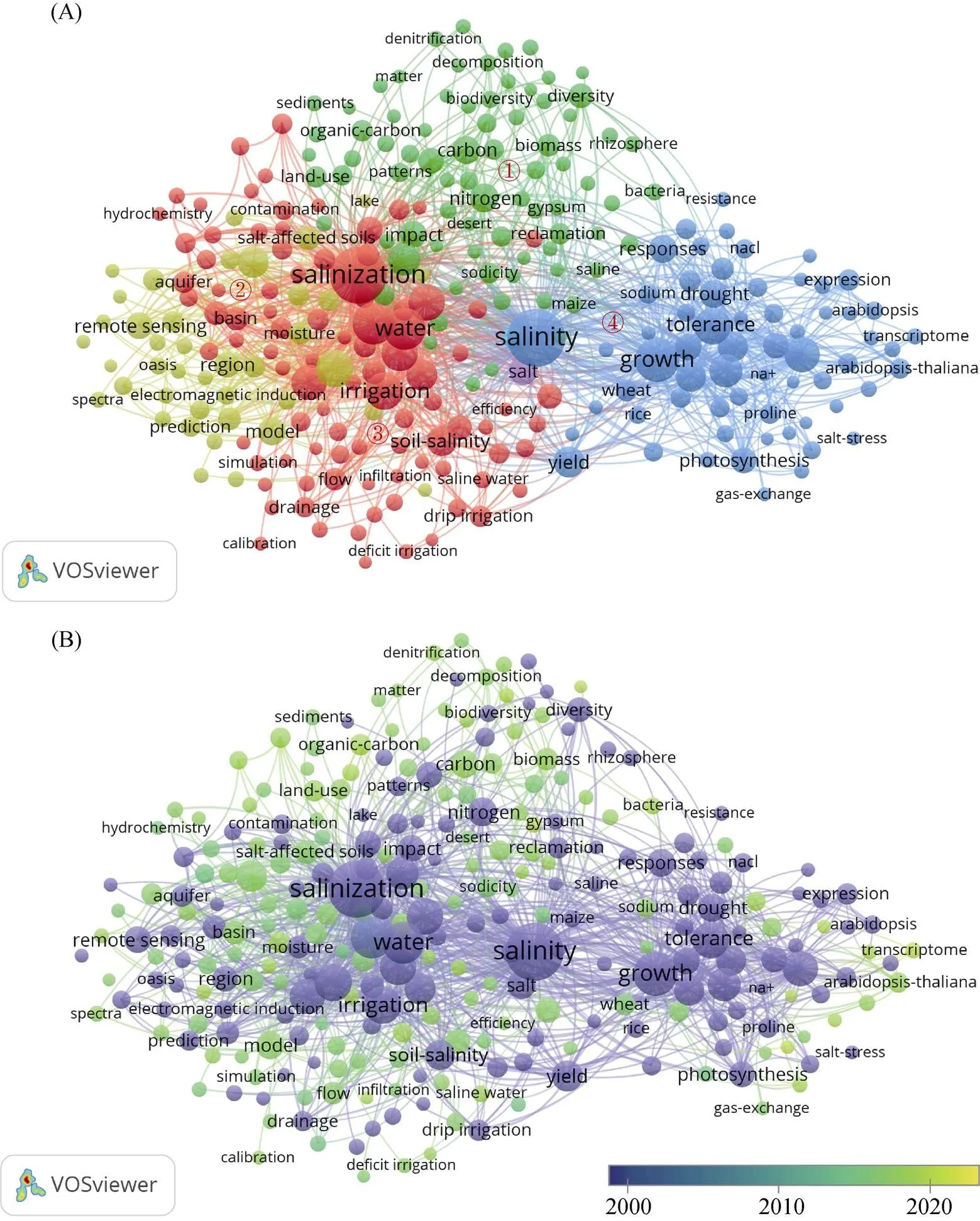
图1 2000—2022年Web of Science数据库以关键词“soil salinization”检索的聚类视图(A)及标签视图(B)
以Web of Science核心数据集同期文献为对象,以“(ALL=(soil salinization)) AND ALL=(phosphorus)”为检索式进行检索,得相关文献共计135篇,将其全部导入VOSviewer进行文献可视化分析(图2),由图2A和图2B的标签视图可以清楚地看出这一研究领域的时间转变:2010年之前主要的研究热点为盐渍土的灌溉、管理、质量、盐分等问题;2010—2020年的研究方向主要集中于盐分和磷的相互作用,两者对植物的影响(主要集中于玉米和小麦等粮食作物)、胁迫和耐受机制、土地利用方式等方面;2020年至今研究主要集中于磷的回收再利用、对作物的生长影响及产量效应、转化动力学过程、酶活性,重点是盐渍土中的磷过程和磷与碳、氮等其他大量营养元素的耦合作用及过程。通过图2C密度视图可以看出,2000年至今这一研究领域的研究关键词包括“磷(phosphorus)”“盐度(salinity)”“盐渍化(salinization)”“生长(growth)”“氮(nitrogen)”“碳(carbon)”,与图1B标签视图呈现的结果相一致,即主要的研究方向是盐渍土中各元素物质的循环转化过程。
2 盐分降低磷有效性的作用机制
盐分通过两方面直接影响磷的有效性:①阳离子作用。对于北方干旱、半干旱碱性土壤,通常是土壤中盐分离子Ca2+、Mg2+等与磷酸根离子结合使之迅速由可溶态转化成难溶态,并最终结合羟基、氧基等形成植物不可利用的羟基磷酸盐、氟磷酸盐;对于南方酸性土壤,磷酸根离子则通常与可溶的Al3+、Fe3+等形成磷酸盐沉淀,即转变成次生矿物,并最终被氧化铁铝胶膜包被覆盖成为“闭蓄态磷”,使磷酸盐丧失溶解释放的机会[29]。②盐分离子会与磷酸根竞争进入细胞膜的结合位点。据推测,膜水平上的物理和化学变化以及芽中磷吸收和积累的抑制是氯(Cl)和磷之间竞争以及竞争以外的相互作用的结果[30]。
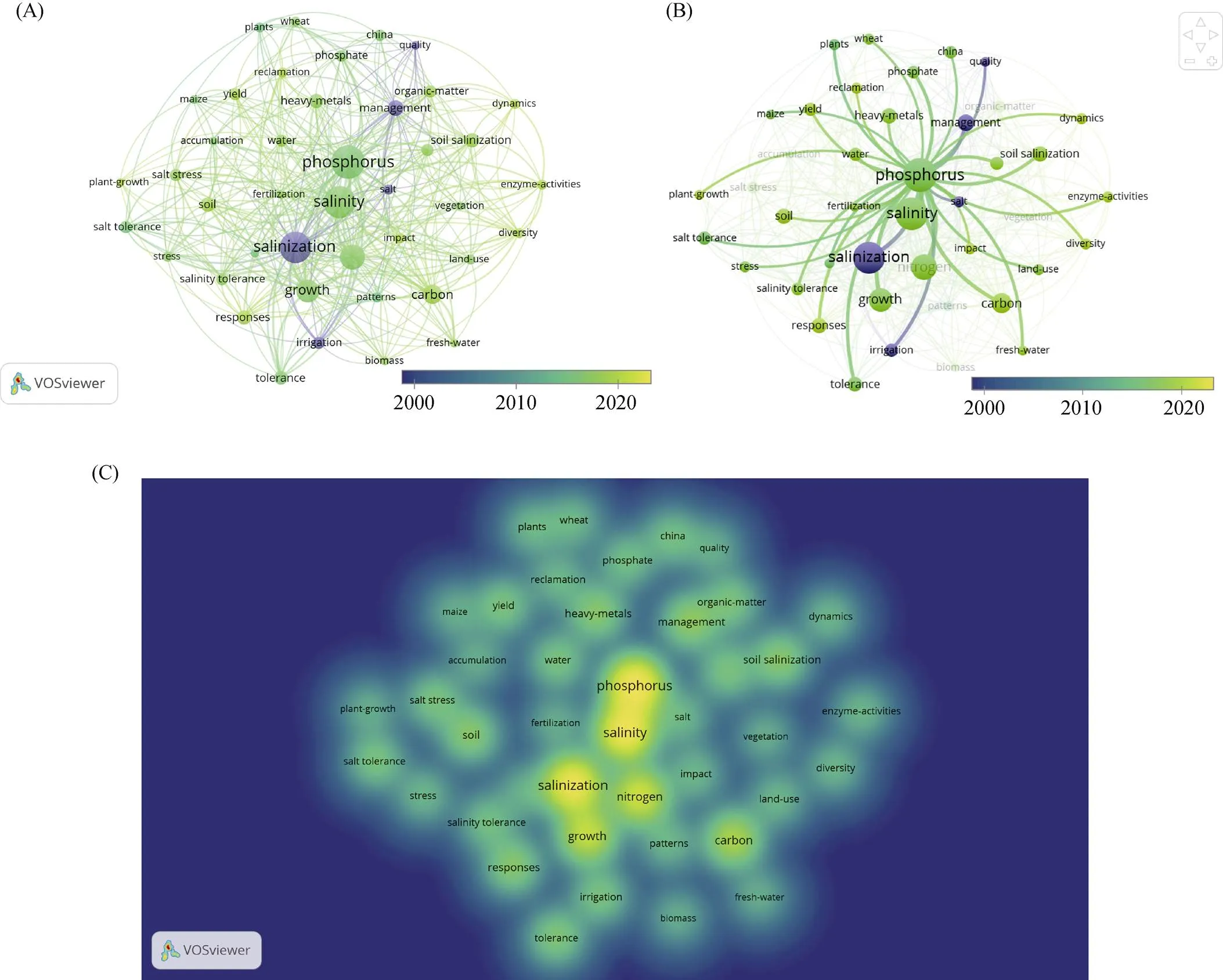
图2 2000—2022年Web of Science数据库以关键词“(ALL=(soil salinization)) AND ALL=(phosphorus)”检索生成的聚类视图(A)、标签视图磷部分(B)和密度视图(C)
而盐分主要通过影响植物和微生物的生命活动间接影响磷的生物有效性。①影响植物根系的渗透势及吸水性:盐分过量,植物不仅需要积累更多的有机和无机溶质来对抗外部环境中的低渗透势[31-32],且植物的吸水性会受到抑制。②盐分离子毒害作用和离子竞争:特定离子包括Na+、Cl–、H3BO4–、HCO3–等,这些离子对大多数植物都具有毒害作用[31-33],其次盐分胁迫会使植物体内产生大量的活性氧自由基,导致膜质系统被氧化破坏[5]。而盐分和养分离子穿过细胞膜被植物吸收时会产生离子竞争,例如,Na+会与K+竞争结合点位,Na+过量导致植物无法获得足够的K+而营养失衡[21,34-35]。③盐分离子的物理效应:尤其是大量Na+存在时,由于其单价离子较小的水合半径,其吸附的扩散层较宽和松散,阻碍了土壤胶体之间的吸附,使土壤结构变差,特别是湿润时还会促进土壤团聚体的崩解和分散,使土壤孔隙被阻塞,通气、透水性大大减弱,严重影响植物正常的生理活动[36]。④影响有机磷的矿化过程:盐渍土的高盐分和pH会抑制碱性磷酸酶的活性,制约有机磷的矿化分解过程,使有机磷的有效性降低[37]。⑤其他复杂作用:盐分还可能导致复杂的相互作用,影响植物代谢或内部营养需求[38]。
3 土壤盐分与磷素的相互、综合及交互作用效应
各文献中盐分和磷素的相互、综合及交互作用效应,尤其是交互作用,概念模糊,众说纷纭。在本研究中,盐分和磷素的相互、综合及交互作用效应均引用毛达如[39]的《植物营养研究方法》复因素试验方案设计中的基本概念定义,即相互作用是指两因素相互之间的影响;综合作用是指两因素的组合效应;交互作用是指两因素配合后相互促进而产生的效应,即两因素综合作用与各因素简单效应的差值,可正、可负、可以等于零。
3.1 土壤盐分对植物磷有效性的影响
养分吸收限制,尤其是磷缺乏,是降低作物生物量和粮食产量的最主要限制因素之一[40]。盐分含量过高引起的有效磷缺乏是盐渍农业生态系统中单独或同时存在的主要非生物胁迫,这对植物生长发育以及植物代谢均有负面影响[41-44]。尤其在干旱和半干旱地区,土壤有效磷缺乏和盐分胁迫是作物生产力的主要制约因素,盐分胁迫会降低磷肥的有效性[33]。无机磷分级结果显示,土壤含盐量的增加会使活性、中等活性无机磷含量降低,稳定性无机磷比例升高[37]。盐分胁迫不仅会降低鹰嘴豆等作物的磷吸收效率[45],且盐分胁迫会降低开心果()叶片、茎和根的磷含量[46]。盐渍土中,盐分胁迫及离子竞争使植物的磷素吸收进一步受到限制,磷固定作用的加强使有效磷含量进一步降低,最终皆导致磷生物有效性降低,因此生产实践中盐渍土中往往需要额外添加磷肥,然而磷肥的额外添加无法促进植物吸收相应的磷量,从而导致盐渍土中磷肥利用效率更低[16]。
但也有一些研究结果表明,盐度增加或对磷的吸收没有影响[28]。有文献报道,仅在80和100 mmol/L氯化钠(NaCl)条件下,西蓝花()叶片磷酸盐含量显著降低,而在根系中,磷酸盐含量随着盐浓度增加到40 mmol/L而增加,且随着盐浓度进一步增加到100 mmol/L而保持不变[47]。盐度对硬绳柄草()地上部磷浓度和磷吸收效率也没有实质性影响,与磷的有效性无关[48]。
因此,对于土壤溶液,盐分离子会促进磷酸盐离子的吸附和沉淀过程,使磷的有效性大大降低;但对于具体盐分环境的具体植物,磷有效性却又因盐分含量、植物种类、植物器官等不同而呈现不同的规律,这也为原位环境筛选适生抗性基因提供了理论支持。
3.2 外源磷添加对盐渍土中植物耐盐性的影响
通过施加适当的磷肥提高植物的耐盐性是一种很有潜力的策略,因为盐胁迫下,植物Na+可能会因施磷而减少,但这一反应似乎依赖于磷比率[49-50]。在大多数情况下,随着磷供应的增加,金皇后叶和茎K+/Na+比以及耐盐星茎和根K+/Na+比显著增加,表明磷肥提高了两个紫花苜蓿品种的耐盐性[33]。Talbi Zribi等[48]也得出,在盐渍土中添加磷可以缓解盐分的负面影响,并能提高耐盐性。且有报道称,在盐渍化土地的复垦方法中,如果氯化物为主要盐分物质,则可以通过施加磷肥提高作物产量,原因为施磷可以降低Cl–及Na+的积累,提高K+、Ca2+和P的吸收,一定程度上缓解盐分造成的离子竞争作用[51]。
然而,在两种紫花苜蓿的研究中只有部分结果支持以上结论:只有金皇后的叶Na+含量和耐盐星的茎Na+含量支持植物Na+含量随着磷添加量的增加而减少[33]。以开心果为试验对象时,施磷甚至增加了叶、茎Na+含量,而降低了根Na+含量和叶、茎、根K+含量[46],且研究显示提高开心果的耐盐性只需要适度的磷供应,而较高的磷供应会导致较高的磷积累,并在植物组织中产生毒性效应,导致生长和整体生产力下降[46,52]。还有研究显示,高或非限制性的磷水平不会影响苜蓿对氯化钠的反应[53],在高盐分条件下增加磷的有效性对风叶草的耐盐性也没有显著影响[54]。此外,与磷供应充足的植株相比,磷缺乏的盐处理植株却表现出更高的耐盐性[11]。综上,营养元素的添加可以增强或降低植物对干旱或盐分的抵抗力,或者根本没有效果,这取决于水分利用效率和盐胁迫程度[21]。
以上这些研究表明,各盐渍土对磷肥的响应各不相同,缘于各盐渍土土壤颗粒表面与磷酸盐离子反应的固定位点类型、位点总数等均不相同,继而导致不同的盐渍土对磷有不同的固定能力。对固持能力未达到饱和的盐渍土,施磷量要高于作物需磷量才有可能满足植物的正常养分需求或促进植物生产力和提高耐盐性;然而,当固持点位全部被磷酸盐离子占据时,过量的磷添加超过作物吸收量,就会产生磷的过度累积,不仅不会提高植物抵抗胁迫的能力,还可能产生离子毒害作用和环境问题。
3.3 盐分胁迫与磷缺乏对植物的综合及交互作用效应
磷缺乏和盐分胁迫的组合可以代表许多生长在钙质盐土中的植物的自然生长状况[26-27,55],但盐分胁迫和磷缺乏之间的综合作用尤其复杂。植物对其的反应在很大程度上取决于自身性状、生长阶段、土壤盐分水平和磷的形态以及试验的环境条件[53,56-57]。Talbi Zribi等[11]探讨了盐分胁迫和磷缺乏综合作用下大麦(L. cv. Manel)的生长差异和生理反应,结果显示,水培条件下磷缺乏对大麦生长的影响更为不利,盐分胁迫对磷缺乏大麦没有严重的不利影响,但在磷供应充足的情况下,盐分胁迫显著降低了植株的生物量,且这两种胁迫的影响不是简单的加和,因为植物对盐分胁迫和磷缺乏的反应与单独磷缺乏条件下相似。但Talbi Zribi等[48]在2018年利用相似的试验发现,盐分胁迫和磷缺乏处理分别显著降低了硬绳柄草生长速率和CO2同化率,其中,盐胁迫的影响更为显著;与对照组相比,磷缺乏和盐分胁迫显著降低了植株全株干重,分别降低65% 和76%,且两种胁迫呈相加效应,其同时存在时对植物生长影响最大,全株生物量下降幅度达91%。不同文献中关于磷缺乏和盐分胁迫对植物生长综合作用的结果也有所差异。在没有盐分或其他胁迫的情况下,影响营养元素可用性、吸收和分配的因素已是非常复杂[58],盐分胁迫的存在给作物的矿物质营养反应增加了新的复杂性[28]。因此,大多数自然生境中盐分胁迫和养分缺乏相互作用同时发生,但它们最终是否会影响作物产量或品质取决于盐分水平、盐分组成、作物种类,以及所涉及的养分和环境因素[38]。
此外,土壤磷缺乏和盐分胁迫可能以交互方式影响植物磷的获取和耐盐性。当植物受到多种胁迫时,可能会导致不同的胁迫应答,这些应答可能是多种胁迫效应的简单相加、协同或拮抗作用,这主要取决于植物的年龄、植物固有的抗逆性或敏感性以及胁迫的严重程度[59]。试验研究中,应将多种胁迫的组合视为非生物胁迫的一种新状态,不能直接从植物对单独施加的每种胁迫的相应反应中推断出来[60]。
综上,当土壤出现高盐分和磷缺乏等多种非生物胁迫时,其综合效应往往不是单因素简单效应的加和,且因盐分含量、盐分组成、植物种类而呈现不同的规律。在盐渍土农业生产中,应当将两种非生物胁迫的组合看作一种新问题,进而积极探索两种非生物胁迫的交互作用效应,为提高盐渍土有效磷含量和磷养分利用效率提供新思路、新方法。
4 盐渍土磷有效性提升策略
相较于根系磷吸收动力学特征,盐渍土的特性对磷有效性影响更为显著。因此,几十年来人们积极探索改善盐渍土理化性质的新方法、新思路,以促进磷营养的均衡及充分吸收。
4.1 物理提升策略
盐渍土磷有效性物理提升策略主要包括实行合理的耕作措施、种植制度和有效的施肥方式。研究显示,深松破板、粉垄深旋、平整土地等耕作措施可以阻断东北苏打盐渍土中向上运送水盐的毛细管,提高散墒作用,减轻盐分对植物根的毒害,从而促进植物营养吸收和正常的生理活动[61]。轮、间、套作等多样化种植模式也可以促进不同作物间的养分协作,加快营养物质的转化和循环,增加作物利用养分的能力[61],且轮作方式的不同主要影响无机磷及不稳定有机磷含量[62]。有研究指出,长期种植粮食作物土壤总磷消耗量远大于草粮混播(草和粮食同时存在的状态)[63];免耕土壤中有机碳、有机磷等含量高,会刺激微生物产生高活性磷酸酶,促进有机磷矿化[64]。通过生物固氮和富氮绿肥种植等多种农业生态功能也可以提高营养元素利用效率,据研究,豆类种植对非豆类作物的生长发育有益[40,65-66]。有效的施肥方式如磷肥集中施用(条施、点施),可以有效减少磷肥与其他物质的接触,进而提高固磷能力较强土壤的磷素有效性[53]。水分条件也会影响磷的有效性,淹水会使土壤的氧化还原电位降低,酸性土壤pH升高,碱性土壤pH降低,进而使有效磷含量升高[67],此时磷的吸附解吸行为主要受铁氧化物形态转化的控制[68]。
总之,物理提升策略主要通过改善盐渍土土壤结构、通气透水性和提高植物物种多样性等途径加强农业生态系统的磷循环。其优点是绿色、经济,但效果相对较小且缓慢,且通过提高植物物种丰富度来促进对潜在磷库磷素的吸收存在众多技术挑战,包括各物种的除草剂和杀虫剂存在相互毒害作用、施肥种类和用量不同、机械化管理受限等。
4.2 化学提升策略
盐渍土磷有效性化学提升策略主要包括合理施用无机和有机改良剂。无机改良剂主要集中于磷石膏的广泛研究。近年来,氮磷肥等的适量施用,尤其是氮磷减施成为研究热点。有机改良剂主要包括黄腐酸、腐殖酸等有机酸,生物质炭,秸秆,生活污水污泥,动物粪便等的应用。
磷石膏是被广泛应用于盐渍土改良的化学改良剂,它不仅可以代换Na+,促进盐分离子淋溶,改善土壤物理结构,而且在磷石膏颗粒周围土壤pH可达3 ~ 4,形成局部的酸性环境,继而促进无机磷的溶解和释放[69]。目前研究多集中于无机和有机改良剂的结合使用。有研究显示,矿质磷和有机肥的联合施用不仅显著增加了土壤磷组分,而且提高了小麦的产量和品质[70]。但在化肥(过磷酸钙)结合有机肥和改良剂(CaSO4)改良黄淮海平原轻、中度盐渍土的试验中发现,所有施肥处理的有效磷含量均降低,可能是由于改良剂促进了植物的生长,加强了磷素的吸收,同时CaSO4的施用引入大量Ca2+,其与PO43–结合生成难溶的沉淀,从而减少了有效磷的含量[71]。可见,无机改良剂搭配有机改良剂在促进植物生长方面效果显著,在有效磷含量提升方面存在差异。
近期有研究表明,在盐渍土中,可通过减少氮肥施用量来提高磷的有效性[72]。氮肥减量不仅可以维持作物产量,且在降低土壤中的Na+和Cl–的同时,增加了0 ~ 10 cm土层有效磷含量和碱性磷酸酶活性[73],甚至在提高有效磷含量方面,30% 氮肥减施处理优于15% 氮肥减施处理和常规化肥施用量处理[74]。这说明该盐渍土不仅有30% 的减氮潜力,而且化肥减施能更好地促进磷素的活化和循环。随着氮肥施用量的增加,不稳定的有效态磷Ca2-P和迟效态的Ca8-P含量均相应减少[72]。可见,氮肥施用过量,不仅造成资源浪费,而且对其他养分(磷)的转化也会产生影响,使其有效性降低。在盐渍土中,适量施用磷肥也可以提高紫花苜蓿的耐盐性,增加其产量[33]。在大麦–玉米双季轮作试验中,磷肥减施能显著提高轻、中度盐渍土的累积磷肥利用率[37]。
氮肥搭配黄腐酸不但可以降低Na+和Cl–含量,而且可以提高冬小麦的产量和碱性磷酸酶的活性,且当氮肥减量配施黄腐酸时,促进了冬小麦的氮、磷吸收效率[73-74]。这充分说明化肥减施不仅不会对产量造成负面影响,且搭配有机酸等有机改良剂可以促进氮、磷等营养物质更好地吸收利用。即使在过量施用氮肥的情况下,氮肥配施黄腐酸也使盐渍土的磷有效性提高,在培养后期,有效态的Ca2-P含量提高,无效态的Ca10-P含量降低,促进了磷的释放[72]。轻、中度盐渍土中施入腐殖酸也均可以减少盐渍土中HCl-Pi含量,促进磷的有效化,但对活性有机磷(NaHCO3-Po)和中等活性有机磷(NaOH-Po)的活化作用有限[37]。
生物质炭也可以显著提高盐渍土磷的有效性。生物质炭施入轻、中度盐渍土,可以促进Na+、Cl–的淋洗,提高Ca2+、Mg2+含量,显著降低钠吸附比(SAR),改善盐分离子组成和土壤物理结构,提高活性无机磷含量[37]。高珊等[75]指出,在轻、中度盐渍土中添加生物质炭,与常规施磷处理相比,有效磷含量分别可以提高40.72%、84.8%,且能显著提高土壤活性无机磷比例。且适宜用量的生物质炭结合黄腐酸,对江苏滨海围垦滩涂区重度盐渍土的系统培肥和肥料利用率提升均有促进作用[76]。在黄河三角洲地区盐渍土中,施用MgO改性的生物质炭增加了土壤有效磷含量,并导致水稻产量提高[77]。在田间试验中,两种铁改良生物质炭也表现出显著的有效磷提高效果,且氧化亚铁改良生物质炭和三氯化铁改良生物质炭分别使土壤有效磷含量增加了78.6% 和90.3%[78]。生物质炭含有大量难降解碳[79],这些碳可在土壤中保留100 ~ 1 000年,加入生物质炭可能同时是增加土壤碳固存的有效途径,从而减缓全球温室气体排放[79-80]。因此,生物炭对于盐渍土的磷有效性提升和碳固存都有巨大的研究价值。
为了实现资源的充分利用,生物质材料及动物粪便等有机物料广泛用于提升盐渍土磷素有效性。生物质材料的应用以秸秆还田为典型策略。研究表明,与秸秆移除相比,秸秆还田可以提高滨海盐渍土磷的活化率和有效磷含量,磷的表观回收率也提高了14% ~ 27%,磷肥的施用量降低了27% ~ 38%,且对籽棉的产量不会造成影响[81]。说明对盐渍土进行秸秆还田不仅能促进磷素的活化,而且能减少矿物质磷的施用,同时又能维持当地的作物生产。在苏北新围垦滩涂农田进行有机肥与秸秆覆盖综合试验发现,两者共同作用对轻度偏中度盐碱障碍消减、pH调控、作物产量都有显著的促进作用[82]。张秀敏等[83]汇总了我国典型区域盐渍土的秸秆还田和配套技术,其中,内蒙古引黄灌区、河套灌区采用秸秆翻耕或覆盖还田,辅以灌排结合、耕作松土和种植耐盐碱植物;东北苏打盐渍土区采用秸秆深翻还田,配套化学改良剂、灌溉排水和生物改良;滨海盐渍土区采用秸秆覆盖还田,配套起高垄、亚沙、节水灌溉和耐盐植物选育与栽培技术。秸秆粉碎深翻,实现了有机质与土壤充分混合,其分解产物可以改善土壤团聚体的结构和功能,其中有机酸可以降低土壤pH,与磷酸根竞争结合点位,进而提高磷的有效性;秸秆覆盖的作用类似地膜覆盖,起到减少水分蒸散、阻止盐分上行、加速盐分淋溶、减轻盐分对根系毒害的作用[83]。总之,秸秆还田是一项提高盐渍土磷素有效性的有效方法。但由于各区域盐渍土的土壤类型等因素不同,各地的秸秆覆盖方式、时间、粉碎长度、深度、种类及其配套进行的措施也不同。且由于秸秆的碳氮比高,微生物分解过程可能出现净固定,与植物竞争养分,进而很多情况下需要额外地补充氮肥。但对秸秆进行预处理,例如堆肥等,显著降低了碳氮含量而使磷富集,能使之成为更高效的磷源。生活污水污泥等生物固体对盐渍土磷素有效性提升也有重要应用。以3种改良物质棉秆粉、生活污水污泥和海滩砂进行的完全随机区组试验发现,除海滩砂外,其他处理都可以提高滨海盐渍土磷的有效性,且生活污水污泥可以显著降低盐渍土电导率,然而相比于氮素有效性,复合处理对磷素有效性却无显著影响[84]。也有研究显示,通过在滨海苏打盐渍土中加入猪粪作为改良剂而实现的磷输入,也可以显著提高土壤生物有效磷含量和土壤磷稳定性[85]。而且,猪粪配合滴灌工程措施,比自然休耕和短期猪粪滴灌能更有效提高滨海盐渍土磷的有效性[86]。总之,许多类型的生物质材料、生物固体和动物粪便均可以作为盐渍土中磷素有效性提升的有效改良剂,且对其处理加工(例如堆肥)可能会使其成为更有效的磷素来源。
现阶段,盐渍土磷素有效性化学提升策略主要以有机改良剂为基础措施,且有机改良剂与无机改良剂或改良方式的复合应用在绝大多数情况下显示出更为良好的提升效果,因此,探索有机改良剂与其他各种有效改良方式结合促进盐渍土磷素有效性提升的方法有巨大潜力。施用有机改良剂虽然能改善各种土壤性质,但其矿化过程缓慢,养分含量有限,且很可能增加土壤的潜在风险,如增加土壤的重金属含量(镉、镍、铅)[70,87],因此,有机改良剂的应用效应仍需进一步探究和试验验证。
4.3 生物提升策略
利用微生物改良盐渍土可能是一种环境可持续、更安全、更有效的方法,其中嗜盐微生物有可能去除盐渍土中的盐分,为研究微生物的抗逆性、适应和反应过程提供了独特的原型,这些过程可能因此被整合到农作物中,以应对气候变化造成的胁迫[88]。近年来,在菌根真菌、解磷细菌提高养分利用效率及机理研究方面也取得了一系列新突破[89]。此外,通过将磷高效利用基因转移到耐盐植物中实现的遗传物质整合也逐渐得到广泛关注。
植物促生菌(PGPB:plant growth promoting bacteria)可以在农业耕作系统中成功地作为替代策略以抵抗作物面临的大多数非生物胁迫,提高作物产量,并将合成肥料的使用量降至最低[90]。研究显示,节肢杆菌()和芽孢杆菌()有助于番茄植株在受控温室条件下更好地应对高盐分和有效磷缺乏的双重胁迫[22]。当NaCl以0.2% 浓度存在时,许多微生物提高了无机磷增溶率[91]。然而,盐分胁迫减少了土壤颗粒吸附的磷,并抑制了植物根系对磷的吸收,盐分胁迫的上述有益影响(提高无机磷增溶率)可能因此而被抵消[92]。而且,在盐渍土中,植物促生根际菌(PGPR:plant growth promoting rhizobacteria)的溶磷活性通常会降低[93]。因此,细菌适应盐渍土有效磷缺乏的能力对其生长、存活和繁殖至关重要。细菌的存活还取决于其运动能力,使其在胁迫环境中具有持久性优势,这是筛选细菌的一个共同特征[22]。
寻找具有耐盐性和高活力等特性的溶磷细菌(PSB),可为改善有效磷缺乏和盐渍土的作物生产力开辟新的方向[22]。解磷微生物,尤其是耐盐解磷微生物,不仅可以加速磷的分解过程,提高磷的有效性,而且可以改善土壤的物理结构,有助于盐渍土的复垦和改良,被认为是高效、经济的磷活化剂[16]。解磷微生物能分泌有机酸,酸化根际土壤;能螯合金属离子,形成铁载体,而释放磷酸盐;能分泌胞外多糖,溶解磷酸盐沉淀;能分泌生长激素和抗氧化物质,促进盐胁迫生境植物的生长[94-96],且这些代谢物有抑制病原体的作用。微生物菌剂尽管具有溶磷特征,但主要还是通过刺激根系生长间接提高磷的有效性[97],需要进一步的研究来揭示耐性应激微生物的多种隐藏机制。针对不同的胁迫条件,解磷微生物会表现出不同的生理活性。因此,针对不同盐渍生境,应分离出适应具体环境的高效菌株,来促进低磷环境植物的营养吸收和正常生长。此外,菌种的繁殖和菌剂的生产也是需要攻克的难题。
丛枝菌根真菌(AMF)是陆地生态系统中最普遍的内共生体,能与约74% 的被子植物根形成共生关系[98]。在盐生环境中通常可观察到,AMF与植物联合作用产生共生菌根,是缓解盐胁迫、磷短缺的重要植物策略[99-102]。石榴植株生长对盐分敏感,但接种AMF后缓解了盐分的负面影响[50]。AMF的菌丝体也能增加土壤的抗水性能,使土壤团聚体的水稳定性得以维持[102]。基因研究表明,植物诱导了一组常见的菌根诱导基因,但也存在变异,表明丛枝菌根(AM)共生体中存在功能多样性,共生相关基因在不同AM组合中的差异表达与真菌种类、植物基因型和环境因素有关[103],且AM必须与根系形成共生体才能完成生活史,尚不能纯培养。因此,通过AM直接摄取途径调节磷素有效性仍需要进一步研究[99,104]。生长在天然盐渍栖息地的物种具有遗传变异性,这是耐盐种群进化的基础[105-107]。通过磷高效基因型物种加强磷循环是保护不可再生磷资源、提高作物磷营养水平和减轻负面环境影响的一种颇具前景的方法[38]。据研究,磷高效基因型菜豆在表层土壤中浅根丰富,该土层磷素含量也相对较高[108]。可见,探索和培育耐性物种为有效利用盐渍土提供了一个更为实际的解决方案。但是,植物磷吸收和利用的主要生理和遗传因素已研究证实,因而对现代作物品种的磷利用效率进行遗传改良的成功率可能会很有限。
上述生物资源因其符合绿色环保、可持续等特征而被应用到盐渍土磷素有效性提升的各种农业策略中,尤其是各种微生物肥料的广泛研究和推广中。但微生物肥料的效果不仅取决于自身的性质,而且与施用的具体环境密切相关,这造成很多微生物存在生防活性不稳定、定殖差异大等问题,因此今后的研究应更加注重微生物、植物、环境等的综合考虑,以建立新环境下的生态平衡。
5 小结与展望
5.1 小结
盐渍土磷有效性提升措施紧紧围绕磷固定的因素开展,根据其可行性包括以下几个方面:①调整pH。磷有效性最高的pH范围为6 ~ 7,大部分基于此原理添加的改良材料可以形成局部酸性环境,促进磷酸根离子的释放,但往往由于土壤的缓冲能力强,改良材料影响的土壤范围较小,H+被快速中和,酸性环境持续时间较短,影响不大。②利用阳离子和阴离子效应。改良剂中含有的高价阳离子与Na+发生阳离子交换反应,配合淋溶可达到改善土壤通气透水性能、提高土壤团聚体功能的作用;阴离子效应是利用阴离子与磷酸根离子竞争结合位点,来实现提高磷酸根离子有效性的作用。③提高有机质含量。有机质可以掩蔽磷酸根结合位点,可以螯合金属离子,可以通过矿化作用,促进磷酸根离子释放;而有机质分解产生的有机酸不仅可以降低土壤pH,也可以与磷酸根离子竞争结合点位,从而提高磷的有效性。④引进微生物。微生物与植物形成共生关系或通过其新陈代谢过程产生相关酶组分,促进植物有效磷水平提高。但盐渍土普遍存在有机质含量低的现状,导致整体环境无法供微生物正常生存,而使其在现实应用中效果较差。⑤改善水分环境。水分是一切反应的溶剂。目前较多研究聚焦盐渍土水稻种植,即是利用淹水条件造就还原环境,促使难溶的磷酸盐矿物转变成相对可溶的状态。其中,提高有机质含量是提高盐渍土磷有效性可行性较高的策略之一。盐渍化农田有机质含量低,需要加强秸秆还田、绿肥作物种植等有机物料引入措施,重视基础培肥。
5.2 展望
多种改良方法的结合是实现盐渍土磷素有效性提升的新方向。但应定期对其他物质或元素进行监测,防止改良剂的添加引入新的污染物。盐渍土改良和地力提升是一个长期的过程,需要长期的地力监测管理策略,以维持有效磷水平的相对稳定。
盐渍土磷有效性提升不仅要促进根系从盐渍土中更充分地吸收有效态磷,而且需要提高植物内部磷的运输、转化效率。这些动态过程的精准监测需要放射性同位素的标记和示踪,现阶段研究常用的为32P、33P以及R值(18O/16O),但迫于需要大规模辐射防护、安全问题以及田间农业管理实践的压力,目前只局限于实验室条件,要建立与现实环境条件中具体转化过程的关系仍然很艰难。
近年来,根际的营养吸收及活化作用被广泛认知,根际过程不仅有助于磷矿溶解和捕获少量可溶解磷化合物,而且解磷微生物、植物促生菌等主要分布于根际。因此,不同类型的根际微区研究十分重要。然而,许多研究也显示,单一的根际研究不能反映植物与土壤及土壤微生物等多介质、多界面的相互作用和响应,这就要求根际的研究应该追根溯源,系统地建立一个物质、能量及信息传输转化的完整运作体系。
现阶段施用磷肥仍然是提高土壤磷有效性和作物产量的有效手段,但不同土壤的施用量应根据具体状况进行相应调整。尤其在盐渍条件下,由于土壤吸附和矿物沉淀降低了磷的有效性,实践中需要在盐渍土中重复施用磷,磷流失的风险大大增加。因此,需要加强对土壤渗滤液中磷浓度的监测,以降低水环境污染风险。同时,要基于不同盐度、基础营养状况、植物种类、土壤类型等因素,确定合理的施肥措施。另外,肥料配方的创新需要加强,应充分发挥营养元素之间的协同效应,积极探索其他营养元素与磷的非特异性相互作用和次生养分相互作用。总之,化肥的适当施用、土壤的实地特性和作物的实际需求是盐渍土磷肥高效利用的关键。
[1] Wicke B, Smeets E, Dornburg V, et al. The global technical and economic potential of bioenergy from salt-affected soils[J]. Energy & Environmental Science, 2011, 4(8): 2669–2681.
[2] 杨劲松, 姚荣江, 王相平, 等. 中国盐渍土研究: 历程、现状与展望[J]. 土壤学报, 2022, 59(1): 10–27
[3] Yadav A N, Saxena A K. Biodiversity and biotechnological applications of halophilic microbes for sustainable agriculture[J]. Journal of Applied Biology & Biotechnolog, 2018: 48–55.
[4] Bertrand A, Dhont C, Bipfubusa M, et al. Improving salt stress responses of the symbiosis in alfalfa using salt-tolerant cultivar and rhizobial strain[J]. Applied Soil Ecology, 2015, 87: 108–117.
[5] Munns R, Tester M. Mechanisms of salinity tolerance[J]. Annual Review of Plant Biology, 2008, 59: 651–681.
[6] Thurston A. The disappearing[J]. Annals of Internal Medicine, 2015, 163(4): 310.
[7] Ma J C, He P, Xu X P, et al. Temporal and spatial changes in soil available phosphorus in China (1990–2012)[J]. Field Crops Research, 2016, 192: 13–20.
[8] Huang C Y, Roessner U, Eickmeier I, et al. Metabolite profiling reveals distinct changes in carbon and nitrogen metabolism in phosphate-deficient barley plants (L.)[J]. Plant and Cell Physiology, 2008, 49(5): 691–703.
[9] Zhu F L, Qu L Y, Hong X G, et al. Isolation and characterization of a phosphate-solubilizing halophilic bacteriumsp. YCWA18 from daqiao saltern on the coast of Yellow Sea of China[J]. Evidence-Based Complementary and Alternative Medicine: ECAM, 2011, 2011: 615032.
[10] Ros M B H, Koopmans G F, van Groenigen K J, et al. Towards optimal use of phosphorus fertiliser[J]. Scientific Reports, 2020, 10(1): 1–8.
[11] Talbi Zribi O, Abdelly C, Debez A. Interactive effects of salinity and phosphorus availability on growth, water relations, nutritional status and photosynthetic activity of barley (L.)[J]. Plant Biology, 2011, 13(6): 872–880.
[12] Vance C P, Uhde-Stone C, Allan D L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource[J]. New Phytologist, 2003, 157(3): 423–447.
[13] Veneklaas E J, Lambers H, Bragg J, et al. Opportunities for improving phosphorus-use efficiency in crop plants[J]. New Phytologist, 2012, 195(2): 306–320.
[14] Feng G, Gai J P, Feng X H, et al. Strategies for improving fertilizer phosphorus use efficiency in Chinese cropping systems[J]. Frontiers of Agricultural Science and Engineering, 2019, 6(4): 341.
[15] Cao N, Chen X P, Cui Z L, et al. Change in soil available phosphorus in relation to the phosphorus budget in China[J]. Nutrient Cycling in Agroecosystems, 2012, 94(2): 161–170.
[16] Cong W F, Suriyagoda L D B, Lambers H. Tightening the phosphorus cycle through phosphorus-efficient crop genotypes[J]. Trends in Plant Science, 2020, 25(10): 967–975.
[17] Penn C, Camberato J. A critical review on soil chemical processes that control how soil pH affects phosphorus availability to plants[J]. Agriculture, 2019, 9(6): 120.
[18] Power J F, Grunes D L, Reichman G A, et al. Soil temperature effects on phosphorus availability1[J]. Agronomy Journal, 1964, 56(6): 545–548.
[19] Grandgirard J, Poinsot D, Krespi L, et al. Costs of secondary parasitism in the facultative hyperparasitoiddubius: Does host size matter?[J]. Entomologia Experimentalis et Applicata, 2002, 103(3): 239–248.
[20] Fink J R, Inda A V, Tiecher T, et al. Iron oxides and organic matter on soil phosphorus availability[J]. Ciência e Agrotecnologia, 2016, 40(4): 369–379.
[21] Hu Y C, Schmidhalter U. Drought and salinity: A comparison of their effects on mineral nutrition of plants[J]. Journal of Plant Nutrition and Soil Science, 2005, 168(4): 541–549.
[22] Tchakounté G V T, Berger B, Patz S, et al. Selected rhizosphere bacteria help tomato plants cope with combined phosphorus and salt stresses[J]. Microorganisms, 2020, 8(11): 1844.
[23] Kumar A, Verma J P. Does plant—Microbe interaction confer stress tolerance in plants: A review?[J]. Microbiological Research, 2018, 207: 41–52.
[24] Shen J B, Yuan L X, Zhang J L, et al. Phosphorus dynamics: From soil to plant[J]. Plant Physiology, 2011, 156(3): 997–1005.
[25] Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell[J]. Annals of Botany, 2009, 103(4): 551–560.
[26] Huang B R. Role of root morphological and physiological characteristics in drought resistance of plants[M]//Plant- Environment Interactions. Beijing: CRC Press, 2000.
[27] Naidu R, Rengasamy P. Ion interactions and constraints to plant nutrition in Australian sodic soils[J]. Soil Research, 1993, 31(6): 801.
[28] Girsova M A, Golovina G F, Drozdova I A, et al. Infrared studies and spectral properties of photochromic high silica glasses[J]. Optica Applicata, 2014, 44(2): 337–344.
[29] Jiang H H, Qi P S, Wang T, et al. Isolation and characterization of halotolerant phosphate-solubilizing microorganisms from saline soils[J]. 3 Biotech, 2018, 8(11): 461.
[30] Papadopoulos I, Rendig V V. Interactive effects of salinity and nitrogen on growth and yield of tomato plants[J]. Plant and Soil, 1983, 73(1): 47–57.
[31] Short D C, Colmer T D. Salt tolerance in the Halophytesubsp.[J]. Annals of Botany, 1999, 83(3): 207–213.
[32] Nublat A, Desplans J, Casse F, et al. sas1, anmutant overaccumulating sodium in the shoot, shows deficiency in the control of the root radial transport of sodium[J]. The Plant Cell, 2001, 13(1): 125–137.
[33] Su R, Zhang Z K, Chang C, et al. Interactive effects of phosphorus fertilization and salinity on plant growth, phosphorus and sodium status, and tartrate exudation by roots of two alfalfa cultivars[J]. Annals of Botany, 2022, 129(1): 53–64.
[34] Hu Y C, Fricke W, Schmidhalter U. Salinity and the growth of non-halophytic grass leaves: The role of mineral nutrient distribution[J]. Functional Plant Biology, 2005, 32(11): 973.
[35] Long R C, Sun H, Cao C Y, et al. Identification of alkali-responsive proteins from early seedling stage of two contrastingspecies by iTRAQ-based quantitative proteomic analysis[J]. Environmental and Experimental Botany, 2019, 157: 26–34.
[36] Zahedi S M, Hosseini M S, Abadía J, et al. Melatonin foliar sprays elicit salinity stress tolerance and enhance fruit yield and quality in strawberry (Duch.)[J]. Plant Physiology and Biochemistry, 2020, 149: 313–323.
[37] 高珊. 苏北滨海盐渍农田土壤磷素有效性及其增效调控研究[D]. 北京: 中国科学院大学, 2019.
[38] Grattan S R, Grieve C M. Salinity–mineral nutrient relations in horticultural crops[J]. Scientia Horticulturae, 1998, 78(1/2/3/4): 127–157.
[39] 毛达如. 植物营养研究方法[M]. 2版. 北京: 中国农业大学出版社, 2005.
[40] Vance C P. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources[J]. Plant Physiology, 2001, 127(2): 390–397.
[41] Yoneyama K, Xie X N, Kim H I, et al. How do nitrogen and phosphorus deficiencies affect strigolactone production and exudation?[J]. Planta, 2012, 235(6): 1197–1207.
[42] Kosová K, Práil I T, Vítámvás P. Protein contribution to plant salinity response and tolerance acquisition[J]. International Journal of Molecular Sciences, 2013, 14(4): 6757–6789.
[43] Carstensen A, Herdean A, Schmidt S B, et al. The impacts of phosphorus deficiency on the photosynthetic electron transport chain[J]. Plant Physiology, 2018, 177(1): 271–284.
[44] Mokrani S, Nabti E H, Cruz C. Current advances in plant growth promoting bacteria alleviating salt stress for sustainable agriculture[J]. Applied Sciences, 2020, 10(20): 7025.
[45] Sadji-Ait Kaci H, Chaker-Haddadj A, Aid F. Enhancing of symbiotic efficiency and salinity tolerance of chickpea by phosphorus supply[J]. Acta Agriculturae Scandinavica, Section B — Soil & Plant Science, 2018, 68(6): 534–540.
[46] Shahriaripour R, Tajabadi Pour A, Mozaffari V. Effects of salinity and soil phosphorus application on growth and chemical composition of pistachio seedlings[J]. Communications in Soil Science and Plant Analysis, 2011, 42(2): 144–158.
[47] López-Berenguer C, Carvajal M, Garcéa-Viguera C, et al. Nitrogen, phosphorus, and sulfur nutrition in broccoli plants grown under salinity[J]. Journal of Plant Nutrition, 2007, 30(11): 1855–1870.
[48] Talbi Zribi O, Slama I, Trabelsi N, et al. Combined effects of salinity and phosphorus availability on growth, gas exchange, and nutrient status of[J]. Arid Land Research and Management, 2018, 32(3): 277–290.
[49] Bargaz A, Nassar R M A, Rady M M, et al. Improved salinity tolerance by phosphorus fertilizer in tworecombinant inbred lines contrasting in their P-efficiency[J]. Journal of Agronomy and Crop Science, 2016, 202(6): 497–507.
[50] Arab Yarahmadi M, Shahsavani S, Akhyani A, et al. Pomegranate growth affected by arbuscular mycorrhizae, phosphorus fertilizer, and irrigation water salinity[J]. Communications in Soil Science and Plant Analysis, 2018, 49(4): 478–488.
[51] Gulshan N, Shahbaz M, Aisha N. Interactive effect of rooting medium application of phosphorus and NaCl on plant biomass and mineral nutrients of rice (L.)[J]. Pakistan Journal of Botany, 2008, 40(4): 1601–1608.
[52] Aslam M, Flowers T J, Qureshi R H, et al. Interaction of phosphate and salinity on the growth and yield of rice (L.)[J]. Journal of Agronomy and Crop Science, 1996, 176(4): 249–258.
[53] Rogers M E, Grieve C M, Shannon M C. Plant growth and ion relations in lucerne (L.) in response to the combined effects of NaCl and P[J]. Plant and Soil, 2003, 253(1): 187–194.
[54] Talbi Zribi O, Barhoumi Z, Kouas S, et al. Insights into the physiological responses of the facultative halophyteto the combined effects of salinity and phosphorus availability[J]. Journal of Plant Physiology, 2015, 189: 1–10.
[55] Zribi O T, Hessini K, Trabelsi N, et al.maintains adequate gas exchange, pigment composition and phenolic contents under combined effects of salinity and phosphorus deficiency[J]. Australian Journal of Botany, 2017, 65(5): 453.
[56] Loupassaki M H, Chartzoulakis K S, Digalaki N B, et al. Effects of salt stress on concentration of nitrogen, phosphorus, potassium, calcium, magnesium, and sodium in leaves, shoots, and roots of six olive cultivars[J]. Journal of Plant Nutrition, 2002, 25(11): 2457–2482.
[57] Villora G, Moreno D A, Pulgar G, et al. Salinity affects phosphorus uptake and partitioning in zucchini[J]. Communications in Soil Science and Plant Analysis, 2000, 31(3/4): 501–507.
[58] Abbas G, Chen Y L, Khan F, et al. Salinity and low phosphorus differentially affect shoot and root traits in two wheat cultivars with contrasting tolerance to salt[J]. Agronomy, 2018, 8(8): 155.
[59] Pandey P, Ramegowda V, Senthil-Kumar M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms[J]. Frontiers in Plant Science, 2015, 6: 723.
[60] Mittler R. Abiotic stress, the field environment and stress combination[J]. Trends in Plant Science, 2006, 11(1): 15–19.
[61] 黄晶, 孔亚丽, 徐青山, 等. 盐渍土壤特征及改良措施研究进展[J]. 土壤, 2022, 54(1): 18–23.
[62] 宇万太, 陈欣, 张璐, 等. 磷肥低量施用制度下土壤磷库的发展变化——Ⅰ.土壤总磷库和有机、无机磷库[J]. 土壤学报, 1996, 33(4): 373–379.
[63] Vázquez M E, Noellemeyer E, Coremberg P. The dynamics of different organic and inorganic phosphorus fractions in soils from the south of Santa Fe Province, Argentina[J]. Communications in Soil Science and Plant Analysis, 1991, 22(11/12): 1151–1163.
[64] McKenzie R H, Stewart J W B, Dormaar J F, et al. Long-term crop rotation and fertilizer effects on phosphorus transformations: II. In a Luvisolic soil[J]. Canadian Journal of Soil Science, 1992, 72(4): 581–589.
[65] Isaac M E, Harmand J M, Drevon J J. Growth and nitrogen acquisition strategies ofseedlings under exponential phosphorus additions[J]. Journal of Plant Physiology, 2011, 168(8): 776–781.
[66] 刘建玲, 张凤华. 土壤磷素化学行为及影响因素研究进展[J]. 河北农业大学学报, 2000, 23(3): 36–45.
[67] 朱昌锋. 淹水对土壤磷有效性影响的研究[D]. 重庆: 西南大学, 2008.
[68] 邵兴华. 水稻土淹水过程铁氧化物转化对磷饱和度和磷、氮释放的影响[D]. 杭州: 浙江大学, 2005.
[69] 杨莉琳, 李金海. 我国盐渍化土壤的营养与施肥效应研究进展[J]. 中国生态农业学报, 2001, 9(2): 79–81.
[70] Ding Z L, Kheir A M S, Ali M G M, et al. The integrated effect of salinity, organic amendments, phosphorus fertilizers, and deficit irrigation on soil properties, phosphorus fractionation and wheat productivity[J]. Scientific Reports, 2020, 10(1): 1–13.
[71] 张密密, 陈诚, 刘广明, 等. 适宜肥料与改良剂改善盐碱土壤理化特性并提高作物产量[J]. 农业工程学报, 2014, 30(10): 91–98.
[72] Liu X Y, Yang J S, Tao J Y, et al. Elucidating the effect and interaction mechanism of fulvic acid and nitrogen fertilizer application on phosphorus availability in a salt-affected soil[J]. Journal of Soils and Sediments, 2021, 21(7): 2525–2539.
[73] Liu X Y, Yang J S, Tao J Y, et al. Integrated application of inorganic fertilizer with fulvic acid for improving soil nutrient supply and nutrient use efficiency of winter wheat in a salt-affected soil[J]. Applied Soil Ecology, 2022, 170: 104255.
[74] Liu X Y, Yang J S, Yao R J. Synergistic effects of fertilizer reduction and fulvic acid application on decreasing NaCl content and N, P availability of salinized soil[J]. Plant Nutrition and Fertilizer Science, 2021, 27(8): 1339–1350.
[75] 高珊, 杨劲松, 姚荣江, 等. 调控措施对滨海盐渍土磷素形态及作物磷素吸收的影响[J]. 土壤, 2020, 52(4): 691–698.
[76] 高婧, 杨劲松, 姚荣江, 等. 不同改良剂对滨海重度盐渍土质量和肥料利用效率的影响[J]. 土壤, 2019, 51(3): 524–529.
[77] Wu L P, Wei C B, Zhang S R, et al. MgO-modified biochar increases phosphate retention and rice yields in saline- alkaline soil[J]. Journal of Cleaner Production, 2019, 235: 901–909.
[78] Wu L P, Zhang S R, Wang J, et al. Phosphorus retention using iron (II/III) modified biochar in saline-alkaline soils: Adsorption, column and field tests[J]. Environmental Pollution (Barking, Essex: 1987), 2020, 261: 114–223.
[79] Cheng C H, Lehmann J, Thies J E, et al. Stability of black carbon in soils across a climatic gradient[J]. Journal of Geophysical Research: Biogeosciences, 2008, 113(G2): G02027, 1–10.
[80] Chowdhury M A, de Neergaard A, Jensen L S. Potential of aeration flow rate and bio-char addition to reduce greenhouse gas and ammonia emissions during manure composting[J]. Chemosphere, 2014, 97: 16–25.
[81] Cao N, Wang J W, Pang J Y, et al. Straw retention coupled with mineral phosphorus fertilizer for reducing phosphorus fertilizer input and improving cotton yield in coastal saline soils[J]. Field Crops Research, 2021, 274: 108–309.
[82] 张建兵, 杨劲松, 姚荣江, 等. 有机肥与覆盖方式对滩涂围垦农田水盐与作物产量的影响[J]. 农业工程学报, 2013, 29(15): 116–125.
[83] 张秀敏, 高日平, 康文钦, 等. 秸秆还田对盐碱地改良的研究进展[J]. 北方农业学报, 2021, 49(5): 85–92.
[84] Zhang T, Wang T, Liu K, et al. Effects of different amendments for the reclamation of coastal saline soil on soil nutrient dynamics and electrical conductivity responses[J]. Agricultural Water Management, 2015, 159: 115–122.
[85] Song Z L, Shan S D, Song Z Y. Bioavailability and interaction of Si and P in a coastal saline soil amended with pig slurry[J]. CLEAN – Soil, Air, Water, 2011, 39(3): 212–218.
[86] Shan S D, Song Z L. Effects of natural fallow and pig slurry drip irrigation on phosphorus accumulation and fractionation in a coastal saline soil[J]. Communications in Soil Science and Plant Analysis, 2010, 41(17): 2109–2121.
[87] Dey G, Banerjee P, Sharma R K, et al. Management of phosphorus in salinity-stressed agriculture for sustainable crop production by salt-tolerant phosphate-solubilizing bacteria—A review[J]. Agronomy, 2021, 11(8): 1552.
[88] Grover M, Ali S Z, Sandhya V, et al. Role of microorganisms in adaptation of agriculture crops to abiotic stresses[J]. World Journal of Microbiology and Biotechnology, 2011, 27(5): 1231–1240.
[89] 申建波, 白洋, 韦中, 等. 根际生命共同体: 协调资源、环境和粮食安全的学术思路与交叉创新[J]. 土壤学报, 2021, 58(4): 805–813.
[90] Majeed A, Muhammad Z, Ahmad H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops[J]. Plant Cell Reports, 2018, 37(12): 1599–1609.
[91] Gupta N, Sahoo D, Basak U. Evaluation ofsolubilization potential of phosphate solubilisingisolated from phyllosphere of(mangrove)[J]. African Journal of Microbiology Research, 2010, 4(3): 136–142.
[92] Rojas-Tapias D, Moreno-Galván A, Pardo-Díaz S, et al. Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize ()[J]. Applied Soil Ecology, 2012, 61: 264–272.
[93] Hafsa C, Allaoua S, Mostefa G. Solubilization of phosphate by theunder salt stress and in the presence of osmoprotectant compounds[J]. Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca Agriculture, 2011, 68(2): 4562–4571.
[94] Onireti O O, Lin C X, Qin J H. Combined effects of low-molecular-weight organic acids on mobilization of arsenic and lead from multi-contaminated soils[J]. Chemosphere, 2017, 170: 161–168.
[95] Vassilev N, Vassileva M, Nikolaeva I. Simultaneous P-solubilizing and biocontrol activity of microorganisms: Potentials and future trends[J]. Applied Microbiology and Biotechnology, 2006, 71(2): 137–144.
[96] Yi Y M, Huang W Y, Ge Y. Exopolysaccharide: A novel important factor in the microbial dissolution of tricalcium phosphate[J]. World Journal of Microbiology and Biotechnology, 2008, 24(7): 1059–1065.
[97] Ludewig U, Yuan L, Neumann G. Improving the efficiency and effectiveness of global phosphorus use: Focus on root and rhizosphere levels in the agronomic system[J]. Frontiers Agricultural Science and Engineering, 2019, 6(4): 357–365.
[98] Brundrett M C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis[J]. Plant and Soil, 2009, 320(1): 37–77.
[99] Hajiboland R, Aliasgharzadeh N, Laiegh S F, et al. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (L.) plants[J]. Plant and Soil, 2010, 331(1): 313–327.
[100] Hallett P D, Feeney D S, Bengough A G, et al. Disentangling the impact of AM fungi versus roots on soil structure and water transport[J]. Plant and Soil, 2009, 314(1): 183–196.
[101] Martin S L, Mooney S J, Dickinson M J, et al. The effects of simultaneous root colonisation by threespecies on soil pore characteristics[J]. Soil Biology and Biochemistry, 2012, 49: 167–173.
[102] Rillig M C, Mardatin N F, Leifheit E F, et al.of arbuscular mycorrhizal fungi increases soil water repellency and is sufficient to maintain water-stable soil aggregates[J]. Soil Biology and Biochemistry, 2010, 42(7): 1189–1191.
[103] Feddermann N, Finlay R, Boller T, et al. Functional diversity in arbuscular mycorrhiza–The role of gene expression, phosphorous nutrition and symbiotic efficiency[J]. Fungal Ecology, 2010, 3(1): 1–8.
[104] Smith S E, Facelli E, Pope S, et al. Plant performance in stressful environments: Interpreting new and established knowledge of the roles of arbuscular mycorrhizas[J]. Plant and Soil, 2010, 326(1): 3–20.
[105] Nedjimi B. Salt tolerance strategies ofL.: A new fodder crop for Algerian saline steppes[J]. Flora - Morphology, Distribution, Functional Ecology of Plants, 2009, 204(10): 747–754.
[106] Al Sherif E A.(L.) All., a salt-tolerant wild leguminous herb with high potential for use as a forage crop in salt-affected soils[J]. Flora - Morphology, Distribution, Functional Ecology of Plants, 2009, 204(10): 737–746.
[107] Ashraf M, Hameed M, Arshad M, et al. Salt tolerance of some potential forage grasses from cholistan desert of Pakistan[M]//Tasks for Vegetation Science. Dordrecht: Springer Netherlands, 2006: 31–54.
[108] White P J, Hammond J P. The ecophysiology of plant-phosphorus interactions[M]. Dordrecht: Springer Netherlands, 2008.
Phosphorus Availability in Saline Soil: A Review
XIAN Jingtian1,2, CHEN Xiaobing2*, WANG Shang3, ZHANG Xiaolong1, XU Gang4
(1 School of Environmental and Materials Engineering, Yantai University, Yantai, Shandong 264005, China; 2 Key Laboratory of Coastal Zone Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai, Shandong 264003, China; 3Department of Soil and Plant Microbiome, Christian-Albrechts- University of Kiel, Kiel 24118, Germany; 4 College of Geography and Tourism, Qufu Normal University, Rizhao, Shandong 276826, China)
Saline soil occupies about 3% of the global land area and it is a strategically important reserve land resource. However, salt stress not only reduces the osmotic potential of soil water and produces ion toxicity, but also inhibits the balanced absorption of plant nutrients. Soil salt especially affects plant macronutrient phosphorus (P), which greatly reduces its availability and seriously affects crop yield. Therefore, although the lack of available phosphorus is a widespread problem in all soil types, the problem of low phosphorus availability in saline soil is more prominent. Salinity and available phosphorus deficiency are two main abiotic stresses, especially for calcareous soils, plants are often exposed to both stresses at the same time. However, at this stage, the effects of salt and phosphorus are not clearly defined and systematically summarized. Therefore, this paper firstly, summarize the mechanisms of action of salinity in reducing phosphorus effectiveness, then review the mutual, integrated and interactive effects of salinity and phosphorus, and finally, summarize the current strategies for enhancing phosphorus effectiveness in saline soils and give an outlook on their research prospects, in order to provide theoretical support for the mitigation of abiotic stresses and optimal management of phosphorus elements in saline soils in the future.
Saline soil; Phosphorus availability; Mechanism; Action effects; Promotion strategy
S156.4
A
10.13758/j.cnki.tr.2023.03.003
咸敬甜, 陈小兵, 王上, 等. 盐渍土磷有效性研究进展与展望. 土壤, 2023, 55(3): 474–486.
国家自然科学基金委员会–山东省人民政府联合基金项目(U180215),国家重点研发计划项目(2019YFD1002702)和中国科学院STS项目(KFZD-SW-112-4)资助。
(xbchen@yic.an.cn)
咸敬甜(1997—),女,山东德州人,硕士研究生,主要研究方向为环境工程。E-mail: xjt18315967232@163.com
猜你喜欢
农业工程学报(2022年8期)2022-08-08
中国调味品(2022年8期)2022-08-05
农业工程学报(2022年5期)2022-06-22
中国土壤与肥料(2021年5期)2021-12-02
河南建材(2021年2期)2021-03-11
国际汉学(2017年2期)2017-01-28
工程建设与设计(2016年8期)2016-03-11
食品工业科技(2014年13期)2014-03-11
土壤与作物(2013年4期)2013-03-11
化学分析计量(2013年5期)2013-03-11
