
枣JMS2×交城5 号F1代果实性状遗传分析与优系筛选
2023-07-27 11:09:44潘依玲鲍荆凯陈万年吴翠云王玖瑞刘孟军闫芬芬
果树学报 2023年6期
潘依玲 鲍荆凯 陈万年 吴翠云 王玖瑞 刘孟军 闫芬芬


摘要:【目的】探究杂交F1代果实性状的分离特点和遗传变异,为枣杂交育种和品种选育提供理论依据。【方法】以枣JMS2×交城5 号的F1代140 个单株及亲本为试材,对果实成熟期、果实外观和内在品质等26 个描述型和数值型性状进行调查测定,并进行遗传变异分析和相关性分析,利用灰色关联度法进行果实品质综合评价。【结果】F1代果实的成熟期表现出分离,36.26%的F1株系成熟期多数在9 月下旬。F1代13 个果实的描述型性状均出现不同的性状分离,其中果实形状出现了6 个变异类型,子代果形倾向父本的倒卵圆形占比最高为35.85%,子代果形倾向母本的卵圆形占比为17.92%;果实质地出现4个变异类型,其中46.23%表现出果肉致密。果肩形状和柱头状态,分离比例依次为3∶1、1∶2∶1,符合孟德尔遗传分离规律。F1代果实大小和果实内在品质指标等13 个性状均呈现连续变异,且符合正态分布或偏正态分布,具有多基因控制的数量性状特征。果实大小、果核大小以及可食率等性状的变异系数在0.85%~32.27%之间,其中单果质量变异系数最大;果实内在品质性状变异系数在11.29%~24.44%之间,其中有机酸含量的变异系数最大。可溶性固形物含量和单核质量2 个数值型性状表现趋高遗传,单果质量、果实纵径、果实横径等11 个数值型性状均表现趋低遗传。果实大小与果核大小性状间存在极显著正相关;可食率与果实大小性状呈极显著正相关,与果核大小呈极显著负相关;可溶性糖与有机酸含量呈显著正相关,与可溶性固形物含量呈极显著正相关。【结论】杂交F1代果实成熟期倾向父本遗传,且果实形状、果实颜色、果实质地等8 个描述型性状倾向父本遗传,果肩形状和柱头状态两性状均可能是受一对和少数几对基因控制的质量性状,果实大小、果核大小、可食率及内在品质指标等13 个数值型性状为多基因控制的数量性状,初步筛选出5 个综合品质表现优良的株系。
关键词:枣;杂交后代;果实性状;遗传变异;评价
中图分类号:S665.1 文献标志码:A 文章编号:1009-9980(2023)06-1085-14
枣(Ziziphus jujuba Mill.)为鼠李科枣属植物,是起源于中国的特色优势果树,亦是栽培历史最为悠久的“五果”之一[1]。枣树适应性强,耐干旱、耐盐碱、易管理[2],具有较高的经济和生态价值。枣果实风味独特、营养价值高,具有鲜食、制干、观赏、加工[3]、药用[4]等多方面用途,深受消费者青睐。据统计中国枣树种植面积已达200 万hm2,产量达900 多万t,年产值约1000 亿元人民币,有近2000 万人口以枣业为主要经济来源[5]。
果树性状遗传变异规律是进行果树杂交育种的前提和基础,而杂交群体是果树性状遗传研究的基础。枣树花小,人工杂交难度大,且坐果率低,种仁败育严重,致使枣树有性杂交困难[6],杂交育种进程仍缓慢,有关枣树杂交群体构建和研究的报道与其他果树树种相比较少[7-8]。目前枣树遗传变异规律已在枣花[9]、叶片[10-11]、针刺[12]、果实大小[13]、果实营养品质[14]等性状展开研究,但不同亲本构建的杂交群体间性状遗传规律存在差异。枣果实性状是反映品种优劣的重要指标,也是枣杂交育种关注的重要性状指标。目前有关枣杂交群体的果实性状研究主要集中在果实大小、可溶性糖含量、可滴定酸含量等[13-16]性状上,品质指标系统性和全面性还有待加强。
笔者以雄性不育系材料JMS2 作为母本[17],交城5 号(简称J5)高抗枣疯病的骏枣优系[18]作为父本,杂交获得的140 株F1 代杂交后代及亲本作为研究材料,对果实成熟期、果实外观和内在品质等26 个描述型性和数值型性状进行调查与研究,探讨遗传变异特点和分离规律,同时利用灰色关联度分析法对F1代果实性状进行综合评价,筛选出果实品质优良的株系,以期为枣的杂交育种、品种选育及果实品质性状的QTL定位等研究奠定基础。
1 材料和方法
1.1 试验材料
母本为雄性不育JMS2,父本为骏枣优系交城5号,父母本于2013 年统一高枝嫁接,砧木为灰枣,株行距1 m×2 m,树体大小一致,留枝量相同。于2015—2016 年连续2 年采用罩网控制蜜蜂授粉技术开展控制杂交,罩网材料为网眼50 目的白色蚊帐布网纱,每网室罩亲本10~12 株。于每年现蕾期(5 月上旬)搭架罩网隔离双亲,去除网室内已开的花朵,初花期(5 月中下旬)向网内放置蜂箱饲养蜜蜂辅助授粉。每网室放置1 个蜂箱,每蜂箱放置2 个巢脾,放蜂期间每隔5~7 d 更换清水和糖水饲养蜜蜂。花期每10 d 喷施1 次0.3% K2HPO4+0.2%硼酸叶面肥增加坐果量,于当9 年月中下旬采收果实冷藏处理,翌年1 月份播种。苗期对杂交后代选用差异互补引物JSSR 311 进行杂种鉴定。2017 获得杂交后代株系,2018 年优选出140 株F1代及亲本试材高接于新疆生产建设兵团第一师十团七连果园中,砧木均为多年生骏枣,肥水管理良好,树体管理水平中等。2021年树体果实性状稳定后开展果实相关性状调查分析。于2021 年9 月1 日—10 月20 日,每隔2~4 d,观
察果实成熟情况,在树整体的果实全红占比为90%以上时进行采摘,采摘枣吊中部大小相近、且无病蟲害的果实,带回实验室放4 ℃冰箱待测。当天对果实外观品质进行调查与测量,后对其果实进行混样(每个样品包含10 个果以上),再将其放到-80 ℃备用。
1.2 果实性状的测定
参考《中国枣种质资源》对果形、果色、果肩形状、果顶形状、果点大小、果点密度、萼片状态、柱头状态、果实光滑度、果皮厚度、果实质地、果实颜色、果实汁液、果肉粗细等描述型性状进行评定(表1);评价标准参照鲜食枣果实评价标准[16],其中综合口感评分指标包含果皮厚度、果实质地、果实汁液、果肉粗细(表2)。果实成熟期以全红期采样时间作为成熟期标准。
单果质量使用电子天平对随机选取30 个果进行称质量,计算其平均值,单位为g,精确到0.01 g;果实纵径、果实横径利用游标卡尺测量,单位为mm,精确到0.01 mm;果形指数是果实纵径比果实横径;可食率利用单果质量和单核质量计算得出,计算公式:可食率(%)=(果实质量-果核质量)/果实质量×100。
果实可溶性固形物含量测定:将已混匀的果肉,利用挤汁装置将枣汁挤出,用手持折光仪测定可溶性固形物含量。可溶性糖含量采用蒽酮比色法[19]测定;可滴定酸含量采用酸碱中和法[19]测定;维生素C含量采用钼酸铵比色法[20]测定。
单核质量使用电子天平进行称质量,计算其平均值,单位为g,精确到0.01 g;果核纵径、果核横径利用游标卡尺测量,单位为mm,精确到0.01 mm;核形指数是果核纵径比果核横径。
1.3 数据处理方法
利用Excle 2019 和SPSS 26.0 对调查所得数据进行整理分析,并计算其平均值(Mean)、标准差(SD)、偏度(Skewness)、峰度(Kurtosis)、变异系数(CV)、遗传传递力(Ta)、超高亲率(HH)、低低亲率(L)、中亲值(VMP)和中亲优势率(RHm)等描述型分析参数,观察遗传变异情况;利用SPSS 26.0 绘制其相应频数分布直方图并进行正态分布检测;同时利用SPSS 26.0 进行性状间的相关性分析;最后利用Excle2019 计算等权和加权灰色关联度对其后代果实品质进行初步综合评价[21],筛选果实品质优良的杂交F1代株系。
2 结果与分析
2.1 果实性状遗传分析
2.1.1 果实成熟期性状遗传分析通过对母本JMS2 和父本交城5 号及其F1代的成熟期调查发现(表3),母本JMS2 成熟期为9 月中旬,父本成熟期在9 月下旬。F1代群体中9 月上旬成熟的植株有10 株,占比为10.99%;9 月中旬成熟的植株有24 株,占比为26.37%;9 月下旬成熟的植株有33 株,占比为36.26%;10 月上旬成熟的植株有24 株,占比为26.37%。其中F1代多数植株与两亲本成熟期相近,在9 月中旬和下旬,但主要集中在9 月下旬成熟,成熟期倾向父本遗传。2.1.2 果实描述型性状遗传分析通过对JMS2×交城5号及其F1代的果实形状、果实颜色、果肩性状、果顶性状、果点大小、果点密度、柱头状态、果面光滑度、果皮厚度、果实质地、果肉颜色、果实汁液、果肉粗细等果实描述型性状进行调查分析(表4),结果显示13个果实描述型性状出现了40个性状分离。
果实外观性状中果实形状变异类型最多,有6个变异类型,母本JMS2 表现为卵圆形、父本交城5号表现为倒卵圆形,F1代主要分离的性状表现为卵圆形、长圆形、倒卵圆形,依次占比17.92%、18.87%、35.85%。F1代中果实颜色的3 个表型中以红为主,占比55.66%;果肩形状的2 个表型中以平为主,占比76.42%;果顶形状的3 个表型中以平为主,占比58.49%;果点密度主要表现为密,占比65.09%;果点大小主要表现为中,占比59.43%;柱头状态主要表现为残存,占比55.66%;果面光滑度主要表现为光滑,占比96.23%。
果实内在性状中果实质地,母本JMS2 表现为酥脆、父本交城5 号表现为致密,F1代出现4 个变异类型,质地致密占比46.23%。F1代中的果皮厚度主要表现为中,占比72.64%;果肉颜色主要表现为浅绿,占比90.57%;果实汁液主要表现为中,占比77.36%;果肉粗细主要表现为中,占比67.92%。
在上述性状中,父母本的果肩形状表现为凸和平,F1代分离出凸和平2 个表型,且分离比例接近于3∶1,对其进行卡平方检验的结果显示χ2(df=1)=0.113,p=0.737>0.05;父母本的柱头状态均表现为残存,F1代中分离出脱落、残存、宿存等3 个表型,分离比例接近于1∶2∶1,对其进行卡平方检验的结果显示χ2(df=2)=1.377,p=0.502>0.05,果肩形状和柱头状态2个果实性状均符合孟德尔遗传规律中等位基因调控的分离情况,推测该性状可能是受一对或少数几对基因调控的质量性状。
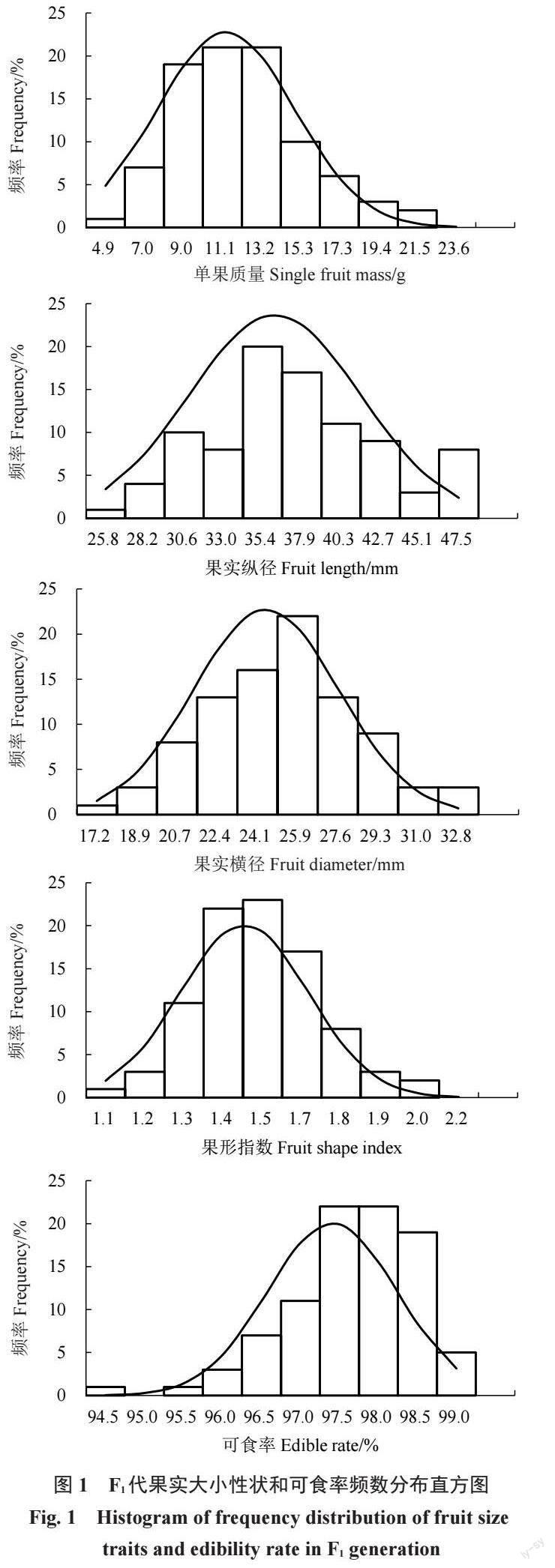
2.1.3 果实大小性状及可食率遗传分析通过对果实大小和可食率性状的频数分布直方图(图1)分析,结果表明其性状呈连续变异,符合正态分布,具有典型的数量性状遗传特征,属于微效多基因控制的数量性状。偏度是度量正态分布偏向某一侧的指标,偏度越接近0,对称越均匀;偏度为正,表明数据分布在右侧更为扩展。偏度为负,表明数据分布在左侧更为扩展[22]。单果质量和可食率的偏度值较大(表5),结合正态分布图可以看出单果质量出现明显的右偏分布,可食率出现明显的左偏分布。
对JMS2×交城5 号杂交F1代单果质量、果实纵径、果实横径、果形指数、可食率等5 个果实性状进行调查分析(表5)。结果表明果实大小性状和可食率在子代中的變异系数在0.85%~32.27%,说明果实大小性状在子代个体中存在较为广泛的分离。其中单果质量变异系数最大,为32.27%;可食率变异系数最小,为0.85%。单果质量、果实纵径、果实横径、可食率的中亲优势率均为负值,F1代平均值均小于亲中值,故推测单果质量、果实纵径、果实横径、可食率在子代中趋低遗传。但在F1代群体中果实大小性状及可食率等5 个性状均有超高亲植株出现,其中果形指数的超高亲率最高,为20.88%,其性状在F1代中具有明显的杂种优势;而单果质量的超高亲率最低,为5.49%,其性状在F1代中的杂种优势不明显。
2.1.4 果实营养性状的遗传变异分析通过对果实营养性状的频数分布直方图(图2)分析,结果表明其性状呈连续变异,符合正态分布,具有典型的数量性状遗传特征,属于微效多基因控制的数量性状。可溶性固形物和维生素C 含量的偏度值较大(表6),结合正态分布图可以看出可溶性固形物含量出現明显的左偏分布;维生素C含量出现明显的右偏分布。
对JMS2×交城5 号杂交F1 代可溶性糖、有机酸、可溶性固形物、维生素C 含量等4 个果实营养性状进行调查分析(表6),结果表明果实营养性状在子代中的变异系数在11.29%~22.44%,说明果实营养性状在F1代个体中存在较为广泛的分离。其中有机酸含量变异系数最大,为22.44%;可溶性固形物含量变异系数最小,为11.29%。除可溶性固形物含量杂种优势率表现出正值;可溶性糖、有机酸、维生素C 含量均为负值,子代平均值均小于亲中值,故推测可溶性糖、有机酸、维生素C含量在子代中趋低遗传。但在F1代群体中果实营养性状有超高亲植株出现,其中可溶性糖和可溶性固形物含量的超高亲率最高,均为16.67%,其性状在F1代具有明显的杂种优势;而有机酸含量的超高亲率最低,为3.85%,其性状在F1代的杂种优势不明显。
2.1.5 果核大小性状的遗传变异分析通过对果核大小性状的频数分布直方图(图3)分析,结果表明其性状呈连续变异,符合正态分布,具有典型的数量性状遗传特征,属于微效多基因控制的数量性状。核形指数的偏度值较大(表7),结合正态分布图可以看出核形指数出现明显的右偏分布。
对JMS2×交城5 号杂交F1代单核质量、果核纵径、果核横径、核形指数等4 个果核性状进行调查分析(表7)。结果表明果核性状在子代中的变异系数在14.52%~31.53%之间,说明果核大小性状在F1代个体中存在较为广泛的分离。其中单核质量变异系数最大,为31.53%;果核纵径变异系数最小,为14.52%。单核质量的中亲优势率是正值,果核纵径、果核横径的中亲优势率均为负值,F1代平均值均小于亲中值,故推测果核纵径、果核横径在子代中趋低遗传。但在F1代群体中果核性状有超高亲植株出现,其中单核质量的超高亲率最高,为34.07%,其性状在F1代具有明显的杂种优势;而果核纵径的超高亲率最低,为9.89%,其性状在F1 代的杂种优势不明显。
2.2 F1代果实性状的相关性分析
JMS2×交城5 号杂交F1 代果实和果核大小及果实可食率等9 个性状的相关性分析结果(表8)表明,单果质量与果实纵径、果实横径存在极显著的正相关关系,且相关系数均在0.700 以上,说明以上性状具有较强相关性;果实纵径与果实横径、果形指数具有显著的正相关关系,而果实横径与果形指数具有极显著的负相关关系。在果核大小性状的相关性分析结果中看出,单核质量与果核纵径、果核横径存在极显著的正相关关系;果核纵径与核形指数存在极显著的正相关关系,而果核纵径与核形指数存在极显著的负相关关系。在果实大小与果核大小性状相关性分析结果中看出,果实的单果质量、果实纵径、果实横径分别与单核质量、果核纵径、果核横径存在极显著的正相关关系,其中果实纵径与果核纵径的相关系数最高,为0.677;果形指数与果核纵径、核形指数具有极显著的正相关关系,同时核形指数与果实纵径也存在极显著的正相关关系。在可食率与果实大小、果核大小性状相关性的分析结果中看出,可食率与单果质量、果实纵径、果实横径存在极显著正相关关系,而与单核质量、果核纵径、核形指数存在极显著负相关关系。
从F1代果实营养性状相关性分析结果(表9)可以看出,果实的可溶性糖与有机酸含量存在显著正相关关系;可溶性糖与可溶性固形物含量存在极显著正相关关系;可溶性固形物与维生素C含量存在显著正相关关系。
2.3 F1代果实品质的综合评价
笔者在本研究中利用等权和加权灰色关联度分析法对果实的单果质量、可溶性糖含量、有机酸含量、维生素C含量、可食率以及综合口感6 个重要性状进行初步综合评价。在选取的果实品质性状中有机酸含量采用平均值,其他性状均采用最高值作为果实品质的“理想类型”构造数据。根据不同性状的重要程度以及变异程度分别给予权重值,依次是0.20、0.25、0.15、0.10、0.05、0.25。在F1代中筛选出了性状表现优良的5 个优系(表10),分别是J20、J47、J52、J56、J93,其加权关联度依次是1.297、0.842、0.989、0.681、0.717。加权关联度结合筛选出F1代的5 个优株果实品质性状的表现见表11,可以看出J20作为鲜食优系时,果实的综合品质表现要优于父母本和其他子代,筛选出的优系果实见图4。
3 讨论
3.1 F1代果实成熟期与描述型性状的遗传变异分析
果实外观和口感是影响消费者购买的主要因素,研究果实外观和内在的描述型性状遗传特点,对于选育外观好、口感佳、商业价值高的优良品系具有重大意义,因此笔者对JMS2×交城5 号的F1代果实外观和内在的描述型性状进行遗传变异分析,调查发现13个果实描述型性状出现了40个性状分离,说明杂交F1代描述型性状变异丰富,具有选育优良枣种质的潜力。其中F1代的果实形状、果实颜色、果肩形状、果顶形状、果点密度、柱头状态、果面光滑度、果实质地8 个描述型性状占比最多的表型与父本的表型一致,推测该性状更倾向于父本遗传。而果点大小、果皮厚度、果肉颜色、果实汁液、果肉粗细等5个描述型性状占比最多的表型同两亲本表型一致,表明这些性状呈现的分离比例较低,F1代性状表现多与亲本一致。果实成熟期是果实性状的重要指标,本研究发现,F1代群体果实的成熟期多数接近两亲本,这一研究结果与刘振中等[23]在苹果上研究结果相似,但就两亲本而言,F1代植株主要与父本成熟期相近,果实成熟期倾向父本遗传,推测在枣树上父本的果实成熟期具有较强的遗传力,但还需不同枣群体进一步验证。
研究发现枣F1代果实描述型性状及成熟期会出现不同程度分离,但多数性状出现明显的偏父本遗传,父本的性状对杂交F1群体影响更大,因此,在今后的枣杂交育种亲本选配工作中,应加强父本的选择和选配,选择果实大小、果实形状、果实质地和果面光滑度等性状优良的父本进行选配,进而获得符合选育目标的大量变异类型,提高育种工作效率。
同时,研究中发现F1代枣果实的果肩形状和柱头状态2 个描述型性状的分离比例符合孟德尔遗传规律,推测可能是受一对或少数几对基因控制的质量性状。结合柱头状态在父母本上的表现型和在子代中的分离情况进一步推测,该性状在受显隐性基因控制的杂合体中柱头状态表现为残存,完全显性或隐性基因的柱头状态表现为宿存或脱落,但具体是显性还是隐性基因控制还有待进一步研究。
3.2 F1代果实品质数值型性状的遗传变异分析
枣树遗传背景复杂,基因高度杂合[6],故在杂交后代中,会出现微效多基因分离,也会出现隐性或多微效基因组合,即形成基因的加性和非加性效应,能表现出某些优良性状,但劣变率普遍较高。笔者在本研究中通过对JMS2×交城5 号的F1 代枣果实大小、果实可食率、果实营养品质、果核大小等13 个数值型果实品质性状调查分析,发现在F1代中均呈现连续变异,表现出多基因控制的数量性状特征,这与谢欢等[15]对冬枣×金丝4 号杂交后代的研究结果相似,且在枇杷[24]、苹果[25]、葡萄[26]等果树上也有相似规律。果实品质性状呈正态分布或偏正态分布,推测可能由于杂交育种过程非加性效应解体,以及栽培过程中气候与环境条件等外界因素的影响。F1代枣果实大小性状、果实可食率、果实营养品质、果核大小等13 个果实性状变异系数在0.85%~32.27%之间,说明其F1代的果实性状能广泛分离,其中单果质量变异系数最大,各性状均表现不同程度的离散分布,部分株系出现超亲极端值,这一现象有利于从杂交F1代群体中筛选出优异种质资源[27-30]。F1代果实的可溶性固形物含量和单核质量的均值高于中亲值,杂种优势率均为正值,分别为5.56%、28.81%;且F1 代均有超高亲植株出现,超高亲率分别为16.67%、34.07%,表现出趋高遗传倾向和较强杂种优势。可溶性固形物含量的趋高变异意味着更容易选育高可溶性固形物含量品种,本研究结果与谢欢等[15]在冬枣×金丝4 号的枣杂交后代中的可溶性固形物含量遗传倾向相同,但单核质量遗传倾向研究结果与之表现出差异,推测是由于不同的杂交群体性状间的遗传规律存在一定相似性和差异性,故仍需要用不同的杂交群体去揭示枣的遗传变异规律。除上述两个性状外,其他果实大小、果实可食率、果实营养品质、果核大小等性状均表现出趋小变异,这一现象主要是由这些性状均受微效多基因控制、亲本品种均具有基因累加效应和互作效应、随着有性过程非加性效应解体而导致的,在其他果树上也有相似的报道[26-27]。
3.3 F1代果实性状综合评价与优系筛选
果实品质是由众多果实性状相互作用的结果,而果实性状形成是多基因控制的数量性状和多种因素相互作用的结果。笔者通过对果实大小、果实可食率、果实营养品质、果核大小等果实性状的相关性分析,发现果实大小与果核大小性状间存在极显著正相关性,说明枣果实越大,果核越大;可食率与果实大小性状呈极显著正相关性,与果核大小呈极显著负相关;可溶性糖与有机酸含量显著正相关,与可溶性固形物含量极显著正相关,这一结果与前人研究结果一致[15-16]。利用JMS2×交城5 号的F1代获得的多个果实品质性状指标对其果实品质进行综合评价,评价结果会因信息重叠而出现偏差。而采用灰色关联度分析法对鲜食枣果实品质进行分析,可以摆脱果实品质指标多、选择难的问题,达到全面评价的目的。目前张梅等[31]和李慧等[21]分别对骏枣和鲜食枣种质应用灰色关联分析法进行筛选并取得了较好效果。笔者在本研究中参照枣树鲜食品种的育种目标,利用灰色关联分析法,选取6 个对果实品质较为重要的性状指标,筛选出5 个综合性状优良的优系,分别是J20、J47、J52、J56、J93,其中J20 鲜食的果实品质表现要明显优于其他子代,可作为鲜食品种选育,而其他4 个优系可以考虑作为干鲜兼制品种选育。笔者通过果实性状评价分析,初步筛选出5个果实品质综合性状表现优良的种质,但影响枣果实品质的性状还有很多,后续将会增加制干品种评价指标和其他影响枣优株综合评价的指标,以期加快新种质筛选进程。
4 结论
本研究結果表明,杂交F1代果实成熟期倾向父本遗传,且26 个果实品质性状均呈现不同程度性状分离。其中果实形状、果实颜色、果实质地等8 个描述型性状倾向父本遗传;果肩形状和柱头状态2个性状均可能是受一对或少数几对基因控制的质量性状。果实大小和果核大小、可食率及果实内在品质等13个数值型性状为多基因控制的数量性状。初步筛选出5 个综合品质表现优良的株系。
参考文献References:
[1] 刘孟军,王玖瑞,刘平,赵锦,赵智慧,代丽,李宪松,刘志国. 中国枣生产与科研成就及前沿进展[J]. 园艺学报,2015,42(9):1683-1698.
LIU Mengjun,WANG Jiurui,LIU Ping,ZHAO Jin,ZHAO Zhihui,DAI Li,LI Xiansong,LIU Zhiguo. Historical achievementsand frontier advances in the production and research of Chinesejujube (Ziziphus jujuba) in China[J]. Acta Horticulturae Sinica,2015,42(9):1683-1698.
[2] LIU M J,WANG J R,WANG L L,LIU P,ZHAO J,ZHAO Z H,YAO S R,ST?NIC? F,LIU Z G,WANG L X,AO C W,DAIL,LI X S,ZHAO X,JIA C X. The historical and current researchprogress on jujube-a superfruit for the future[J]. HorticultureResearch,2020,7(1):119.
[3] 毕金峰,于静静,白沙沙,王沛,丁媛媛. 国内外枣加工技术研究现状[J]. 农产品加工(学刊),2010(2):52-55.BI Jinfeng ,YU Jingjing ,BAI Shasha ,WANG Pei ,DING
Yuanyuan. Research status of jujube processing technology athome and abroad[J]. Academic Periodical of Farm Products Processing,2010(2):52-55.
[4] 郭盛,段金廒,唐于平,钱大玮. 中国枣属药用植物资源化学研究进展[J]. 中国现代中药,2012,14(8):1-5.
GUO Sheng,DUAN Jinao,TANG Yuping,QIAN Dawei. Progresson resource chemistry of the medicinal plants in genusZiziphus[J]. Modern Chinese Medicine,2012,14(8):1-5.
[5] 刘孟军,王玖瑞. 新中国果树科学研究70 年:枣[J]. 果树学报,2019,36(10):1369-1381.
LIU Mengjun,WANG Jiurui. Fruit scientific research in NewChina in the past 70 years:Chinese jujube[J]. Journal of FruitScience,2019,36(10):1369-1381.
[6] 马庆华,续九如,王贵禧,姚立新. 枣树杂交育种研究进展[J].中国农学通报,2008,24(11):174-178.
MA Qinghua,XU Jiuru,WANG Guixi,YAO Lixin. Progress onhybrid breeding of Chinese jujube (Ziziphus jujuba Mill.) [J].Chinese Agricultural Science Bulletin,2008,24(11):174-178.
[7] 刘孟军,王玖瑞,刘平,林敏娟,肖京,刘志国,孙学超. 枣树免去雄杂交育种的设计与实践[J]. 园艺学报,2014,41(7):1495-1502.
LIU Mengjun,WANG Jiurui,LIU Ping,LIN Minjuan,XIAOJing,LIU Zhiguo,SUN Xuechao. Design and practice of emasculation-free cross breeding in Chinese jujube[J]. Acta HorticulturaeSinica,2014,41(7):1495-1502.
[8] 闫芬芬. 枣规模化杂种创制技术体系的建立及重要性状遗传分析[D]. 保定:河北农业大学,2017.
YAN Fenfen. Establishment of technical system for large- scalehybrid creation of jujube and genetic analysis of important characters[D]. Baoding:Hebei Agricultural University,2017.
[9] 杨植,张川疆,王振磊,闫芬芬,吴翠云,王玖瑞,刘孟军,林敏娟. 枣和酸枣杂交F1 代花性状分离规律与混合遗传分析[J].分子植物育种,2023,21(1):231-242.
YANG Zhi,ZHANG Chuanjiang,WANG Zhenlei,YAN Fenfen,WU Cuiyun,WANG Jiurui,LIU Mengjun,LIN Minjuan. Segregationand genetic mixing analysis of flower traits in the F1 progenyofZ. jujuba Mill. ×Z. acido jujuba Cheng et Liu[J]. MolecularPlant Breeding,2023,21(1):231-242.
[10] 鮑荆凯,王艳,潘依玲,卢登洋,闫芬芬,吴翠云. 枣杂交后代叶片表型性状的遗传规律分析[J/OL]. 分子植物育种,2021 :1-17. [2021-12-31]. https://kns.cnki.net/kcms/detail/46.1068.s.20211231.0928.006. html.
BAO Jingkai,WANG Yan,PAN Yiling,LU Dengyang,YANFenfen,WU Cuiyun. Analysis of inheritance of leaf phenotypiccharacters in jujube hybrid progeny[J/OL]. Molecular PlantBreeding,2021:1- 17. [2021- 12- 31]. https://kns.cnki.net/kcms/detail/46.1068.S.20211231.0928.006.html.
[11] 仇倩倩,夏怡蕾,鲍荆凯,闫敏,杨植,王程成,闫芬芬,吴翠云. 基于叶片表型性状的枣杂交F1 代遗传规律分析[J/OL]. 分子植物育种,2021:1-22. [2021-11-22]. https://kns. cnki. net/kcms/detail/46.1068.S.20211122.1348.004. html.
QIU Qianqian,XIA Yilei,BAO Jingkai,YAN Min,YANG Zhi,WANG Chengcheng,YAN Fenfen,WU Cuiyun. Study on thegenetic law of jujube hybrid F1 generation based on leaf phenotypictraits[J/OL]. Molecular Plant Breeding,2021:1-22. [2021-11- 22]. https://kns.cnki.net/kcms/detail/46.1068.S.20211122.13-48.004.html.
[12] 齐靖,董祯,申连英,毛永民,李艳辉,刘杰,王晓玲. 枣针刺长度的数量性状位点定位与分析[J]. 园艺学报,2009,36(6):807-813.QI Jing,DONG Zhen,SHEN Lianying,MAO Yongmin,LI Yanhui,
LIU Jie,WANG Xiaoling. Analysis of QTL for needlelength in Chinese jujube[J]. Acta Horticulturae Sinica,2009,36(6):807-813.
[13] 仇倩倩.‘JMS2ב邢16杂交后代高密度遗传图谱构建及果实大小相关性状的QTL 定位[D]. 阿拉尔:塔里木大学,2021.
QIU Qianqian. High-density genetic map construction and QTLmapping of fruit size-related traits of‘JMS2בXing16population[D]. Alar:Tarim University,2021.
[14] 夏怡蕾.‘JMS2ב邢16杂交后代糖酸性狀遗传规律及品质性状QTL 定位[D]. 阿拉尔:塔里木大学,2022.
XIA Yilei. Genetic law of sugar and acid traits and QTL mappingof quality traits in Chinese jujube hybrid progenies[D].Alar:Tarim University,2022.
[15] 谢欢,王中堂,李明玥,李新岗. 枣杂交后代果实性状遗传分析[J]. 经济林研究,2022,40(2):125-134.
XIE Huan,WANG Zhongtang,LI Mingyue,LI Xingang. Geneticanalysis of fruit characters in hybrid progeny of Chinese jujube[J]. Non-Wood Forest Research,2022,40(2):125-134.
[16] 袁野,胡兰,刘平,刘孟军.‘蜂蜜罐枣实生后代果实性状变异分析与评价[J]. 植物遗传资源学报,2018,19(3):539-545.
YUAN Ye,HU Lan,LIU Ping,LIU Mengjun. Evaluation offruit character in naturally-pollinated progeny ofZiziphus jujubaMill.‘Fengmiguan[J]. Journal of Plant Genetic Resources,2018,19(3):539-545.
[17] 王玖瑞,刘玲,刘孟军,周俊义. 枣树雄性不育新种质的获得[J].园艺学报,2006,33(2):374-377.
WANG Jiurui,LIU Ling,LIU Mengjun,ZHOU Junyi. Acquirementof new male sterile germplasm of Chinese jujube[J]. ActaHorticulturae Sinica,2006,33(2):374-377.
[18] 肖京,杨艳荣,赵锦,刘孟军. 骏枣不同株系间的枣疯病抗性多样性[J]. 中国农业科学,2013,46(23):4977-4984.
XIAO Jing,YANG Yanrong,ZHAO Jin,LIU Mengjun. Diversityof resistance to jujube witchesbroom disease among thestrains ofZiziphus jujuba Mill. Junzao[J]. Scientia AgriculturaSinica,2013,46(23):4977-4984.
[19] 高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006:144-147.
GAO Junfeng. Experimental guidance for plant physiology[M].Beijing:Higher Education Press,2006:144-147.
[20] 李军. 钼蓝比色法测定还原型维生素C[J]. 食品科学,2000,21(8):42-45.
LI Jun. Study on molybdenum blue method of L- VC test byspectrometry[J]. Food Science,2000,21(8):42-45.
[21] 李慧,魏天军. 基于主成分和灰色关联度分析的鲜食枣果实品质评价[J]. 经济林研究,2021,39(1):60-67.
LI Hui,WEI Tianjun. Evaluation of fruit quality of fresh-eatingjujube based on principal component and gray correlation analysis[J]. Non-Wood Forest Research,2021,39(1):60-67.
[22] 刘畅,张经常,唐斯旸,黄烁,冯章丽,卜海东,顾广军,张武杰,于文全,刘延杰,叶万军. 秋子梨杂种后代叶片性状的遗传倾向研究[J]. 中国果树,2019(5):35-40.
LIU Chang,ZHANG Jingchang,TANG Siyang,HUANG Shuo,FENG Zhangli,BU Haidong,GU Guangjun,ZHANG Wujie,YU Wenquan,LIU Yanjie,YE Wanjun. Study the genetic tendencyof leaf traits in hybrid progeny ofPyrus ussunensis[J].China Fruits,2019(5):35-40.
[23] 刘振中,樊红科,高华,赵政阳. 苹果杂交后代果实性状遗传变异分析[J]. 北方园艺,2012(5):5-8.
LIU Zhenzhong,FAN Hongke,GAO Hua,ZHAO Zhengyang.Analysis on the genetic variation of fruit characters in apple hybrids[J]. Northern Horticulture,2012(5):5-8.
[24] 赵崇斌,郭乙含,李舒庆,徐红霞,黄天启,林顺权,陈俊伟,杨向晖. 宁海白×大房枇杷F1 杂交群体果实性状的相关性及遗传分析[J]. 果树学报,2021,38(7):1055-1065.
ZHAO Chongbin,GUO Yihan,LI Shuqing,XU Hongxia,HUANG Tianqi,LIN Shunquan,CHEN Junwei,YANG Xianghui.Correlation and genetic analysis of fruit traits in F1 hybridpopulation of loquat generated from Ninghaibai × Dafang[J].Journal of Fruit Science,2021,38(7):1055-1065.
[25] 刘志,伊凯,王冬梅,杨巍,杨锋,张景娥. 富士杂交后代果实内在品质性状的遗传[J]. 果树学报,2004,21(2):95-102.
LIU Zhi,YI Kai,WANG Dongmei,YANG Wei,YANG Feng,ZHANG Jinge Studies on the fruit internal characteristics inheritancetrends of Fuji apple variety crossed progenies[J]. Journalof Fruit Science,2004,21(2):95-102.
[26] 刘政海,董志刚,李晓梅,谭敏,杨镕兆,杨兆亮,唐晓萍.‘威代尔与‘霞多丽葡萄杂交F1 代果实性状遗传倾向分析[J].果树学报,2020,37(8):1122-1131.
LIU Zhenghai,DONG Zhigang,LI Xiaomei,TAN Min,YANGRongzhao,YANG Zhaoliang,TANG Xiaoping. Inheritancetrend of fruit traits in F1 progenies of‘Vidaland‘Chardonnayof grape[J]. Journal of Fruit Science,2020,37(8):1122-1131.
[27] 白冰. 砂梨種质资源的调查与杂交后代果实性状的遗传特性研究[D]. 南京:南京农业大学,2017.
BAI Bing. Evaluation ofPyrus pyrifolia germplasm and the inheritanceof fruit traits in pear crossing progenies[D]. Nanjing:Nanjing Agricultural University,2017.
[28] 武晓红,景晨娟,陈雪峰,赵习平,袁立勇,梁爽,张宪成,唐焕英,李立颖.‘金太阳与‘串枝红杏正反交后代果实性状的遗传倾向研究[J]. 江西农业学报,2018,30(10):13-18.
WU Xiaohong,JING Chenjuan,CHEN Xuefeng,ZHAO Xiping,YUAN Liyong,LIANG Shuang,ZHANG Xiancheng,TANGHuanying,LI Liying. Studies on genetic tendency of fruit charactersin F1 generation of reciprocal crosses between apricot cultivars‘Jintaiyangand‘Chuanzhihong[J]. Acta AgriculturaeJiangxi,2018,30(10):13-18.
[29] 陆琦文. 枣实生后代主要性状遗传变异分析及优异种质筛选[D].阿拉尔:塔里木大学,2020.
LU Qiwen. Genetic variation analysis and excellent germplasmscreening of jujube seedings[D]. Alar:Tarim University,2020.
[30] CHO K H,HUR Y Y,NAM J C,PARK S J,KIM S H,HAN JH,LEE H C,JO S H. Evaluation of major fruit traits in progeniesderived from a cross between‘Tano Redand‘Ruby Seedlessgrapevines(Vitis spp. )[J]. Korean Journal of Breeding Science,2017,49(4):324-333.
[31] 张梅,王利娜,王姝婧,杨智鹏,马路婷,刘伟峰,魏喜喜,李建贵. 基于层次-关联度的新疆骏枣品质性状分析及综合评价[J].中南林业科技大学学报,2022,42(1):78-85.
ZHANG Mei,WANG Lina,WANG Shujing,YANG Zhipeng,MA Luting,LIU Weifeng,WEI Xixi,LI Jiangui. Analysis andcomprehensive evaluation of Xinjiang Jun jujube quality by hierarchy-relation analysis[J]. Journal of Central South Universityof Forestry & Technology,2022,42(1):78-85.
猜你喜欢
石油沥青(2021年4期)2021-10-14 08:50:44
世界科学技术-中医药现代化(2021年10期)2021-03-02 05:52:06
海南医学(2020年1期)2020-01-18 02:35:28
电子制作(2019年24期)2019-02-23 13:22:18
广西林业科学(2016年3期)2016-03-16 05:43:34
广西林业科学(2016年3期)2016-03-16 05:43:22
中国康复理论与实践(2015年10期)2015-12-24 05:42:44
中国教育技术装备(2015年19期)2015-03-01 02:43:07
中国工程咨询(2015年2期)2015-02-14 02:59:26
食品工业科技(2014年5期)2014-03-11 18:14:08
