
非耕作生境对相邻耕地步甲和蜘蛛分布影响的差异性*
2023-07-24 02:48边振兴吴佳璇杨玉静王楚翘张宇飞
中国生态农业学报(中英文) 2023年7期
边振兴,吴佳璇,杨玉静,王楚翘,张宇飞
(沈阳农业大学土地与环境学院/辽宁省自然资源厅耕地立体保护与监测重点实验室 沈阳 110866)
农业景观的生物多样性保护是未来农业可持续发展的重要基础,对生态系统服务功能提升有重要作用。以往研究表明,农业集约化是导致耕地系统中天敌多样性大量丧失和生物害虫控制功能降低的关键因素[1]。为了应对这种生物多样性和相应的生态系统服务下降,农业环境计划(AES)和欧盟的共同农业政策(CAP)鼓励农民留出一定比例的土地建立非耕作生境,提升耕地系统中的生物多样性。欧洲的生态学家认为农业景观中非耕作生境对相邻耕地生物多样性具有重要的保护和维持功能[2]。与耕地内部相比,非耕作生境通常具有提供较高生物多样性的潜力,这会直接影响耕地系统景观结构和耕地系统的稳定性,因此非耕作生境被认为是生物多样性保护的关键区域。耕地边缘通常具有复杂的植被结构,它们作为连接相邻两个生境的通道,不仅有相邻两个生境的共同生物种群,而且有其独特的物种[3]。并且由于溢出效应明显,非耕作生境内部的生物会通过耕地边缘向耕地内部不断移动,导致耕地边缘表现出明显的正效应,即比相邻的生境具有更为优良的特性,如生物多样性更丰富[4]。此外,在农业景观中,非耕作生境类型通常是决定其有利于天敌多样性的关键因素,不同非耕作生境类型所维持的天敌类群也存在较大的差异。
非耕作生境可以将碎片化、孤立或残留斑块连接成一个完整的生态区域,这有利于景观中的生物迁移,从而增加基因交换以及不同资源斑块的选择和利用,以保持种群稳定[5]。但由于物种在取食类型、移动能力、栖息地需求存在着一定的差异,导致它们对不同类型生境的响应模式明显不同[6]。例如,研究认为森林生境植被类型更为丰富,且人为干扰相较于其他生境更少,更适合大部分群落栖息[7]。耕地以农作物为主,植被类型单一,人为干扰较大,大大降低了生物多样性;相较于耕地,复合生境拥有较多植被类型,能为生物群落提供更多的栖息条件。以往研究虽然全面丰富,但大多数研究只针对特定的生境类型,忽视了非耕作生境对相邻耕地边缘和耕地内部的作用。
步甲(Carabidae)和蜘蛛(Araneida)是地表节肢动物天敌类重要组成[8],对害虫的生物控制服务起着重要的作用。研究认为步甲对生境的变化十分敏感[9],不同生境间步甲种类会存在较大差别[10],因此步甲常作为指示环境和生物多样性变化的重要指标[11],检测耕地系统环境变化情况。蜘蛛作为耕地系统的重要组成部分,对害虫有重要的调控作用。近年来国内外学者针对蜘蛛生态调控进行了系统性研究[12],分析了不同管理类型、不同生境条件对蜘蛛群落结构及多样性的影响,研究认为蜘蛛群落多样性与周围环境及人为干扰密切相关[13]。由于不同类群天敌的生物学特性有所差异,目前对于步甲和蜘蛛在不同生境溢出效应机制仍未达成统一结论,应进一步探讨不同类型生境天敌的活动规律[14]。
基于此,本文选取辽宁省昌图县为研究区,重点研究了非耕作生境对耕地边缘及耕地内部步甲和蜘蛛多样性的影响,旨在回答以下问题:1)不同生境内步甲和蜘蛛群落分布特征;2)非耕作生境对相邻耕地边缘及耕地内部步甲和蜘蛛多样性的影响差异。为优化耕地边缘生境、促进天敌的多样性、提高生物害虫防控效率、维持耕地生态系统结构稳定提供理论依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于辽宁省最北部,松辽平原南端的昌图县(123°32′~124°26′E、42°23′~43°29′N),属温带大陆性季风气候,气候温和,四季分明,年降水量655 mm,降水年内分布不均。县域面积总面积4317 km2,耕地面积约占全县面积78.7%,主要种植作物为玉米(Zea mays),是典型的东北旱作农业区。昌图县地形地貌自东向西由低山丘陵向平原过渡,东部为低山丘陵区,适宜种植果园、林木等;中部为平原区,适宜发展农业;西部为辽河冲积平原区,区内水利条件优越,地势平坦;西北部为风沙区,受内蒙古地区风沙影响较大,适宜种植经济作物以及发展林牧业。本研究结合研究区2019 年土地利用现状数据及遥感影像,利用ArcGIS10.2 计算提取景观特征、耕地面积等信息,分析农业景观中邻近不同非耕作生境的耕地,选择有代表性的11 个样区(图1)。

图1 研究区土地利用类型及采样区位置分布图Fig.1 Land use type and location distribution of sampling units in the study area
1.2 试验设计
在耕地系统中,耕地与邻近耕地的非耕作生境之间会形成过渡带,交接、关联两个不同生态系统,学者们通常把这种不同于两侧基质或用地类型的狭长地带称作耕地边缘[15]。本研究共选取了11 块采样区,根据耕地周围生境(2 块果园、3 块原生草原、3块其他林地、3 块乔木林地),可分为8 种生境类型,划分方式及特征如表1 所示。在非耕作生境内部及相邻耕地内部和耕地边缘分别进行采样,耕地内部设置5 个采样点,在非耕作生境、耕地边缘各设置3个采样点,每个样点设置3 个陷阱,采样单元及陷阱瓶分布如图2 所示。

表1 研究区生境类型的划分方式、数量及特征Table 1 Division,quantity and characteristics of habitat types in the study area
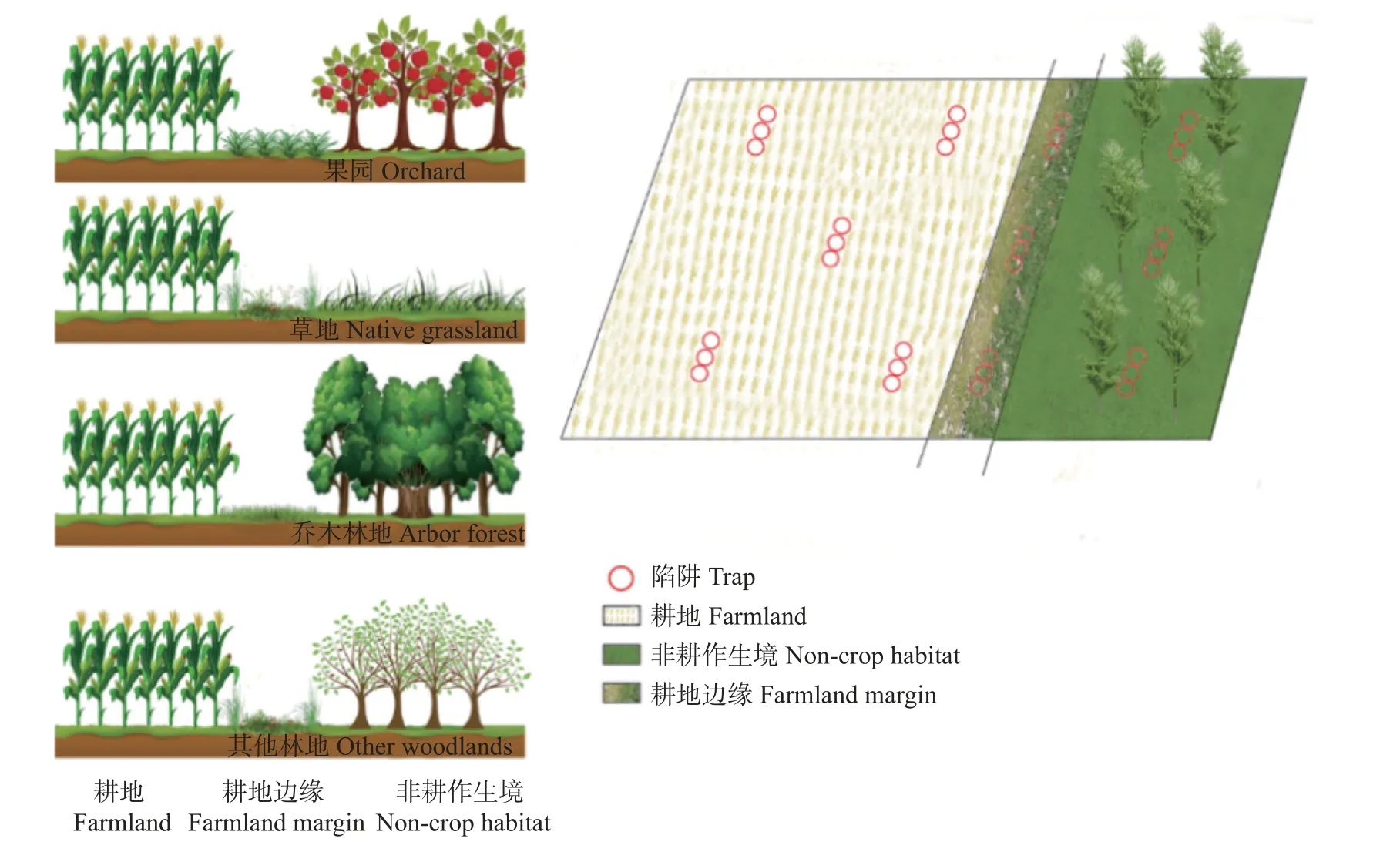
图2 不同类型耕地-耕地边缘-非耕作生境(非耕作生境分为果园、草地、其他林地和乔木林地)的结构布局和每个样品区内的陷阱布局Fig.2 Structural layout of different types of farmland -farmland margin -non-crop habitat (non-crop habitat includes orchard,native grassland,other woodlands and arbor forest) and specific method of trap layout in each sample area
1.3 生物取样及调查
1.3.1 步甲和蜘蛛取样
使用陷阱法采集步甲和蜘蛛,采样时间为2021年8−9 月。在每个采样单元耕地内部设置5 个采样点,在非耕作生境、耕地边缘各设置3 个采样点,每个样点设置3 个陷阱,共计布置363 个,陷阱间隔均大于10 m,如图2 所示。利用手持GPS 确定并记录各样点位置、编号。
将上口径9.5 cm、下口径4.5 cm、高11 cm、容积为500 mL 的PP 塑料杯埋入土壤,杯口与地表齐平,在杯中倒入150~200 mL 浓度为20%的乙二醇溶液及1~2 滴洗涤剂,并将防雨罩用三根铁丝固定插入陷阱上方1~2 cm 处。陷阱在野外放置6 d 后收回。将捕获的地表节肢动物放入PE 瓶中,并倒入可没过瓶中地表节肢动物的酒精(75%)进行保存。在实验室使用蔡司体式显微镜(ZEIS:Stemi 2000-C),并参照《中国土壤动物检索图件》对地表节肢动物进行鉴别分类。其中,蜘蛛鉴别到科,步甲鉴别到种。
1.3.2 草本植被类型调查
对非耕作生境内草本植物类型开展调查,在非耕作生境和耕地过渡带(即耕地边缘)动物陷阱周围随机布设6 个0.5 m×0.5 m 草本植物采集样方,记录样方内草本植种名、株(丛)数、平均高度(AH)和盖度(AC)。
1.4 数据分析
为解决取样期间陷阱破坏造成的取样不均,首先将步甲和蜘蛛的多度数据进行标准化。合并每个取样点3 个陷阱瓶的数据取平均值,用于分析不同类型生境对耕地天敌的影响。地表节肢动物多样性指数利用PAST 软件进行计算,选取个体数(G)、物种数(S)、Shannon 多样性指数(H)、Pielou 均匀度指数(E) 4 个指数[16]进行群落多样性描述。采用单因素方差分析(One-way ANOVA)明确不同生境对天敌多样性的影响,比较具有显著影响的天敌多样性指数在不同生境间的差异,方差分析通过SPSS 25 软件完成。合并同一类型生境陷阱数据,对不同生境地表节肢动物个体数进行层次聚类分析(Hierarchical Clustering),以明确不同生境地表节肢动物组成的相似性。采用基于CNESS (Chord Normalized Expected Species Shared)相似系数的非度量多维尺度法(Nonmetric Multi-Dimensional Scaling,NMDS)分析比较不同生境类型步甲和蜘蛛群落结构的相似性,反映生物类群因环境造成的异质性。合并植被样方数据,用PAST 软件计算多样性指数,通过主成分分析(Principal Component Analysis,PCA)进行降维,将原植被数据重新组合成一组新的互相无关的几个综合数据变量,并使综合变量尽可能多地反映原植被数据信息。采用冗余分析法(Redundancy Analysis,RDA)定量研究耕地边缘草本植被结构因子与步甲和蜘蛛间的关系。冗余分析通过CANOCO5 进行计算。
2 结果与分析
2.1 步甲和蜘蛛群落组成及群落结构
在研究区耕地内部、耕地边缘以及非耕作生境共捕获步甲1921 只、蜘蛛567 头(表2),捕获步甲分属于10 个种。其中,耕地内部捕获步甲372 只,耕地边缘捕获976 只,非耕作生境捕获573 只;优势种为普通暗黑步甲(Amara brevicollis)和蠋步甲(Dolichus halensis),占捕获步甲总个体数的25.8%和25.0%;常见种包括广屁步甲(Pheropsophus occiptalis)、大暗黑步甲(Amara majuscula)和黄斑青步甲(Chlaenius micans),分别占捕获步甲总个体数的12.2%、11.3%和10.8%。共捕获蜘蛛567 头,分属于3 个科,其中,耕地内部捕获89 只,耕地边缘捕获279 只,非耕作生境捕获199 只;优势科为长奇盲蛛科(Phalangiidae)和漏斗蛛科(Agelena labyrinthica),占捕获蜘蛛总个体数的48.5%和44.8%。

表2 不同生境地表节肢动物类型及数量统计表Table 2 Statistics of arthropod types and numbers in different habitats
聚类分析结果表明(图3),8 种类型的生境可分为两类,果园生境(W2)与邻近果园的耕地生境(W1)为一类,其他类型生境为一类。由结果可知,草地(W4)、其他林地(W6)、乔木林地(W8)及其相邻耕地具有相似的步甲和蜘蛛群落结构,其他林地(W6)内步甲和蜘蛛群落的组成与乔木林地(W8)及邻近乔木林地耕地(W7)最接近,而果园(W2)和相邻果园耕地(W1)步甲和蜘蛛群落的组成与其他生境存在明显的差异性。

图3 8 种生境类型步甲和蜘蛛层次距离聚类分析个体数热点图Fig.3 Hotspot map of cluster analysis of carabid and spiders in different habitat types
2.2 非耕作生境对耕地内部步甲和蜘蛛的影响
通过单因素方差分析可知(图4):不同非耕作生境类型对耕地内部步甲和蜘蛛的个体数、物种数和多样性影响显著(P<0.001),对于均匀度指数影响不显著(P=0.125)。其中,草地(W4)内部步甲的个体数、物种数和多样性显著影响其他非耕生境(P=0.048),乔木林地(W8)内部蜘蛛的个体数和物种数显著高于其他生境(P<0.001),其他林地(W6)内部蜘蛛的多样性影响显著高于其他生境(P=0.08)。相较于其他耕地内部,步甲和蜘蛛在相邻果园耕地(W1)内部维持着较高的个体数、物种数和多样性。

图4 非耕作生境内部及邻近农田内部步甲(A)和蜘蛛(B)多样性分析Fig.4 Diversities of carabid beetle (A) and spider (B) in non-crop habitats and adjacent farmlands
NMDS 分析结果显示(图5),4 种类型非耕作生境及其相邻耕地内步甲和蜘蛛群落分布差异显著,Stress 值为0.1163,表明排序结果良好。其他林地及邻近耕地与乔木林地及邻近耕地内群落具有较高的相似性。草地及邻近耕地与其他林地及邻近耕地之间分离较为明显,表明其内部物种组成差异性较大,群落存在高度异质性。

图5 不同生境类型步甲和蜘蛛非度量多维度分析 (NMDS) 分析Fig.5 Non-metric Multidimensional Analysis (NMDS) of carbid beetle and spider in different habitat combinations
2.3 非耕作生境对耕地边缘步甲和蜘蛛的影响
就步甲而言(图6A),在分析取样时期耕地边缘与耕地内部和非耕作生境的差异时,步甲多样性均为耕地边缘显著高于耕地内部,虽然耕地边缘均高于非耕作生境,但均未达到差异显著水平。就蜘蛛而言(图6B),蜘蛛多样性耕地边缘与耕地内部和非耕作生境也无显著差异。比较4 种耕地边缘,步甲的多样性在邻近乔木林地的耕地边缘和邻近草地的耕地边缘较高;蜘蛛的多样性在邻近其他林地的耕地边缘较高(图6C)。
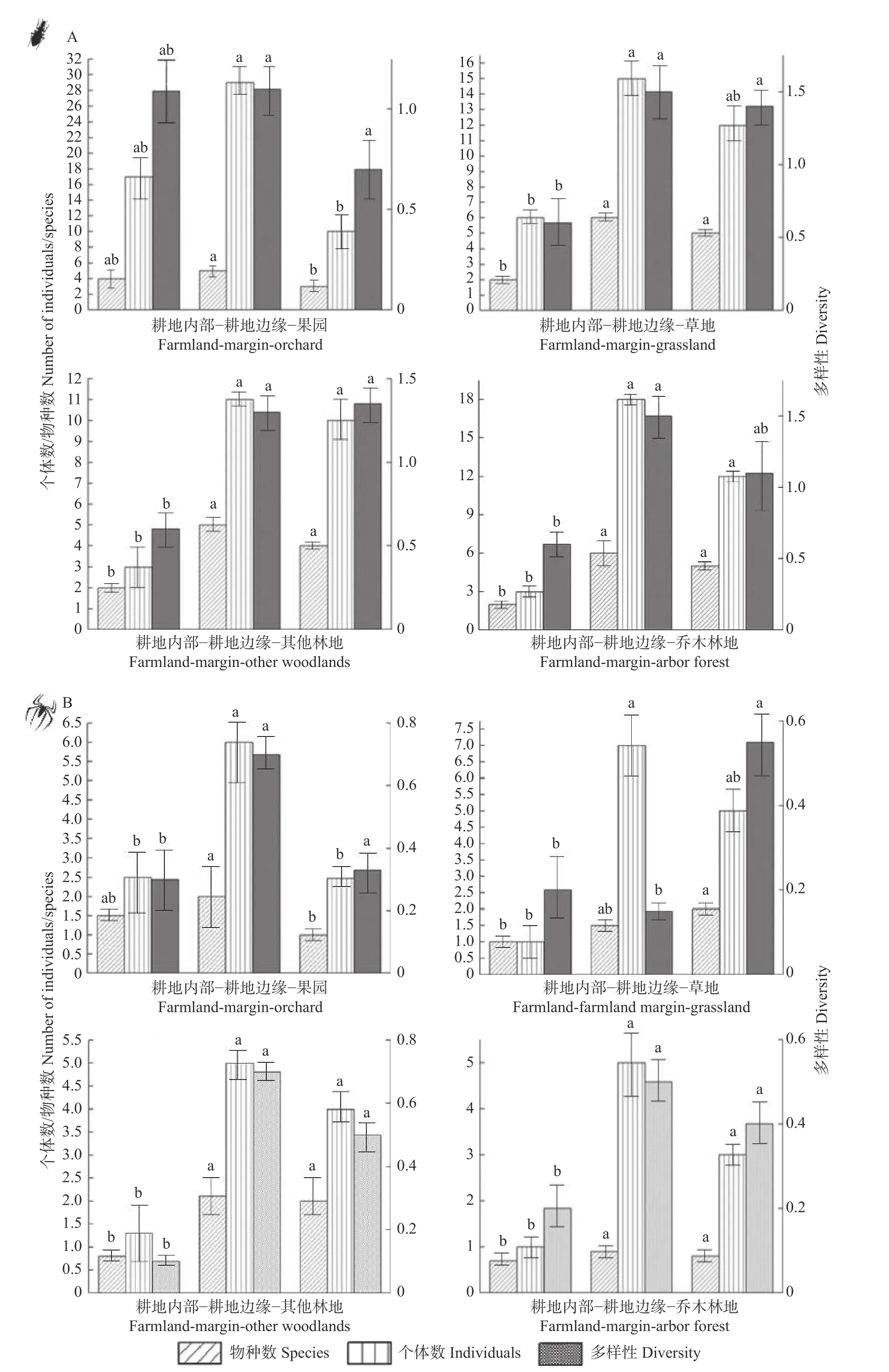
图6 边缘效应对步甲(A)和蜘蛛(B)多样性的影响及不同类型耕地边缘步甲和蜘蛛多样性对比(C)Fig.6 Influence of edge effect on diversity of carabid beetles (A) and spider (B) and comparison of diversity of carabid beetles and spider on edge of different cultivated land (C)
2.4 草本植被结构对步甲和蜘蛛多样性的影响
为进一步探究步甲和蜘蛛多样性与草本植被因子之间的关系,本研究对步甲和蜘蛛的优势种、常见种与草本植被结构因子进行RDA 排序(图7),RDA 前4 排序轴特征值之和为0.591,累积解释了物种−环境关系的99.3%,表明排序结果有较好的解释率。由图7 可知,植被盖度(AC)和植被Shannon 多样性(H)对步甲和蜘蛛群落分布的影响达显著水平(植被盖度F=3.3,P=0.026;植被多样性指数F=3.8,P=0.024),二者共解释了34.7%的步甲和蜘蛛群落数量。蠋步甲(SP1)和球蛛(SP11)与植被多样性指数和植被盖度之间夹角均较小,说明其与植被盖度和植被多样性指数呈正相关关系;广屁步甲(SP7)与植被盖度和植被多样性指数之间夹角较大,呈负相关关系。黄斑青步甲(SP5)和漏斗蛛(SP12)受植被盖度影响更显著(F=4.819,P=0.011;F=6.206,P=0.006),大暗黑步甲(SP2)、普通暗黑步甲(SP4)和淡鞘婪步甲(SP10)受植被多样性影响更显著(F=7.074,P=0.016;F=4.819,P=0.030;F=3.439,P=0.023)。

图7 步甲和蜘蛛优势物种与关键植物结构因子关系冗余分析(RDA)二维排序图Fig.7 Redundancy analysis (RDA) of the relationship between dominant and common carabid beetle and spider and key plant structural factors
3 讨论
3.1 不同类型非耕作生境对步甲和蜘蛛群落结构的影响
非耕作生境对物种多样性具有十分重要的影响,非耕作生境具有丰富的食物资源和植被多样性;可为邻近耕地内的步甲和蜘蛛群落提供稳定的生存环境和长期资源供应[17]。目前国内外关于天敌多样性的研究大多集中于大尺度研究[18],而在中小尺度中,需要通过不同的生境类型来区分和讨论[19]。国内外学者就不同生境对步甲和蜘蛛的影响进行了大量研究,主要集中在自然、半自然和人工3 种生境类型,并讨论了不同生境类型对步甲和蜘蛛多样性的影响[20]。
大量研究表明,由于不同的土地利用方式,非耕作生境类型的差异也会导致步甲和蜘蛛群落结构有不同的变化[21],这在我们的研究中也得到了进一步的补充。例如,林地生境通常具有较大的生境面积以及复杂的植被群落结构[22],可以为步甲和蜘蛛提供更多的栖息地、避难所和食源[23],因此林地生境生物多样性会优于其余人为干扰较大的生境[24],本研究结果也证明乔木林地生境对蜘蛛个体数和物种数影响显著,其他林地生境对蜘蛛多样性影响显著。根据中度干扰假说理论,在中等干扰的情况下,物种种类较多;在轻度干扰的情况下,物种个体数量较多[25]。而本研究的采样区昌图县处于三北防护林体系内,乔木林地多为沿耕地边缘建造的防护林带,人为活动高于规模较大的其他林地。因此,相较于其他林地,乔木林地内拥有更高的物种数和个体数。草地生境通常也能够维持较高的天敌多样性[26],是农业景观中进行害虫生物防治的重要景观因子,步甲可以在草地和相邻耕地两种生境间自由扩散发挥生物防治功能[21],这也与本研究结果相符合。
同时,非耕作生境地产生的溢出效应会影响周围耕地内部步甲和蜘蛛的多样性。当相邻耕地害虫爆发成灾时,这些天敌种群能够迅速通过边缘界面扩散到耕地中发挥生物控害功能[27]。由本研究结论可知,步甲和蜘蛛在草地、其他林地和乔木林地内多样性较高,并且会通过耕地边缘不断向耕地内部扩散,使耕地内部步甲和蜘蛛多样性也有相应的提升。另外,在本研究中,果园生境内步甲和蜘蛛多样性低于其他3 种生境类型,但其相邻耕地内部步甲和蜘蛛多样性却高于其他非耕作生境相邻耕地,该结论与溢出效应作用机制不相符。该结果出现的可能原因是,本研究采样季节为8 月末到9 月初,正值昌图县研究区果园收获季节,而耕地内作物收割还未开始,果园人为扰动大于相邻耕地,步甲和蜘蛛由果园生境迁移到耕地生境,可能导致了相邻耕地内部步甲和蜘蛛多样性增加[28]。也可能是果园生境处边缘效应对步甲和蜘蛛的影响大于溢出效应,这需要进一步讨论得出结论。
生物多样性在人类农业发展中越来越起到至关重要的作用,在2023 年中央一号文件中,国家更加重视耕地的生物特性、农业发展与生物多样性的关系,并且将生物特性与地力水平、耕作条件和健康状况确定为评价耕地质量的4 个一级指标。天敌生物多样性和耕地生态系统的稳定密切相关,良好的生态环境是生物多样性存在的基础,而生物的多样性又改善生态环境。生物种类的数量越多,耕地生态系统就越稳定。反之,生物种类数量越少,耕地生态系统就越脆弱。地表节肢动物多样性对提升耕地质量,建设高标准农田,保障国家粮食安全和重要农产品有效供给具有重要意义。
3.2 不同类型非耕作生境对步甲和蜘蛛边缘效应的影响
作为不同类型生境的空间连接方式,边缘在生态过程中具有重要意义[29]。与张旭珠等[30]研究结果相同,本研究中步甲和蜘蛛的多度、丰富度和多样性均表现为耕地边缘处较高,且与耕地内部差异显著。耕地内部的天敌群落通常以害虫为食物来源,耕地边缘处天敌则以杂草种子为食源,食物的差异造成了耕地边缘和耕地内部的群落结构的差异。因此,即使耕地边缘可以维持大量的步甲和蜘蛛多样性,也可能由于边缘效应,导致边缘处的物种不能向耕地内部流动[3]。
赵紫华等[31]认为,农田与不同类型植被边界带相邻的生境界面上,边界效应所导致的物种流动情况具有显著不同。本研究发现步甲在邻近草地的耕地边缘多样性更高,蜘蛛在邻近其他林地的耕地边缘多样性更高。并且结果表明,耕地边缘步甲和蜘蛛的多样性与非耕作生境无显著差异,这与溢出效应作用结果一致。但早期的研究指出,步甲和蜘蛛通常将耕地边缘作为越冬场所,随后便会向耕地内部扩散,而本研究中耕地内部步甲和蜘蛛多样性却与耕地边缘和非耕作生境存在显著差异。这可能因为在耕地内部存在着特有种[32],在未来研究中,需要进一步探究耕地内部与耕地边缘存在的种类差异,更深入地探讨步甲和蜘蛛由边缘向内部的扩散机制。此外,由于功能群内部存在较为强烈的种间竞争,而步甲在种间竞争中取得优势[33],因此在本研究中邻近草地的耕地边缘处,蜘蛛的多样性低于相邻耕地和非耕作生境。
3.3 非耕作生境内植被结构对步甲和蜘蛛的影响
研究认为,非耕作生境植被群落组成结构是维持天敌多样性的关键,只有特定的植被功能群才能维持大量的天敌多样性[34]。本研究分析显示,草本层盖度、多样性对步甲和蜘蛛群落分布有显著影响。植被盖度的增加会降低鸟类等捕食者的捕食强度,这为步甲和蜘蛛提供了躲避天敌的小生境;异质草本植物结构的组成通常可以为各种分类群提供多样化的微生境,从而为更多生物的生存提供更广的生态位[35]。且不同生物类群对周围环境的响应不同[36],如球蛛科与植被盖度和多样性呈正相关关系,由于草本植被盖度大,有利于其结网、隐蔽及捕食[37];而广屁步甲与其呈负相关关系,相较于非耕作生境,耕地内部害虫密度为捕食性步甲提供了更丰富的食物来源。故在非耕作生境及其与耕地边缘中播种盖度适宜及多样性高的草本植物,可显著提升非耕作生境及其耕地边缘的质量,对增强生物维持、害虫生物防治功能以及提升农业生态系统服务有益。
4 结论
在农业集约化和生物多样性危机的背景下,本文从天敌类地表节肢动物(步甲和蜘蛛)多样性保护的角度,基于4 种常见耕地边缘类型生物多样性数据,讨论了非耕作生境对邻近耕地边缘及内部步甲和蜘蛛群落分布的影响。结论如下:1)不同非耕作生境类型对耕地内部溢出效应明显,但产生的影响有所差异,草地对耕地内部步甲多样性影响显著,其他林地对耕地内部蜘蛛的多样性影响显著。2)不同非耕作生境类型产生的边缘效应与溢出效应规律一致,均会促进耕地内部步甲和蜘蛛的多样性;步甲的多样性在邻近草地的耕地边缘显著高于其他边缘类型,蜘蛛的多样性在邻近其他林地的耕地边缘显著高于其他边缘类型。3)非耕作生境的植被盖度和多样性显著影响步甲和蜘蛛的多样性。本文验证了耕地边缘的非耕作生境可以提升耕地边缘及内部生物多样性。在未来的耕地生态系统保护过程中,可以通过合理布设丰富的非耕作生境,为步甲和蜘蛛提供适宜的生存空间,使其更好地发挥天敌的生态功能。
猜你喜欢
落叶果树(2021年6期)2021-02-12
小朋友·快乐手工(2018年3期)2018-04-22
小学阅读指南·低年级版(2017年6期)2017-06-12
水土保持研究(2016年4期)2016-10-21
新农业(2016年15期)2016-08-16
体育科技(2016年2期)2016-02-28
土壤与作物(2015年3期)2015-12-08
小朋友·快乐手工(2015年1期)2015-03-13
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
应用海洋学学报(2014年2期)2014-11-26
