
5个乌骨鸡品种线粒体 ND6基因遗传多样性及系统进化分析
2023-07-20 09:20:24贾晓旭唐修君陆俊贤樊艳凤马尹鹏黄胜海葛庆联高玉时
西北农业学报 2023年8期
贾晓旭,唐修君,陆俊贤,樊艳凤,顾 荣,马尹鹏,黄胜海,葛庆联,高玉时
(江苏省家禽科学研究所,江苏扬州 225125)
种质资源是农业高质量发展的“芯片”,是解决中国畜牧业种源“卡脖子”问题的物质基础。中国是世界上鸡遗传资源最丰富的国家之一,经过国家畜禽遗传资源委员会鉴定的地方鸡品种已经有116个。经过中国劳动人民长期的人工选择和驯养,形成了以乌骨鸡为代表的独特的变异性状和特有的种质资源。400多年前明代医药家李时珍所著《本草纲目》一书中,就有乌骨鸡的记载:“乌骨鸡,有白毛乌骨者,黑毛乌骨者、斑毛乌骨者,有骨肉皆乌者、肉白乌骨者,但观鸡舌黑者,则骨肉俱乌、入药更良”[1],对这些具有优势特色性状的种质资源进行评价是一项十分重要的工作。
目前对鸡种质资源进行评价的分子标记很多,应用较多的为全基因组水平的遗传标记[2]、微卫星标记[3]和线粒体DNA标记[4]。鸡线粒体基因组全长约16 kp,具有进化速率快、基因结构简单、缺少基因重组和母系遗传等特点,被广泛用于研究种群的起源进化、遗传多样性和品种鉴定等方面[5-8]。目前对鸡线粒体DNA的研究多集中于线粒体控制区(D-loop区)[9]、C氧化酶亚基I(cytochrome C oxidase subunit,COI)[10]、细胞色素b基因(cytochrome b,Cytb)[11]等,利用NADH脱氢酶基因研究鸡遗传多样性和系统进化关系相对较少。在NADH脱氢酶中,NADH 脱氢酶亚基6(NADH dehydrogenase subunit 6,ND6)基因具有序列片段短、进化速率中等、不含影响测序的复杂结构、含有高分辨率的系统发育信息等优点,适合研究物种的遗传多样性和系统进化关系。目前,利用ND6基因对中国乌骨鸡的遗传多样性和系统进化的研究尚未见报道。本研究以中国乌骨鸡地方品种(丝羽乌骨鸡,东乡绿壳蛋鸡,金湖乌凤鸡,余干乌骨鸡)和商业化乌骨鸡品种(新兴竹丝鸡3号配套系,亲本为丝羽骨鸡和隐性白羽鸡,以下简称竹丝鸡)为材料,以线粒体ND6基因全序列为分子标记,评估其线粒体DNA的遗传多样性和系统发育关系,以期为中国乌骨鸡品种的保护、选育及资源鉴定提供理论 依据。
1 材料与方法
1.1 试验材料
丝羽乌骨鸡群体1(n=30)、东乡绿壳蛋鸡(n=30)和金湖乌凤鸡(n=30)样品来自国家级地方鸡种基因库(江苏),丝羽乌骨鸡群体2(n=42)样品来自江西省泰和县泰和鸡原种场、余干乌骨鸡样品(n=36)来自余干县畜禽良种场,竹丝鸡样品(n=30)来自太仓温氏畜牧有限公司。所有采样品种内个体间没有直接的亲缘关系。用血液DNA提取试剂盒[天根生化科技(北京)有限公司]提取基因组DNA,-80 ℃保存,备用。
1.2 试验方法
PCR扩增和测序所用引物相同,正向引物:5′-CCCACAATCGGAACACTA-3′;反向:5′-TCTAACCAAGCGGGAATA-3′,由生工生物工程(上海)股份有限公司合成。PCR反应体系总体积为50 μL,模板DNA(100~200 ng)2 μL, 2×PCR Master Mix(南京诺唯赞生物科技股份有限公司)25 μL,10 μmol/L上下游引物各 1 μL,灭菌水21 μL。PCR反应条件:95 ℃预变性5 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s,运行35个循环;再72 ℃延伸7 min;最后4 ℃终止反应。PCR产物经20 g/L琼脂糖凝胶电泳检测,确认扩增片段与目的片段一致后,送生工生物工程(上海)股份有限公司测序。
1.3 数据处理与分析
用DnaSP 5.10.1软件统计核苷酸多样性、单倍型多样性、平均核苷酸差异、变异位点数、单一突变位点数、简约信息位点数和单倍型数[12]。用MEGA 7.0软件统计序列的平均碱基组成、品种内和品种间的遗传距离,采用邻接法(neighbor-joining,NJ)构建系统发育树[13]。采用Network 10.2.0软件绘制Median-joining单倍型网络图[14]。
2 结果与分析
2.1 线粒体 ND6基因序列变异分析
PCR扩增片段长度为732 bp,减掉两端非ND6基因序列区域片段,成功获得197条完整的线粒体ND6基因序列,长度全部为522 bp,没有发现插入或缺失。碱基T、C、A和G的平均比例分别是10.27%、38.77%、41.50%和9.46%, A+T的平均含量为51.77%,略高于G+C的含量48.23%,表现出一定的AT碱基偏好性,与鸟类线粒体DNA碱基组成的特征相符。
利用DnaSP 5.10.1软件对197条序列进行单核苷酸多态性分析,共检测到8处多态位点(表1),占核苷酸序列总长度的1.53%(8/522),其中2个单一突变位点位于204、467位,6个简约信息位点位于64、70、178、253、403和418位。所检测到的8处多态位点均为转换,其中A/G之间转换4 次,T/C之间转换4 次。
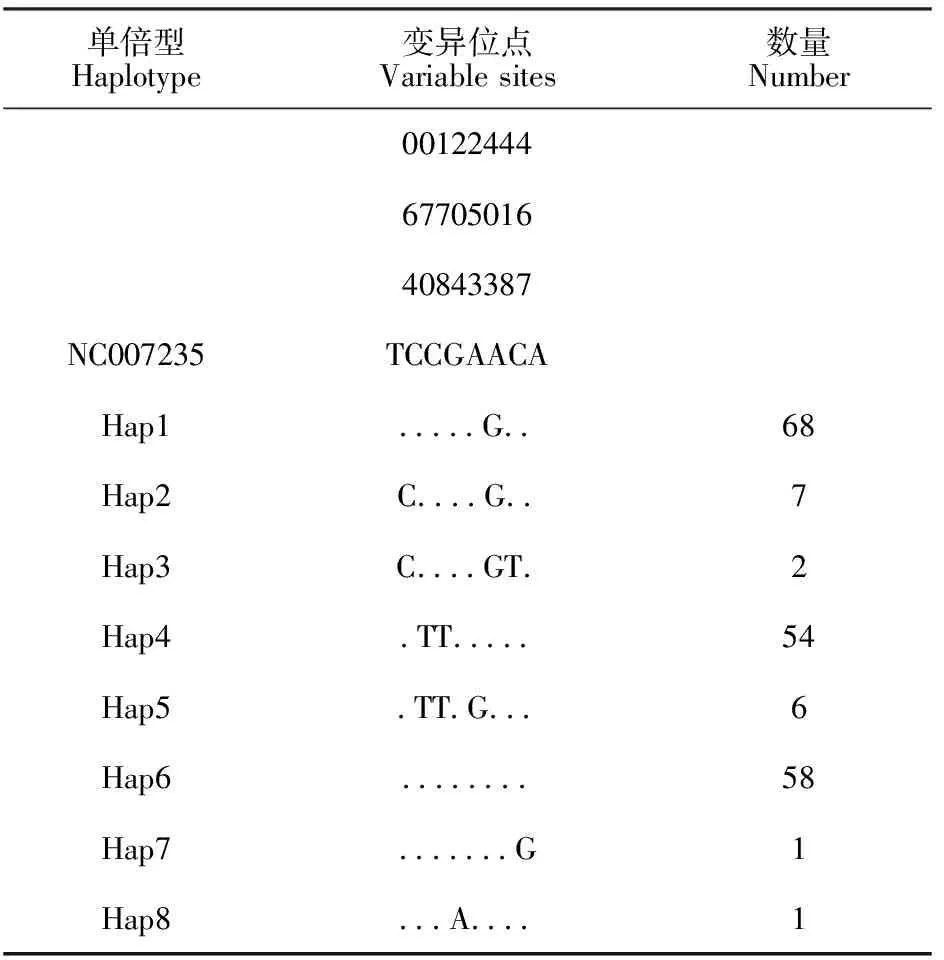
表1 基于 ND6基因序列变异信息的乌骨鸡单倍型分类
2.2 线粒体 ND6基因单倍型多样性分析
基于197条乌骨鸡序列发现的8个多态位点,共界定8种单倍型,命名为Hap1~Hap8。各单倍型的数量及在品种间的分布见表1和表2。其中单倍型Hap1出现频率最高,共68次;其次是单倍型Hap6,为58次;然后是单倍型Hap4为54次。Hap1、Hap2、Hap4和Hap6为共享单倍型,其中Hap4和Hap6在所有群体都有检测到;Hap1除丝羽乌骨鸡2个群体,其他品种均能检测到;Hap2除丝羽乌骨鸡群体1和竹丝鸡,其他群体均能检测到。Hap3、Hap5、Hap7和Hap8为独立单倍型,Hap3只分布于余干乌骨鸡,Hap5只分布于东乡绿壳蛋鸡,Hap7和Hap8分别只分布于丝羽乌骨鸡群体1和余干乌骨鸡。

表2 6个乌骨鸡群体单倍型分布
2.3 不同品种遗传多样性和距离分析
6个乌骨鸡群体遗传多样性见表3。总体单倍型多样性为0.720±0.012,最高的为东乡绿壳蛋鸡为0.766±0.042,最低的竹丝鸡为0.191±0.093。总体核苷酸多样性为0.002 91± 0.000 92,东乡绿壳蛋鸡核苷酸多样性最高为 0.003 83±0.001 08,竹丝鸡最低为0.000 85±0.000 84。平均核苷酸差异为1.518,余干乌骨鸡平均核苷酸差异最高为1.998,竹丝鸡最低为 0.444。

表3 6个乌骨鸡群体的遗传多样性
基于Kimura双参数模型计算6个群体乌骨鸡的遗传距离见表4,可以看出6个群体品种内遗传距离为0.000 86~0.003 87,竹丝鸡最低,东乡绿壳蛋鸡最高。品种间遗传距离为0.000 10~0.003 29。竹丝鸡与2个丝羽乌骨鸡群体之间的遗传距离较远,双参数距离分别为0.003 29和 0.001 57;余干乌骨鸡与金湖乌凤鸡的遗传距离最近,双参数距离值为0.000 10。

表4 基于Kimura双参数模型计算的6个乌骨鸡群体品种内和品种间平均遗传距离
2.4 系统进化关系分析
用Network 10.2.0构建6 个乌骨鸡群体的中介网络图,如图1所示,6 个乌骨鸡群体以Hap6为中心呈星状发散分布,没有按鸡体型外貌或地理来源的不同进行分类,只是在单倍型分布和比例存在差异。丝羽乌骨鸡群体1包含3种单倍型,Hap4单倍型在该鸡群体中占优势;丝羽乌骨鸡群体2有3种单倍型,Hap6单倍型在该群体中占优势;东乡绿壳蛋鸡、金湖乌凤鸡、余干乌骨鸡和竹丝鸡分别有5、4、6和3 种单倍型,优势单倍型都为Hap1。采用NJ法构建6 个群体的系统发育树(图2),发育树分成2 个大枝,丝羽乌骨鸡群体1和丝羽乌骨鸡群体2聚为一支,东乡绿壳蛋鸡、金湖乌凤鸡、余干乌骨鸡和竹丝鸡聚成一支。

不同颜色表示不同群体

图2 基于mtDNA ND6基因采用NJ法构建的系统发育树
3 讨 论
5个品种乌骨鸡线粒体ND6基因序列的碱基T、C、A和G的平均比例分别是10.27%、38.77%、41.50%和9.46%,并且G含量最低,显示出较明显的碱基偏倚,这是由于蛋白质编码基因的核苷酸突变在密码子第3位点上受到的自然选择压力较小所致,证明密码子第3位点能够更清晰地表明线粒体基因组核苷酸组成的不均一性[15]。序列变异一般认为是生物适应环境变化的结果,是产生生物多样性的根本来源,本研究发现乌骨鸡mtDNAND6基因序列长度为522 bp,发现的8处多态位点为64~467 bp,具有较高的突变率,可作为鸡遗传多样性和起源进化研究的分子标记。
核苷酸多样性和单倍型多样性是衡量群体遗传多样性高低的两个重要指标,数值越大,具有较大的适应能力、生长能力和进化潜力,更容易开拓新环境[16]。本研究结果发现除5个乌骨鸡品种除东乡绿壳蛋鸡和余干乌骨鸡具有很高的遗传多样性以外,其他品种均处于较低水平。丝羽乌骨鸡群体2(样本来自原产地群体)的遗传多样性大于丝羽乌骨鸡群体1(样本来自基因库保种群体),并且在单倍型分布比例上存在差异,可能与保种地和原产地环境差异有关,提示保种过程中要定期与原产地群体进行基因交流[17]。竹丝鸡作为商业化品种,单倍型多样性和核苷酸多样性分别只有0.191和0.000 85,品种内遗传距离为0.00 86,均低于其他乌骨鸡地方品种。这种现象说明品种培育过程中,为了提高生产性能进行高强度人工选择会使群体的遗传多样性大大降低。
丝羽乌骨鸡已经有数百年的繁衍历史,曾作为珍贵贡品上供朝庭,是中国最古老的鸡品种之一。丝羽乌骨鸡外貌与其他鸡种有明显的不同,标准的丝羽乌骨鸡具有“十全”特征:桑葚冠、缨头、胡须、绿耳、丝羽、毛脚、五爪、乌皮、乌肉和乌骨。同时丝羽乌骨鸡具有性情温驯、适应性强、外形美观、肉质鲜嫩、药用功能,是药、肉、蛋、观赏兼用型多用途鸡种[1]。同中国其他地方鸡品种一样,丝羽乌骨鸡生产性能不高,通过本品种选育很难有大幅度提升。竹丝鸡是在丝羽乌骨鸡基础上培育而成。竹丝鸡和丝羽乌骨鸡虽然体型外貌相似,但生长速度和繁殖性能都得到大幅度提升,最终产品的价格也要便宜很多。本研究表明竹丝鸡与两个丝羽乌骨鸡群体之间的母系遗传距离较远,双参数距离值分别为0.003 29和0.001 57,与竹丝鸡父本来自丝羽乌骨鸡,母本来自隐性白羽肉鸡培育过程相符。同时可基于二者单倍型的不同,进行丝羽乌骨鸡和竹丝鸡产品甄别,保证优质优价。
线粒体作为细胞器位于细胞核外,线粒体DNA的遗传信息是通过卵细胞传递给下一代,同时线粒体DNA变异具有累积效应,来自同一母系的单倍型会聚类在一起,称为线粒体单倍型类群。本研究中6个乌骨鸡群体以Hap6为中心呈星状发散分布,没有像D-loop区序列一样形成多个单倍型类群,可能与ND6基因进化速率低于 D-loop区有关,但过高的进化速率会产生重复突变,从而干扰系统发育树的构建,特别是在种间及以上的系统发育分析中不能准确地反映各个支系的关系[5]。鸡的线粒体基因组长度一般为 16 783~16 787 bp,除了线粒体D-loop区,还包含13个编码蛋白基因、22个tRNA基因、2个rRNA基因(12S rRNA和16S rRNA),包含更多的遗传信息[18],因此本研究认为利用线粒体基因组或几个不同区域的序列组合能够更为全面准确地掌握鸡的母系起源情况。
4 结 论
乌骨鸡线粒体ND6基因多态信息含量丰富,可作为遗传多样性和起源进化的研究分子标记。东乡绿壳蛋鸡和余干乌骨鸡具有丰富的遗传多样性,丝羽乌骨鸡和金湖乌凤鸡的遗传多样性相对较低,商业化品种竹丝鸡的遗传多样性处于极低水平。可以利用竹丝鸡与丝羽乌骨鸡母系来源的不同,开发线粒体分子标记区分竹丝鸡和丝羽乌骨鸡。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
中国畜禽种业(2020年7期)2020-12-16 14:47:06
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
卷宗(2018年7期)2018-06-06 05:27:16
艺术与设计·理论(2018年3期)2018-02-02 03:26:40
三联生活周刊(2016年35期)2016-08-29 17:37:58
中国畜牧兽医文摘(2016年7期)2016-01-30 23:38:24
癌变·畸变·突变(2014年2期)2014-03-01 04:39:42
祝您健康(2012年1期)2012-06-04 09:31:15
