
禽致病性大肠杆菌体外感染鸡巨噬细胞模型的建立
2023-07-08 12:34:42石海涛谭跃荣申思阳彭璐媛伊鹏霏付本懂
中国兽医学报 2023年5期
石海涛,谭跃荣,申思阳,彭璐媛,伊鹏霏,付本懂
(吉林大学 动物医学学院,吉林 长春 130062)
鸡大肠杆菌病是由禽致病性大肠杆菌(avian pathogenicEscherichiacoli,APEC)引起的一种禽类细菌性疾病,作为家禽养殖业中常见的群发病,饲养环境、应激反应或者其他疾病均可成为此病发生的诱因[1]。
巨噬细胞作为肺脏中常驻免疫细胞,成为大肠杆菌呼吸道感染的第一道免疫防线,对鸡抵抗大肠杆菌感染具有重要作用[2]。
目前鸡大肠杆菌病模型,大多用大肠杆菌直接感染鸡体,例如腹腔注射,灌胃,胸肌注射等[3],但尚未建立成熟稳定的大肠杆菌体外感染巨噬细胞HD11的方法,并且HD11细胞的培养条件大多维持在37℃,并未模拟鸡的体温在41℃条件下培养[4]。因此,本研究通过采用APEC-O78感染HD11细胞,通过比较不同感染复数,不同感染时间,不同温度下APEC感染HD11细胞后,炎症基因表达的变化,筛选出最佳感染条件,建立稳定的APEC体外感染模型。
1 材料与方法
1.1 菌液制备APEC-O78菌株来自中国兽医微生物菌种保藏管理中心,将菌种干粉进行活化,鉴定后划线培养,取单菌落于LB肉汤培养基中进行培养,测定生长曲线,并取对数期细菌待用。
1.2 细胞培养与传代HD11细胞由扬州大学朱国强教授惠赠,使用RPMI-1640培养基、8%胎牛血清,2%鸡血清配制完全培养基,待HD11细胞长满T25细胞培养瓶后,弃掉培养基,加入1 mL预热的PBS清洗细胞后,加入300 μL胰酶(Gibco)消化15 s 弃掉,加入2 mL完全培养基,使用1 mL移液枪吹打细胞至脱落后,转移至2 mL离心管,使用离心机1 000 r/min离心5 min,弃上清,使用完全培养基混悬后,分至2个T25细胞培养瓶,放入37℃、5% CO2细胞培养箱中培养。
1.3 2种温度对APEC增殖的影响以5 g∶200 mL 的比例,配制LB肉汤培养基(青岛海博生物技术有限公司)于2个锥形瓶,各100 mL,各加入100 μL对数期APEC菌液,分别培养于37℃与41℃摇床中,间隔2 h以Q6000测定其D600 nm,绘制生长曲线。
1.4 APEC感染对HD11细胞损伤的检测将细胞以1×105/孔接种至48孔板,于37℃培养箱中培养12 h后,弃培养基,以MOI=50、MOI=51、MOI=52、MOI=53的感染复数,将APEC接种至48孔板,分别在0.5,1,2,4,6 h吸取培养基上清,离心去除沉淀后,使用碧云天LDH检测试剂盒按照说明书检测细胞上清中LDH含量。
1.5 荧光定量PCR法检测不同感染复数大肠杆菌在连续时间内对HD11细胞炎性基因表达的影响将细胞以1×106/孔接种至6孔板,于37℃培养箱中培养12 h后,弃培养基,以MOI=50、MOI=51、MOI=52、MOI=53的感染复数,将APEC接种至孔板内,分别在0.5,1,2,4 h时收集细胞。
1.6 荧光定量PCR法检测不同感染复数APEC在2种温度下对HD11细胞炎性基因表达的影响将细胞以1×106/孔接种至6孔板,培养12 h后,弃培养基,以MOI=53、MOI=52、MOI=51、MOI=50、MOI=5-1、MOI=5-2、MOI=5-3的感染复数,将APEC接种至6孔板,分别置于37,41℃含5% CO2恒温培养箱培养,4 h后收集细胞。使用TRIzol法提取细胞总RNA,使用Q6000测定RNA的质量和质量浓度,以每个样本1 μg的体系使用反转录试剂盒按照说明书进行反转录,采用20 μL的荧光定量PCR体系:cDNA模板1 μL,上、下游引物各0.5 μL,Roche FastStartTM通用SYBR®Green预混液(ROX)10 μL,并用无酶水补足至20 μL,采用3步法进行信号采集:95℃预变性10 min;95℃变性15 s,55℃退火20 s,60℃延伸30 s,40个循环。引物序列如表1所示。

表1 实时荧光定量PCR引物序列
1.7 Griess法检测细胞上清中NO水平将细胞以5×105/孔接种至24孔板,培养12 h后,弃培养基,以MOI=50、MOI=51、MOI=52、MOI=53的感染复数,将PAEC接种至24孔板,分别在41℃培养0.5,1,2,4 h后收集细胞上清,按照南京建成NO检测试剂盒说明书对细胞上清进行处理,并用酶标仪测定各孔D550 nm值。
1.8 DCFH-DA法检测细胞ROS水平将细胞以2×104/孔接种至24孔板,培养12 h后,弃培养基,使用纯培养基清洗细胞3次,使用无血清培养基将DCFH-DA(Sigma)稀释至10 μmol/L并加入到96孔板内,41℃孵育30 min后再次清洗干净未装载的探针,以MOI=50、MOI=51、MOI=52、MOI=53的感染复数,将APEC接种至孔板内,41℃培养,分别在0.5,1,2,4 h后,使用488 nm激发波长,525 nm发射波长,检测荧光强度。
1.9 统计学分析所有数据经过Excel软件初步处理整合后,使用SPSS 19.0统计软件进行方差分析,使用绘图软件GraphPad Prism 9.0进行绘图。
2 结果与分析
2.1 不同温度下APEC的生长曲线首先为了确定常规培养温度37℃与试验温度41℃对APEC生长是否有影响,分别在2种不同温度下对APEC的生长曲线进行测定。如图1所示,APEC-O78在相同培养时间内,在不同温度下显示出相同的生长活性,并在前5 h内,其生长状态无明显差异。

图1 不同温度下大肠杆菌生长曲线
2.2 APEC感染对HD11细胞LDH的影响为了筛选出APEC对于HD11细胞的最大感染时间以及感染复数,确定对细胞无直接损伤的感染范围,对不同感染复数在不同时间内HD11细胞培养上清的LDH进行检测,以lysis作为阳性对照,结果如图2所示,用不同MOI感染HD11细胞,在0~4 h,除MOI=53组在4 h有显著变化外,其余各组细胞LDH释放均无显著变化;而当感染时间到达6 h,不同MOI感染细胞的LDH释放量均显著高于对照组,因此后续大肠杆菌感染HD11细胞的作用时间均小于4 h。
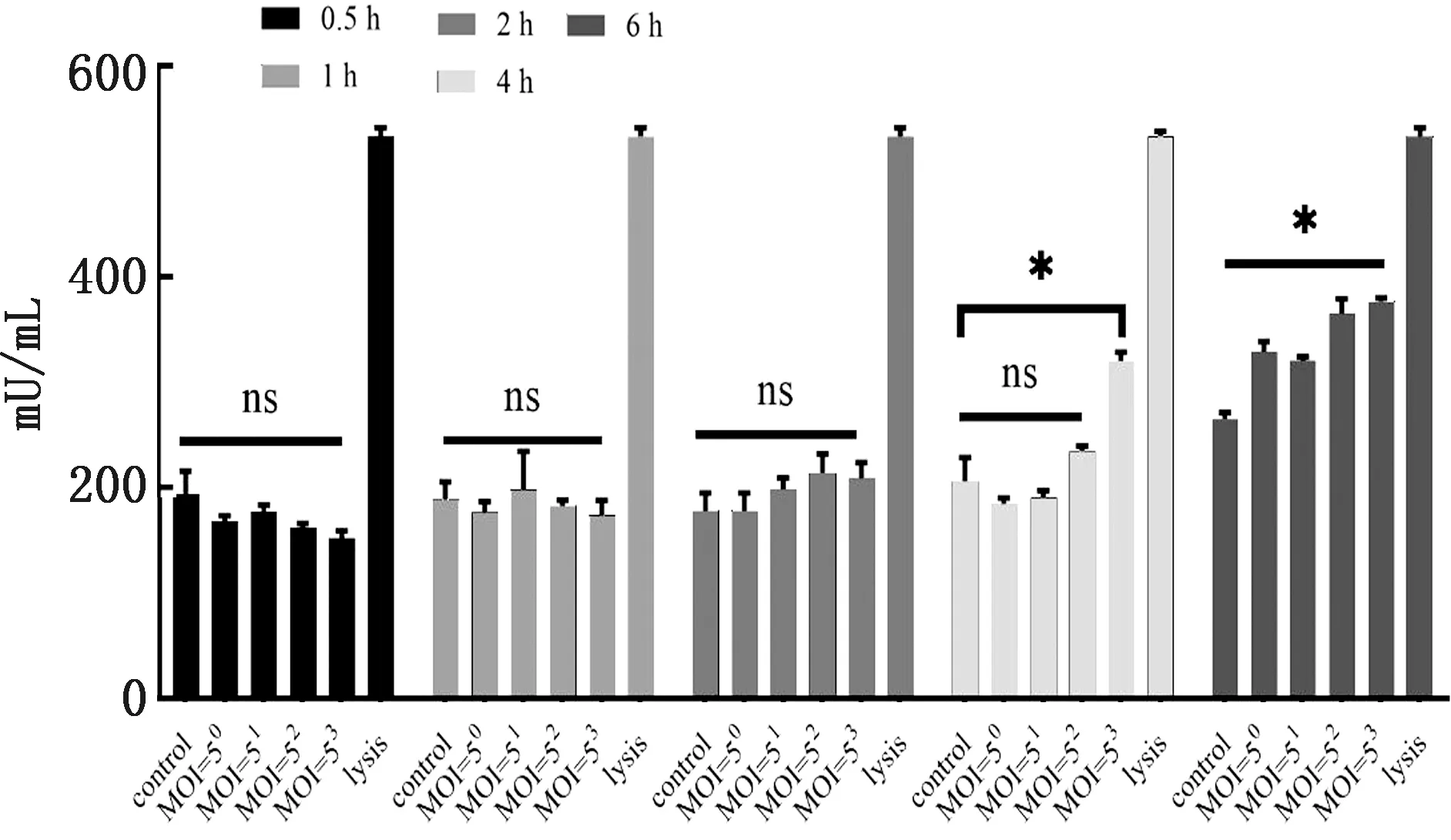
ns.表示P>0.05;*.表示P<0.05;**.表示P<0.01。下同
2.3 不同MOI APEC感染HD11细胞后炎性基因表达变化为了对比不同的APEC感染浓度在不同的感染时间内对于HD11细胞的影响,对APEC感染HD11细胞后相关炎症因子进行检测,结果如图3所示,当APEC感染HD11细胞的作用时间为0.5 h时,炎症基因的表达量与对照组相比并无明显差异。当MOI=53,作用时间为1 h时,仅IL-6相对表达量显著高于对照组(P<0.05)。而当APEC感染HD11细胞的作用时间为2 h时,4种炎症基因的相对表达量显著高于对照组(P<0.05),但IL-6 mRNA相对表达量在MOI=53时与MOI=51、MOI=52时相比有降低趋势。当APEC感染HD11细胞的作用时间为4 h,当MOI=50、MOI=51、MOI=52、MOI=53时,IL-1β、IL-6、TNF-α、iNOS的相对表达量均极显著或显著高于对照组(P<0.01或P<0.05);但当MOI=52、MOI=53时,4种基因的相对表达量相较于MOI=50、MOI=51时均有所降低。因此,在0~4 h内,4种基因的相对表达量呈时间依赖性,且在4 h时,4种炎症基因的相对表达量达到峰值,但与感染复数并无正相关关系。
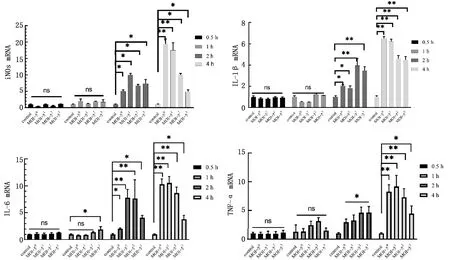
图3 在时间梯度内不同MOI APEC感染对HD11细胞炎症因子相对表达量的影响
2.4 不同温度下APEC感染对HD11细胞、促炎基因表达的影响因4种基因的相对表达量在4 h时达到峰值,因此确定PAEC感染HD11的作用时间为4 h,并扩大感染复数的梯度范围,以确定2种温度下不同感染复数对4种基因表达的影响。如图4所示,当培养温度为41℃时,iNOS的相对表达量在不同MOI下,均显著高于37℃(P<0.05),其中当MOI=50、MOI=51、MOI=52时,差异极显著(P<0.01);IL-1β的相对表达量除MOI=5-3以外,均显著高于37℃(P<0.05),其中MOI=51、MOI=52、MOI=53差异极显著(P<0.01);IL-6的相对表达量在MOI=51、MOI=52、MOI=53时均显著高于对照组(P<0.05),其他组无明显变化;TNF-α的相对表达量在MOI=50、MOI=51、MOI=52、MOI=53时显著高于37℃(P<0.05),其中当MOI=51时,差异极显著(P<0.01)。可见当培养温度为41℃时,HD11细胞对于大肠杆菌的反应活性显著高于37℃。当MOI=51时,HD11细胞对PAEC的反应活性相对于其他感染复数更为强烈。
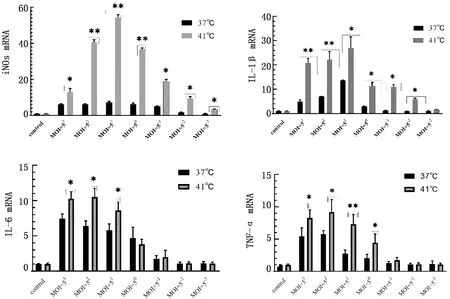
图4 不同温度下大肠杆菌感染HD11细胞对炎性基因表达的影响
2.5 APEC感染对HD11细胞IL-10、ROS以及NO的影响通过前面的试验发现,APEC感染后HD11细胞的活化与感染时间和感染复数并无直接的正相关关系,为查明原因,我们对HD11细胞的IL-10、ROS以及NO的变化进行分析。
2.5.1在梯度时间内不同感染复数APEC感染对HD11细胞IL-10基因表达的影响 如图5所示,IL-10在共培养1 h以内无明显变化,当时间达到2 h后,IL-10的表达显著升高(P<0.05)。
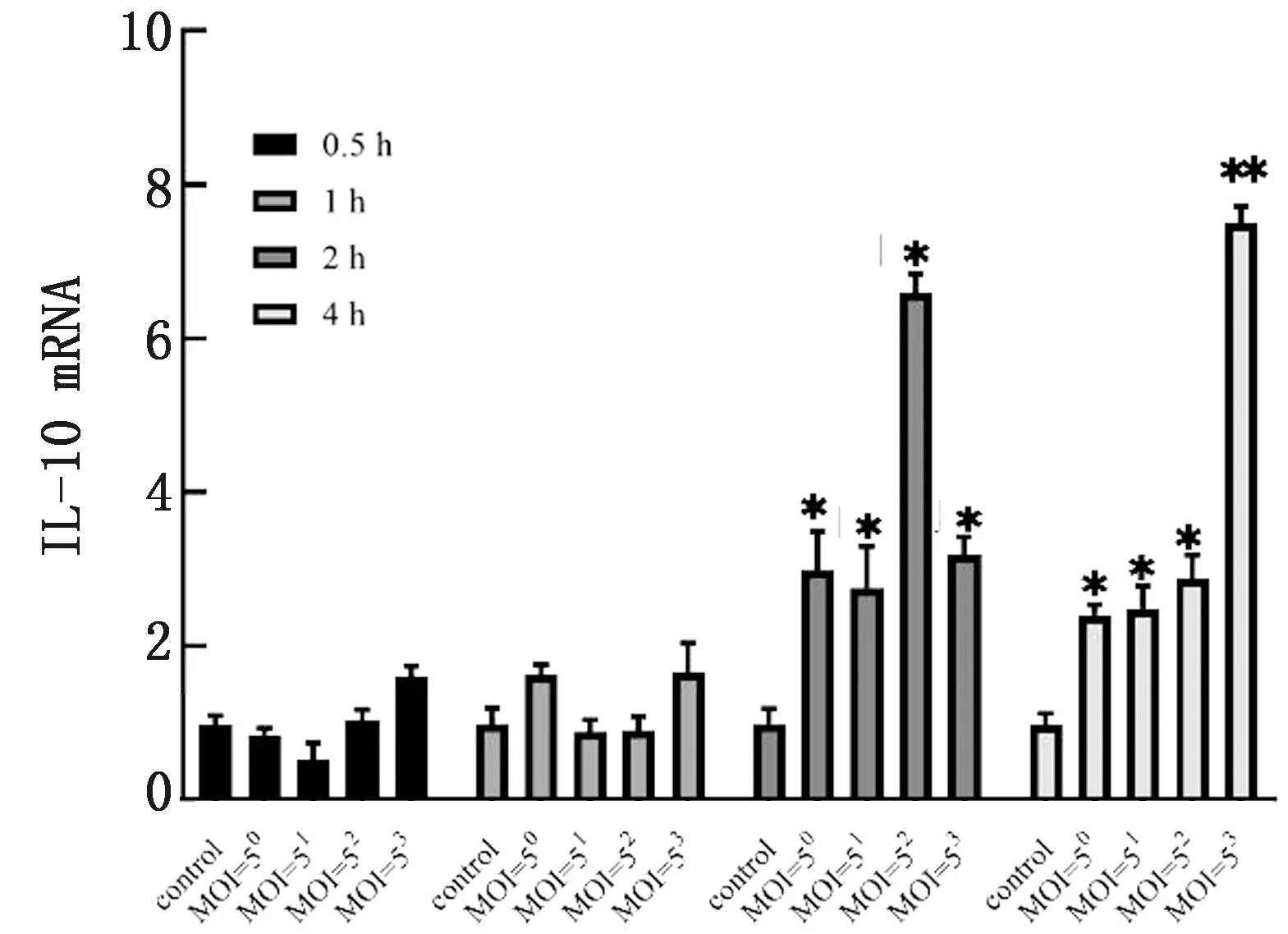
图5 不同MOI APEC感染在0~4 h内对HD11细胞IL-10基因表达的影响
2.5.2在梯度时间内不同感染复数APEC感染对HD11细胞NO分泌的影响 为探究不同的感染时间下,不同浓度APEC感染HD11细胞后对NO的产生的影响,使用NO检测试剂盒对培养上清进行检测,如图6所示,利用不同MOI APEC感染HD11细胞,不同时间内NO含量具有明显的变化。其中孵育时间为0.5 h,MOI=52、MOI=53组内NO含量显著上升(P<0.05),随着时间延长至1 h,MOI=53组NO含量极显著升高(P<0.05),但当时间到达2 h时,虽然MOI=52、MOI=53组NO含量仍高于正常组,但相比于之前的时间,MOI=53组的NO含量有所下降,当时间到达4 h后,MOI=53组的NO含量显著下降(P<0.01),而其他组逐渐趋于正常。

图6 不同MOI APEC对HD11细胞在0~4 h内NO释放的影响
2.5.3在梯度时间内不同感染复数APEC感染对HD11细胞ROS的影响 已知促炎因子会导致巨噬细胞M1型极化,可以引起ROS的产生,因此对APEC感染后HD11细胞的ROS变化进行检测,如图7所示,HD11细胞在受到感染刺激后,0.5 h时ROS水平具有显著升高(P<0.05),且与MOI呈正相关,随着时间延长至4 h,MOI=53组的ROS水平显著高于正常组,但当时间为4 h时,MOI=52、MOI=53组的ROS水平相较于2 h有所降低。
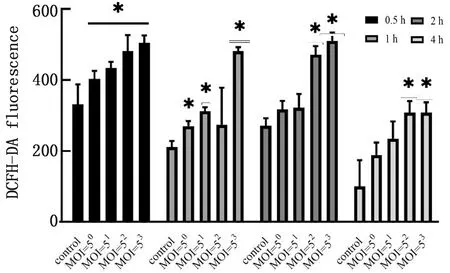
图7 不同MOI APEC感染在0~4 h内对HD11细胞ROS的影响
3 讨论
巨噬细胞作为大肠杆菌突破免疫屏障的第一道防线,建立体外大肠杆菌感染模型是研究大肠杆菌致病机制的另一种途径。本研究在建立模型的过程中发现,大肠杆菌感染巨噬细胞的作用时间并非是越久越好,当时间达到6 h后,大肠杆菌在不同感染复数下,都可对巨噬细胞产生严重的破坏,引起LDH显著升高,在4 h左右,巨噬细胞的反应就可达到最大值,当确定作用时间后,筛选感染复数过程中发现,因大肠杆菌复制速度较快,过高的感染复数非但不会增强巨噬细胞反应性,相反对巨噬细胞炎性反应产生抑制作用,在LDH水平并无明显变化的情况下,过高浓度的大肠杆菌可能会抑制巨噬细胞的某些作用。而不同的培养温度也可能会导致巨噬细胞活性改变。禽巨噬细胞由BEUG等[5]建立之初,就培养于37℃下,后续研究也大多维持于37℃条件下[6],37℃对于细菌和细胞而言都是传统的最佳培养条件,但是考虑到APEC与HD11细胞都来源于鸡,因此对培养温度是否应模拟鸡的生理温度(41.5±0.5)℃而产生疑问,因此在这2种温度下进行了试验。通过比较2种温度下HD11细胞对APEC刺激的基因表达情况,发现41℃下HD11细胞的炎症反应相较于37℃强烈的多。41℃作为鸡的正常生理温度,在应对APEC感染时,HD11细胞能够产生更多的促炎因子,从而可以引起机体免疫系统针对APEC感染产生更好的防御能力,而37℃降低了HD11细胞的反应性,或许这可以解释饲养环境温度低会成为大肠杆菌病发生的诱因之一。但HD11细胞因抵抗APEC感染而导致促炎因子基因水平的提高并非没有限制,相反,随着感染时间的延长以及APEC数量的逐渐增多,HD11的活性也会受到抑制,其中TNF-α、IL-1β、IL-6这3种促炎因子,显示出相同的变化趋势,而 iNOS作为巨噬细胞M1型极化的一种标志性基因[7-8],其拥有促进巨噬细胞产生NO的作用[9],其表达趋势的变化与促炎因子极为相似,即低感染浓度与高感染浓度都会产生较低的基因表达水平。当低感染浓度时, APEC的数量不足以诱导HD11细胞活化,而高感染浓度相关基因表达水平低,可能与APEC具有某些免疫抑制的作用而相关。为了验证这一结果,又对M2极化的一种标志基因—IL-10[10]进行检测分析,结果发现随着感染浓度的升高,IL-10反而有显著升高的变化趋势。当APEC感染浓度导致HD11细胞的3种促炎相关基因以及M1型标志基因iNOS的表达水平显著降低时,IL-10的表达水平急剧升高,也因此可排除HD11细胞可能因感染而死亡导致RNA降解的可能性。类似于结核分枝杆菌会导致M1极化的巨噬细胞向M2表型转换的现象[11],诱导巨噬细胞产生IL-10,有利于细菌在巨噬细胞内的存活和生长[12]。通过对HD11细胞产生ROS的水平进行检测,发现M1型巨噬细胞支持ROS持续产生的功能受到抑制,因此认为对于APEC体外感染HD11细胞的浓度,需要限定在一定范围之内。
综上所述,HD11细胞在41℃条件下,相比于37℃,其对APEC感染具有更高的反应活性;在MOI=51时,其应对感染的产生的反应更剧烈,因此,本研究选择使用MOI=51的APEC-O78作用于HD11细胞,41℃条件下培养4 h,可建立相对反应性较好的APEC体外感染巨噬细胞的模型,并可以此为基础,从细胞水平上探究大肠杆菌与巨噬细胞之间的互作关系,进一步深入研究鸡大肠杆菌的致病机制,为本病的防控提供理论依据。
猜你喜欢
核科学与工程(2022年3期)2022-10-18 01:25:14
中学生数理化(高中版.高考数学)(2021年11期)2021-12-21 05:34:28
中学生数理化(高中版.高二数学)(2021年4期)2021-07-20 07:18:48
中学生数理化(高中版.高二数学)(2021年4期)2021-07-20 07:18:46
广州化工(2020年6期)2020-04-18 03:30:20
新世纪智能(数学备考)(2020年12期)2020-03-29 02:15:34
现代矿业(2018年9期)2018-10-16 09:37:02
中央民族大学学报(自然科学版)(2016年3期)2016-06-27 07:55:28
兽医导刊(2016年12期)2016-05-17 03:51:54
中国医药生物技术(2015年4期)2015-12-26 08:26:36
