
肠道菌群短链脂肪酸与肺结核相关性研究进展
2023-07-01 07:35:38张晓萌李敏柴英辉周静雷红
中国防痨杂志 2023年7期
张晓萌 李敏 柴英辉 周静 雷红,
肺结核是由结核分枝杆菌(Mycobacteriumtuberculosis, MTB)引起的世界范围内最致命的慢性全身性传染病之一,主要经过呼吸道传播,也是单一传染病导致死亡的主要原因。世界卫生组织发布的《2022年全球结核病报告》[1]指出,2021年全球新发结核病患者数高达1060万例,我国估算的患者数为78万例,大约全球人口的1/4存在结核分枝杆菌感染。因此,提出新的抗结核治疗方法,开发新的抗结核药物已经迫在眉睫。
随着研究的深入,众多学者认为肺结核是MTB的免疫反应失衡造成的,且越来越多的研究表明肠道菌群的代谢产物与机体免疫系统之间存在明显的相互作用[2-5]。短链脂肪酸(short-chain fatty acids,SCFA)是肠道菌群的主要代谢产物,由膳食纤维经肠道菌群酵解后生成[5]。研究表明,SCFA与多种呼吸系统疾病具有内在关联性,比如:慢性阻塞性肺疾病、哮喘、肺囊性纤维化和肺结核等[6-8]。肺结核是常见的传染性高的呼吸道疾病,它与SCFA之间的相互作用关系成为了研究的热点[8-9]。因此,笔者系统阐述SCFA与肺结核的相关性,SCFA参与调节肺结核的可能机制,以及通过调节肠道菌群改善SCFA的丰度,治疗或辅助治疗肺结核的可行性。
一、SCFA概述
SCFA是指含有1~6个碳原子的有机羧酸,主要分为甲酸、乙酸、丙酸、丁酸及其支链脂肪酸和一些盐类,是由没有被吸收的食物残渣中的碳水化合物经结肠内厌氧菌产生[10]。SCFA是肠道菌群的主要代谢产物,肠道中包含最多的SCFA是乙酸、丙酸和丁酸,约占SCFA总和的95%以上[11]。其中,乙酸是肠道菌群为机体提供能量的主要来源,提供的能量约占人体日常所需的10%[12]。产生乙酸的主要菌种有:拟杆菌、双歧杆菌、普氏菌、瘤胃球菌、布劳特氏氢营养菌、 梭状芽孢杆菌和链球菌等。丙酸可参与丙酮酸逆转化为葡萄糖的过程,且能够阻碍胆固醇合成并促使其代谢[13]。产生丙酸的主要菌种为:拟杆菌、厚壁菌、考拉杆菌、小类杆菌、韦荣菌、埃氏巨型球菌、粪球菌、沙门菌、罗氏菌和瘤胃球菌等。丁酸是可以提供肠道所需能量的60%~70%,并且可以抑制癌细胞的侵袭和增殖,并诱导其凋亡[14]。产生丁酸的主要菌种为:粪球菌、丁酸弧菌、粪球菌、直肠真杆菌、霍氏真杆菌、柔嫩梭菌和罗氏菌等。3种SCFA在结肠中的摩尔比约为 60∶25∶15,且从近端结肠到远端结肠浓度逐渐降低,近端结肠浓度约为70~140 mmol/L,远端约为20~70 mmol/L。不同种类SCFA的浓度比不但与肠道内SCFA产生菌的组成有关,而且与宿主代谢和食物中的纤维含量有关,合理的补充膳食纤维可以起到调节肠道中SCFA浓度和比例的作用。
SCFA通过参与消化吸收营养、调节免疫系统、保护机体免受病原入侵等生理过程,从而对机体产生有益的作用[15-16]。研究表明,SCFA与多发性硬化症[17]、肥胖[18-19]、糖尿病[20]、心血管疾病[21]、多种癌症[22-23]及肺部疾病[6-8]均具有相关性,不同种类的SCFA与各类疾病相关性见表1。
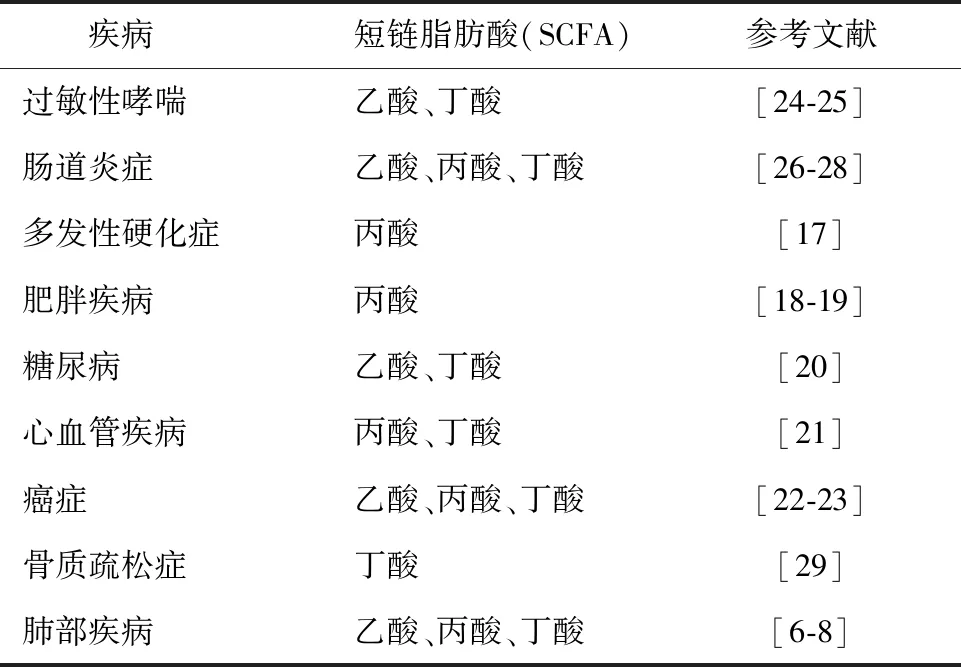
表1 短链脂肪酸与疾病相关性
二、SCFA与肺结核的相关性
SCFA与肺结核之间存在明显的直接相关性[8]。目前验证SCFA与肺结核相关性的方法为:其一,对比肺结核患者与健康对照组的SCFA丰度,验证SCFA与肺结核之间存在关系。常采用的检测方法为:(1)通过液相-质谱联用对血清代谢物检测;(2)通过气相-质谱联用对粪便代谢物检测。其二,建立动物模型,探究SCFA在动物模型中对免疫指标、病理切片等产生的改变,从而验证SCFA可改善肺结核。
研究表明,与健康人相比,肺结核患者的肠道微生物的物种数量和多样性均明显下降,主要表现为SCFA的产生菌以及相关代谢途径均明显减少[30-32]。SCFA的产生菌包括5种丁酸盐产生菌(roseburia inulinivorans、人乳杆菌、肠乳杆菌、直肠真杆菌和球菌)、2种乙酸盐和丙酸盐产生菌(青春双歧杆菌和长双歧杆菌);与SCFA相关的代谢途径包括CENTFERM-PWY、P124-PWY、P461-PWY、PWY-5676和FERMENTATION-PWY。采用抗生素治疗肺结核时会明显影响SCFA(丁酸、戊酸和己酸)的丰度,致使肠道产生不良的影响;为肺结核患者补充干酪乳杆菌可有效调节其SCFA,从而降低抗结核药物引起的肠道损伤[33]。Lachmandas等[34]研究发现,SCFA增加了2型糖尿病患者的肺结核易感性。其研究的丁酸盐生理浓度对人类外周血单核细胞中MTB细胞因子反应表明,丁酸盐降低了MTB诱导的促炎细胞因子的反应,同时增加了白细胞介素-10(IL-10)的产生。这种抗炎作用与丁酸盐对组蛋白脱乙酰酶的抑制作用是相互独立的,并且不伴随Toll样受体信号通路、二十烷类通路或细胞代谢的变化。总之,肺结核导致SCFA的丰度降低,进而影响宿主免疫系统和炎症因子的反应;而宿主免疫系统失调和炎症因子紊乱则会促进肺结核的发展。
部分SCFA自身具有抗结核的作用,可以抑制MTB在机体内的增殖和传播。研究发现,SCFA中的吲哚-3-丙酸(indole-3-propionic acid, IPA)在体内和体外均具有抗MTB的活性。IPA是肠道菌群中的产孢子菌和梭状芽孢杆菌的代谢产物,是人类肠道菌群产生的第一种抗结核代谢产物。IPA治疗活动性结核病小鼠后,其脾脏中的MTB负荷减少到1/7[35]。初步探明的IPA抗结核作用的机制是:色氨酸合成是MTB体内必需的生物合成途径,而IPA通过模拟色氨酸合成抑制剂邻氨基苯甲酸合成酶(anthranilate synthase TrpE)的生理变构,从而阻断MTB中色氨酸的生物合成,抑制了MTB在宿主内繁殖和扩散[36]。
三、SCFA参与调节肺结核的可能机制
目前尚无直接证据证明SCFA参与调节肺结核的作用机制。笔者旨在通过分析既有的研究成果,探究SCFA参与肺结核患者机体调节可能存在的作用机制,以期望为SCFA参与肺结核的调节机制研究提供思路。按照MTB入侵机体的过程,SCFA 对肺结核的调节作用可以划分为两个阶段:阶段一:MTB入侵机体前——SCFA通过发挥对肠黏膜屏障的保护作用来降低MTB的易感性;阶段二:MTB入侵机体后——SCFA通过参与机体的免疫调节来治疗肺结核。
(一)SCFA保护肠黏膜屏障降低MTB易感性
肠黏膜屏障和肠道完整性在降低肺结核易感性方面起着重要作用[37-38]。根据“肺-肠轴”理论,研究表明肠道菌群和肺部菌群的生态平衡是相互影响的。主要表现为:一方面,当肺部由于细菌感染而发生微生态紊乱时,肠道微生态的平衡也会随之而破坏[39]。另一方面,当肠道微生态的平衡遭到破坏时,机体会产生更多的炎性介质,这些炎性介质会通过血液进入肺部,从而引起肺部菌群微生态的紊乱,使得病原菌更容易侵入机体[40]。
SCFA有助于形成强大的肠黏膜屏障,可以防止有害物质通过肠道入侵机体,维持肠道完整性和稳定性,从而维持肺部微生态的平衡,降低结核分枝杆菌潜伏感染的风险[41-43]。根据功能和位置不同,肠黏膜屏障主要分为以下4个部分:机械屏障、化学屏障、免疫屏障和生物屏障。
1.机械屏障:机械屏障又被称为物理屏障,其主要是指肠道上皮的屏障功能。肠道上皮组织主要由肠道上皮细胞及功能不同的紧密连接蛋白组成,紧密连接蛋白使得肠道上皮细胞之间形成了紧密连接,共同保障了肠道上皮屏障功能的完整性。紧密连接蛋白包括跨膜蛋白(闭合蛋白,Claudins;闭锁蛋白,Occludin;连接黏附分子,JAM)和细胞质蛋白(闭合小环蛋白,ZO)两大类。研究表明,SCFA不但可以通过给上皮细胞提供营养的方式促使其增殖,而且可以通过提高紧密连接蛋白的表达,改善肠道物理屏障功能[44-46],具体见图1。
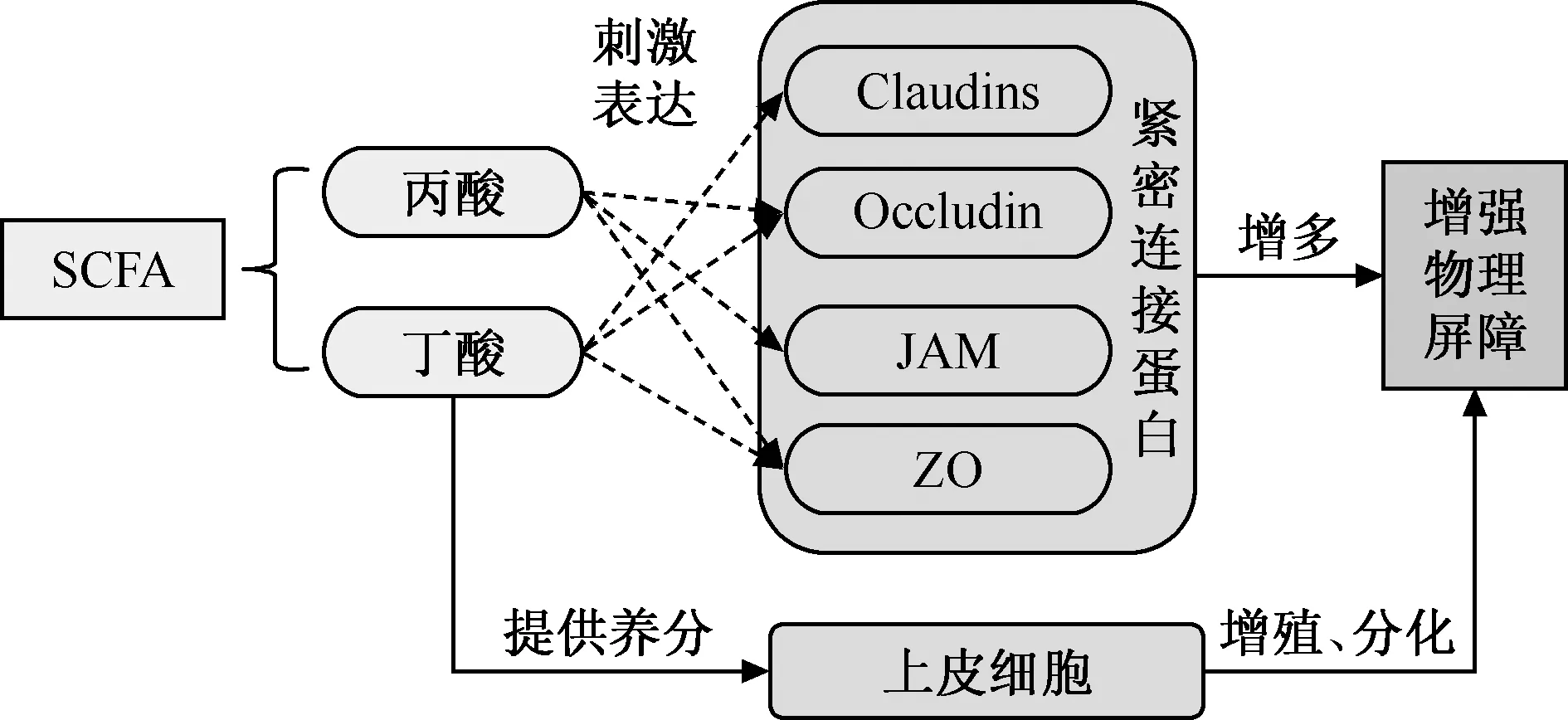
注 SCFA:短链脂肪酸;Claudins:闭合蛋白;Occludin:闭锁蛋白;JAM:连接黏附分子;ZO:闭合小环蛋白
2.化学屏障:化学屏障指的是肠上皮细胞外的黏液层。黏液层对肠道的保护作用主要体现在两个方面。其一,可以发挥黏液层的黏附作用,隔离化学物质、毒素和过敏原[47];其二,黏液层是肠道共生菌的生存场所,可为肠道共生菌提供养分,并且其中存在一种可以把肠道中的病原菌和共生菌与宿主分离开的黏蛋白——MUC2,这种黏蛋白对肠黏膜具有直接的保护作用,进而维持肠道稳态[48]。研究表明,SCFA中的丁酸盐可刺激MUC2的表达,从而增厚黏液层,并增强黏液层的黏附作用,即增强黏液层的化学屏障作用[49]。
3.免疫屏障:肠黏膜固有层中存在大量的淋巴细胞,其可参与肠道免疫调节,形成一道肠道免疫屏障。SCFA可以与肠道中不同的G蛋白偶联受体进行结合,调控淋巴细胞的增殖、分化、凋亡等过程,参与促炎、抗炎因子的产生,从而调控肠道稳态[50]。
4.生物屏障:生物屏障又可以称为“肠道菌群屏障”,由附着在肠道上的菌群以类似于生物膜的方式构成,其通过生物拮抗作用抵御病原菌的入侵。SCFA主要由生物屏障中的肠道菌群产生,并且会反过来对肠道菌群产生影响。SCFA对肠道菌群产生影响的方式主要为调节肠道环境的pH值,从而抑制病原菌的生长,促进益生菌的增殖,调控肠道微生态的平衡。肠道内病原菌的适宜生长环境的pH值大多处于6.0~7.0之间,pH值在4.0以下时会基本失去活性;而益生菌的适宜生长环境的pH值处于3.0~4.5之间。由于SCFA是以SCFA-(阴离子)的状态存在于肠道中,因此,当其浓度升高时会促使产生更多的H+离子,从而使得肠道内环境的酸性增强,pH值降低,抑制肠道内的病原菌的生长或将其杀死,并促进益生菌的生长和增殖,维持肠道微生态的稳定[51]。
(二)SCFA通过参与机体免疫调节抗结核
机体感染MTB后,SCFA通过参与机体免疫调节的方式抗结核。T淋巴细胞(Th)和调节性T细胞(Treg)在机体的适应性免疫调节中发挥着重要的作用,影响MTB在机体内的增殖和传播。Th1细胞可以有效的限制MTB的传播,其产生的γ-干扰素可以激活肺泡中的巨噬细胞[52]。Th17细胞是CD4+T细胞的一个亚群,SCFA诱导募集的Th17细胞可在肺结核患者机体免疫中起到防御的作用[53-55]。近年来研究证明,Th17细胞及细胞因子IL-17可参与Th1/Th2细胞因子功能调节过程,在自身免疫性疾病、抗胞内菌感染等方面有重要作用。SCFA可促进肺内皮细胞趋化因子配体20(CCL20)的表达增加,然后CCL20通过受体6促进Th17细胞募集进入肺部[56]。Th17细胞有利于形成结核保护性免疫,并加速MTB的再次免疫应答,促进中性粒细胞、巨噬细胞和Th1细胞聚集到感染部位,进而清除MTB,并参与机体免疫应答控制感染过程[57]。
Treg细胞是一种抑炎细胞,是CD4+T细胞的另外一个亚群,通过分泌抑制性细胞因子(IL-2、IL-4、IL-8、IL-10和转化生长因子-β)维持免疫稳态[58],抑制肺结核并发的炎症反应。研究表明,丁酸盐可以通过激活表达转录因子Foxp3的表达或通过抑制组蛋白去乙酰化酶的活性来促进初始T细胞分化为Treg 细胞[59-61]。再者,丁酸盐与G蛋白偶联受体相互作用时,会刺激树突状细胞和巨噬细胞分泌更多的IL-10[62],并且巨噬细胞可显示出更高的诱导T细胞分化为Treg细胞的能力[25,27],Treg细胞可以有效抑制肺结核并发炎症过程的发生。
SCFA是肠道菌群有益作用的关键介质。SCFA 还通过与肠黏膜屏障功能、葡萄糖稳态、免疫调节、食欲调节、肥胖相关的许多组织特异性机制直接调节宿主健康。与肺结核相关方面,SCFA可以通过增强肠黏膜屏障的保护作用来降低MTB的易感性,也可以通过调节结核病患者的免疫系统来抑制MTB的增殖和肺结核并发炎症的产生。
四、SCFA的抗结核思路
随着肺结核耐药性的增强及抗结核药物会使患者产生不同程度的不良反应,因此,需要提出一种新的抗结核治疗方法。基于肺-肠轴理论,通过调节肠道菌群中SCFA产生菌的数量,提高SCFA的丰度,从而对预防、辅助治疗或者治疗肺结核产生积极的作用。目前,提高SCFA丰度的方法主要有:(1)生活中自行调整饮食结构;(2)采用口服益生菌、益生元/合成元及粪菌移植的微生物疗法;(3)褪黑素、某些中药成分亦具有提高SCFA丰度的作用。
(一)调整饮食结构
饮食结构直接影响肺结核患者营养状况,而且营养不良者更容易感染肺结核。研究表明,肺结核患者维生素C和锌的摄入不足与肺结核治疗失败率相关[63]。Bhargava等[64]的社会医学调研发现,与罹患活动性结核病的父母一起生活的儿童在得到充分的营养补充后,结核病发病率降低了。也有研究发现,体质量指数与结核病发病率之间存在指数关系[65-66]。低BMI组的新发肺结核发病率是正常BMI组的12倍[67]。
SCFA的生产底物主要为膳食纤维、抗性淀粉、低聚糖,饮食结构可直接影响SCFA的丰度,进而影响肺结核的发病率和治疗效果。膳食纤维摄入不足可能会增加对炎症性疾病的易感性[68],而多摄入富含膳食纤维的蔬菜、水果及粗粮可缓解肺结核的症状。Yang等[69]给仔猪喂食高脂高纤维饮食后发现,其代谢产物SCFA明显增加,且高脂高纤维饮食可以减少肺部损伤,促进肺功能恢复。
部分研究表明,茶叶可增加肠道中SCFA的丰度。叶颖等[70]发现六堡茶水提取物能显著增加大鼠肠道中的丙酸和丁酸含量,对两种SCFA的促进率分别达到了24.81%和23.03%。Gao等[71]发现红茶可以增加Sprague-Dawley大鼠肠道菌群的α-多样性,调节了β-多样性,增加SCFA的丰度,增强肠屏障功能。
(二)微生物疗法
近年来,广谱抗生素如异烟肼(INH)、利福平(RFP)、吡嗪酰胺(PZA)、乙胺丁醇(EMB)等已经被成功用于肺结核的治疗。然而,药物不良反应及肺结核耐药性增强使得通过广谱抗生素治疗肺结核仍然具有很强的局限性[72]。因此,越来越多的学者提出采用微生物疗法辅助治疗肺结核。微生物疗法是近年来在微生物与机体生理病理相关性研究基础上发展而来的一种新型疾病治疗方法,目前在糖尿病[73]和肿瘤[74-75]的治疗方面取得了一定的进展。目前,常见的微生物疗法有:口服益生菌、口服益生元/合成元和粪菌移植。
1.口服益生菌:口服益生菌治疗即让患者口服活或死的单个或多个菌株,当补充的数量足够时,即可起到治疗肺结核的作用。研究表明,由于抗生素治疗造成的肺结核患者肠道微生态失调,可在益生菌干预两周后趋于正常[76],且益生菌辅助治疗可以明显减少MTB的定植[77]。此外,研究人员通过小鼠实验证明了补充乳杆菌可以在一定程度上降低肺结核带来的肝损伤[78],也保护了胃肠道黏膜,且具有一定的抵抗病原菌感染的能力[79]。Yoon等[80]从泡菜中分离出来一种乳酸葡萄球菌(Pediococcus acidilactici PMC202),该益生菌具有抗肺结核的作用。
2.口服益生元/合成元:益生元指的是某些不被消化或者难以消化的食物残渣,其可以定向刺激某些益生菌的产生。合成元指的是益生元和益生菌的组合物,两者可以协同对机体发挥作用[81]。研究表明,相比于单独使用益生菌或者益生元,合成元具有更好的调节肠道菌群的效果[82]。然而,益生菌、益生元/合成元调节肠道菌群的作用机制及治疗效果是否与使用剂量、菌种比例、益生菌和益生元比例相关等问题还需要进一步探究。
3.粪菌移植:粪菌移植是指将健康供者的粪便放入患者的胃肠道,调节患者肠道微生态,从而治疗疾病的一种新型治疗技术[83]。具体来说,粪菌移植的核心意义是从健康人或动物的粪便中经过筛选、离心、过滤等步骤,提取有益的微生物群,移植到相应受体的胃肠道中,从而重塑受体的肠道微生态并使其正常发挥作用,进而预防或治疗肺结核[84]。粪菌移植已成功应用于肠道疾病的治疗,包括肠易激综合征、艰难梭菌感染、结肠癌等[85-87]。特别是艰难梭菌感染的治愈率高达90%,且较少产生不良反应。粪菌移植是目前公认的一种安全有效、不良反应少的治疗方法,且每个受体均可在短时间内成功定植健康肠道菌群。但是单次粪菌移植后受体的菌群维持时间一般为3~6个月,因此在部分菌群消失后需对受体再次进行粪菌移植[88-89]。鉴于个体差异性和菌群存活时间等问题,需要在粪菌移植后,对受体进行长期的临床治疗效果监测,以合理地确定再次粪菌移植的时间。粪菌移植理论上可以应用于临床肺结核的治疗,但相关应用方法及安全性仍需进一步讨论。
(三)褪黑素及某些中药成分
褪黑素可以改善动物和人类的脂质代谢和肠道菌群。小鼠口服补充褪黑素后,缓解了其脂质积累,并改善了其肠道菌群失调,包括肠道菌群多样性、拟杆菌属和Alistipes的相对丰度。在高脂肪饮食喂养的小鼠中,SCFA减少,而褪黑素治疗后,则提高了乙酸的产生。且相关性分析发现,乙酸的产生与拟杆菌类和Alistipes的相对丰度之间存在明显相关性[90-91]。
某些中药成分被证明可以提高SCFA的丰度。大鼠实验表明,女贞子可增加乙酸盐、丙酸盐、丁酸盐的浓度[92];杜仲叶可增加肠道厚壁菌门和其他SCFA产生菌的丰度,增加乙酸和异丁酸的生成量[93];葛根可提高SCFA水平和修复肠黏膜完整性[94];黄连可提高丁酸盐产生菌的丰度[95];淫羊藿可提高普雷沃菌属和拟杆菌属的丰度,从而提高SCFA的丰度[96];刺桐皮可提高乙酸盐和丁酸盐的丰度[97]。
五、小结与展望
综上所述,SCFA作为肠道菌群的代谢产物与肺结核密切相关,在其发生发展过程中发挥着重要的作用。肺结核导致SCFA丰度降低,进而影响宿主免疫系统和炎症因子的反应;而宿主免疫系统失调和炎症因子紊乱则会促进肺结核的发生和发展。然而,目前SCFA参与调节肺结核的作用机制尚不明确,且相关微生物疗法的治疗过程、作用机制和治疗效果也尚待进一步研究。因此,加强SCFA参与调节肺结核的作用机制研究,探究不同种类和丰度的SCFA对肺结核转归的影响,可为微生物疗法在肺结核中的成功应用提供理论依据,从而解决传统药物不良反应和MTB耐药性的问题。
利益冲突所有作者均声明不存在利益冲突
作者贡献张晓萌:文献检索、资料收集整理和文稿撰写;李敏、柴英辉和周静:资料收集整理、文稿修订和编辑;雷红:确定研究主题、项目分析、资金支持、文稿修订、审核和编辑
猜你喜欢
科学导报(2023年82期)2023-12-02 12:08:36
江苏安全生产(2022年8期)2022-11-01 09:14:32
中国饲料(2021年17期)2021-11-02 08:15:10
小资CHIC!ELEGANCE(2021年36期)2021-10-15 14:36:34
山东第一医科大学(山东省医学科学院)学报(2021年7期)2021-10-13 06:41:06
当代水产(2021年2期)2021-03-29 02:57:48
传染病信息(2021年6期)2021-02-12 01:52:14
艺术评鉴(2020年5期)2020-04-30 06:47:57
广州医科大学学报(2020年1期)2020-03-03 03:58:12
分子影像学杂志(2019年3期)2019-03-25 12:09:39
