
催产素受体在山羊结状神经节的分布
2023-06-26 14:04陈文东朱秀娟何玉鹏徐永平
西北农业学报 2023年7期
陈文东,朱秀娟,何玉鹏,王 昱,徐永平
(1.陇南师范高等专科学校 农林技术学院,甘肃成县 742500;2.西北农林科技大学 动物医学学院,陕西杨凌 712100)
催产素(Oxytocin,OT)是在动物机体下丘脑室旁核(Paraventricular nucleus)和视上核(Supraoptic nucleus)中产生的一种九肽类激素,其分泌主要受神经反射弧调节,经神经垂体释放进入血液或脑脊液中,在血液循环中以自由肽的形式存在,催产素的功能是通过其与催产素受体(Oxytocin receptor,OTR)的特异性结合激活信号通路后而实现的。Caldwell 等[1]发现了一些新的催产素受体表达位点,包括内皮细胞、胰岛细胞、几种类型的癌细胞、脂肪细胞、心肌细胞、骨细胞和成肌细胞,这些发现扩大了传统催产素的作用不仅仅是子宫收缩和排乳反射诱导物的范围,当应用催产素时,这些不同位点的催产素受体表达可以激活各自组织胞内信号转导通路[2]。最新对OTR定位数据表明,催产素受体在消化道内广泛存在,比如肠吸收上皮细胞和肠腺上皮细胞,平滑肌和肠神经细胞特别是肌间神经丛,催产素可促进人类的胃排空,催产素受体抑制剂延迟人类的胃排空[3],但是,催产素延迟鼠的胃排空[4],表明催产素可以以内分泌调节的方式对胃肠道发挥 作用。
结状神经节(nodose ganglion,NG)中的神经细胞为假单极神经细胞,这些假单极神经细胞分为两部分,下行树突为分布在大范围消化道、心血管和呼吸器官的感觉神经末梢,是迷走神经的感觉神经纤维成分,上行树突(中枢突)向脑干投射[5],在胃肠道、呼吸器官和心血管内形成的信号可以通过迷走神经传递到脑干。有文献报道食道、心、肺、胃和肠等内脏传入神经细胞主要分布在结状神经节内。贲门、幽门和胃底等部位的感觉传导是通过迷走神经完成的[6-7]。关于催产素通过外周神经对内脏活动影响的研究中,陈文东等[8-9]和任妮等[10-11]运用免疫组织化学技术分别在腹腔肠系膜前、后神经节,颈前神经节和颈胸神经节上发现了催产素受体的分布,并推断催产素除了以分泌方式对一些内脏器官发挥调节作用外,还有可能通过影响支配这些内脏器官的外周交感神经细胞活性而发挥神经调节作用,这表明催产素可能通过内脏器官反射弧上的传出神经纤维对内脏器官发挥调节作用。那么催产素是否同样可通过与NG内催产素受体特异性结合,从而通过内脏器官反射弧上的传入纤维对内脏器官发挥神经调节作用呢?本研究采用免疫组化方法检测山羊NG内催产素受体的表达分布特点,为探索山羊NG是否具有成为OT对内脏器官进行内分泌调节和通过自主神经对内脏器官进行神经调节两者相互协调作用节点的条件提供形态学依据。
1 材料与方法
1.1 试验动物
杨凌农户养殖的成年关中奶山羊,雌雄各5只,健康,雌性未怀孕。
1.2 试剂及仪器
免疫组化SP超敏试剂盒和内源性生物素阻断试剂盒(迈新生物);一抗兔抗OTR多克隆抗体(北京康为世纪);四盐酸3,3-二氨基联苯胺(DAB)试剂盒(金桥生物);Leica石蜡切片机;显微成像系统(赛默飞世尔)。
1.3 试验方法
颈总动脉放血致死山羊,找出颈部迷走交感干,小心分离迷走神经和交感神经,然后顺着迷走神经向胸部找到左右两侧结状神经节,在靠近神经节处剪断上下连接的神经纤维,取下结状神经节,迅速放入装有4%多聚甲醛溶液的广口瓶里固定。按陈文东等[8]方法制作石蜡切片。制作出的石蜡切片分别进行HE染色和免疫组织化学染色,HE染色用于观察NG组织形态学特点,其染色步骤为:脱蜡,梯度酒精复水,苏木精染色,自来水反蓝,伊红染色,酒精脱水,二甲苯透明,最后中性树胶封片。用于免疫组织化学染色的石蜡切片分成3 组,第一组为免疫组织化学SP法染色,用于OTR阳性染色定位及图像分析,OTR阳性着色为棕褐色、黄褐色或淡黄色,第二组为免疫组织化学SP法染色,苏木精复染,用于观察OTR阳性细胞分布特点,第三组为空白对照,用于确定OTR染色的特异性。免疫组织化学SP法染色步骤为:石蜡切片经过脱蜡,复水,抗原修复后,按免疫组化SP超敏试剂盒说明书进行染色,试剂盒A液(内源性过氧化物酶阻断剂)、B液(羊非免疫血清)、C液(生物素标记的羊抗兔IgG)和D液(链霉菌抗生物素蛋白-过氧化酶)在湿盒内分别孵育15、15、20、20 min,第一组在B液之后滴加兔抗OTR多克隆抗体(以1∶200的体积比稀释),4 ℃冰箱过夜,D液之后加DAB显色5 min,以上每个步骤完成后均需用0.01 mol/L PBS充分漂洗(连续3次,每次5 min)。第二组在免疫组化SP法染色后用苏木精复染,其他步骤同SP法染色,第三组在B液孵育后滴加抗体稀释液替代兔抗OTR多克隆抗体,其他步骤同第二组。以上切片经常规脱水、透明、封片后,Motic生物显微镜下观察,拍照,并记录试验结果。
1.4 图片分析及统计分析
用motic生物显微镜观察免疫组织化学染色后的组织切片,并在显微成像系统下拍照。选出10张免疫组织化学切片,每张切片再随机拍5张400×照片,用Image-pro plus 6.0对照片进行光密度测定,操作步骤为首先设置光密度校正、测量项目(IOD和area)和分色选择参数,然后在Count/Size窗口选择Manual选项,点Select Colors调出segmentation工具选择AOI (area of interest),最后点击Count测定结状神经节神经细胞和非神经细胞结构的平均光密度(平均光密 度=光密度累加值IOD/目标分布区域面积 area)[12]。用SPSS 20分析数据,数据表示形式为“平均数±标准差”,用单因子方差分析(One- way ANOVA,LSD)法检验比较结状神经节内神经细胞和非神经细胞结构的OTR分布差异性。
2 结果与分析
2.1 HE染色结果
由图1可知,结状神经节中可见神经细胞成群分布,相对于其他细胞,神经细胞体积较大,胞体结构各异,各神经细胞之间及神经细胞与周围组织之间边界清晰,结构完整,还可见部分无核神经细胞。神经细胞胞质着色为紫红色;胞核呈圆形或卵圆形,在神经细胞的边缘或正中央位置,着色为蓝色,少部分胞核内可见核仁结构,染色较核质颜色更深,此外可见到部分多核或双核神经细胞。神经细胞周边有支持细胞环绕,其形态扁平状,结构清晰。在众多神经细胞之间有神经纤维和微血管通过,神经纤维上包裹有雪旺细胞,胞核呈长卵圆状;微血管内壁上的血管内皮细胞清晰可见,微血管管腔内可见有少量血细胞存在,着色为红色。
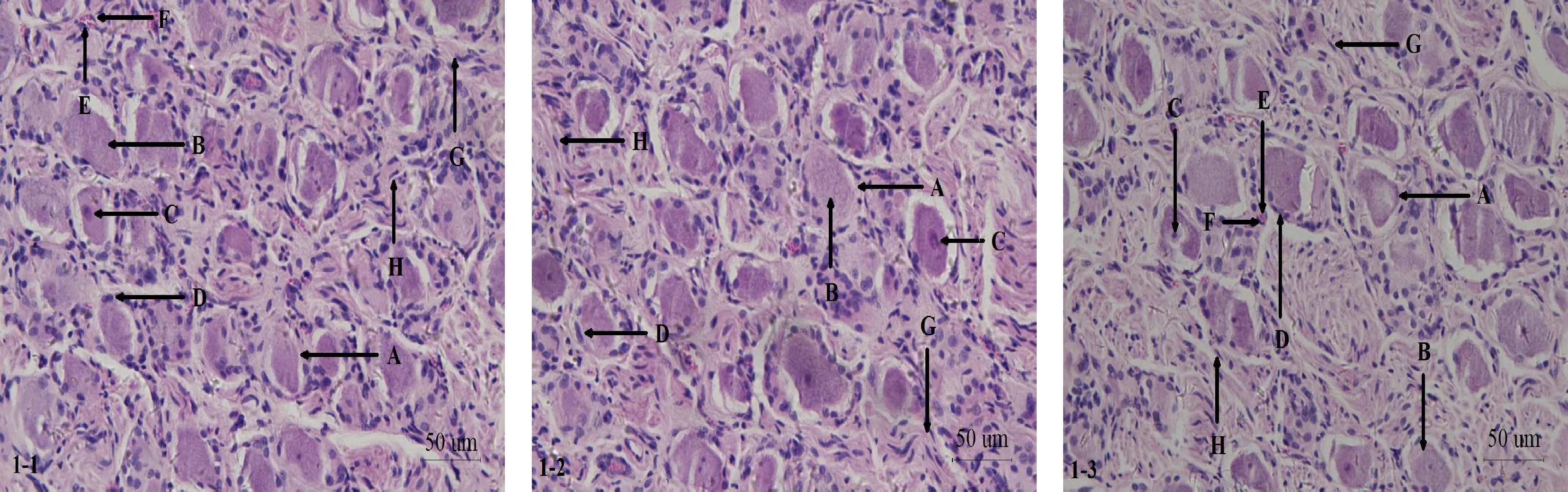
1-1.免疫组织化学SP法对照组山羊结状神经节HE染色;1-2.免疫组织化学SP法染色及苏木精复染组山羊结状神经节HE染色;1-3.免疫组织化学SP法染色组山羊结状神经节HE染色;A.神经细胞;B.神经细胞胞质;C.神经细胞胞核;D.支持细胞;E.微血管;F.血管内皮细胞;G.过路神经纤维;H.雪旺细胞
2.2 免疫组织化学染色结果
对照组切片复染结果(图2-1)可见,神经细胞胞体着色为浅紫色,神经细胞胞核着色为深紫色,整个切片背景为浅蓝色,没有其他着色。试验组切片复染结果(图2-2)可见,神经细胞胞体着色为棕褐色或黄褐色,其他非神经细胞结构胞核着色为紫色,证明催产素受体免疫组化SP染色结果没有非特异性着色。
免疫组织化学染色结果(图2-3)可见,结状神经节各种细胞着色深浅不一,根据着色深浅将OTR表达分为4 个等级:棕褐色为强阳性;黄褐色为中等阳性;淡黄色为弱阳性;无着色为阴性。试验组免疫组化SP染色结果可知,结状神经节内OTR免疫阳性产物分布广泛,神经细胞、过路神经纤维、血管内皮细胞、支持细胞和雪旺细胞均有不同程度的着色。神经节内着色神经细胞大小不等,最大神经细胞直径达61 μm,最小直径为32 μm(测量图片内有核神经细胞,Image-pro plus 6.0测量)。神经细胞的胞核和支持细胞着色最深,为棕褐色,OTR分布强阳性。大部分神经细胞胞质(图2-B)着色为黄褐色,OTR分布中等阳性,这部分神经细胞胞质约占神经细胞总数的78%。血管内皮细胞、细胞膜,以及一小部分神经细胞胞质着色较浅,为淡黄色,OTR分布弱阳性。神经细胞胞核的核仁呈圆形,着色极浅或不着色,OTR分布阴性。

2-1.OTR的免疫组织化学SP法对照组及苏木精复染;2-2.OTR的免疫组织化学SP法染色及苏木精复染;2-3.OTR的免疫组织化学SP法染色;A.神经细胞;B.神经细胞胞质;C.神经细胞胞核;D.支持细胞;E.微血管;F.血管内皮细胞;G.过路神经纤维;H.雪旺细胞
2.3 NG中OTR的表达量
图像分析结果表明,结状神经节中神经细胞内的OTR表达量极显著大于非神经细胞结构 (P<0.01,表1)。

表1 结状神经节OTR免疫组化染色的平均光密度值Table 1 Mean density of OTR stained by immunohisto chemistry in nodose ganglions
3 讨 论
OT是第一个被确定生物活性和化学结构的性激素,它经垂体后的下丘脑神经终末进入血液循环,可以加强雌性动物子宫收缩和促进乳腺排乳。Jankowski 等[13]研究发现,大鼠皮下注射不同剂量的OT可引起血浆催产素浓度在生理学范围内升高,增加低钠饮食大鼠的尿钠排泄作用,且这种作用可以被OTR抑制剂所阻断,证明OT诱导的钠尿排泄是依赖OTR而实现的。研究发现,人类和大鼠的所有心脏腔室、血管内皮细胞、血管平滑肌细胞[14]和肾脏[15]均有催产素受体存在,OT通过G1或Gq蛋白偶联通路与这些受体相互作用调节心率和心肌收缩力、舒缩血管,以及调节肾小球的血流量。催产素诱导豚鼠胃窦平滑肌细胞放松[16],且降低狗和大鼠的胃动力[17-19]。上述资料表明,催产素可以直接与相应组织器官上的催产素受体特异性结合而发挥调节作用。OTR还存在于大鼠胰腺内胰岛周边及颈动脉体内[20],因此,OT可调节胰岛素或胰高血糖素的产生而间接对大鼠血糖指数产生影响。
有关催产素受体在人类体内分布的研究发现,催产素受体基因位于3p25和3p26染色体区域上,包含4 个外显子和3 个内含子,跨越19 210 个碱基对,其在人类下丘脑、杏仁核和海马等“社会”脑区分布密度最大[21],单剂量催产素给药时可显著提高孤独症谱系障碍(ASD)患者对面部表情的识别能力、提高采访者面部眼睛区域的注视点数和注视时间、增加社会注意加工等[22]。鼻吸催产素可以增加人类个体对面孔眼区部分的注意,增加自我知觉,提高男性的情绪共情和观点采择能力,调节人类的应激反应,提高对morphing面孔图片中自我面孔的识别边界[23]。研究表明,除了盲肠和胆囊外,人类胃肠道的各部分均表达催产素和催产素受体的mRNA[24],人类催产素全身用药可增加胃排空,吸入催产素可以使患有严重慢性便秘的妇女排便次数增加一倍[25]。以上资料表明,催产素可与人类中枢神经系统及外周组织内分布的催产素受体特异性结合而发挥调节作用。此外,催产素受体在人的成骨细胞和破骨细胞表面也有存在,催产素通过与这些受体结合而对骨代谢发挥调解作用[26-27]。
OTR在动物中枢神经系统中广泛表达分布[28],对于啮齿动物类,OTR主要在嗅球和嗅结节、新皮层、海马结构、中央和外侧杏仁核、伏隔核、终纹床核和下丘脑腹内侧核表达分布[29-30]。催产素通过与上述部位的催产素受体特异性结合发挥减少焦虑和压力,促进社会交往、孕产妇保健和避免条件性逃避行为。Huber 等[31]对中央杏仁核的急性脑切片应用高度特异性的OTR激动剂,可使224个神经细胞中21%的自发峰值频率增加,50%的神经细胞峰值频率下降,且两种反应是完全可逆的、可重复的,而且应用OTR阻断剂对这种反应具有拮抗作用。Yoshida 等[32]发现,中缝核中相当数量的血清胺神经细胞表达OTR,这些受体可能通过刺激血清素释放发挥抗焦虑作用。直接将OT注入到大鼠脑室或侧脑室内,动物表现出明显的镇痛功效,诱发丘脑束旁核内关于痛兴奋的神经细胞放点频率表现一定程度的降低,缩短关于痛的抑制性神经细胞放电时间[33],以上作用均在OTR的介导下完成的。
催产素受体在外周神经也有表达分布,GABA受体和催产素受体可共存于背根神经节细胞膜,在动物处于某些刺激条件下的时候,垂体释放OT与背根神经节细胞膜上的OTR结合,在脊髓水平对感觉信息,特别是痛觉信息进行调节[34]。有研究表明,催产素的鞘内给药可激活存在于脊髓交感节前神经细胞的催产素受体,而起到增加心率,引起血压轻微升高的作用[35]。支配阴茎的副交感神经节前神经细胞有催产素受体存在,催产素注入脊髓副交感神经核可引起大鼠阴茎勃起,阻断催产素受体可去勃起作用[36-37]。以上资料提示,催产素可通过与神经细胞内的催产素受体特异性结合,从而通过神经细胞纤维对支配器官发挥作用。本研究通过免疫组织化学试验技术证实,OTR免疫阳性产物在山羊结状神经节内广泛表达分布,且结状神经节神经细胞内的OTR阳性产物表达量极显著大于其他非神经细胞结构(P<0.01)。表明催产素受体主要表达分布在山羊结状神经节内脏感觉神经细胞中,内脏感觉神经细胞具备对催产素刺激作出反应的条件,提示OT发挥作用的主要靶细胞为山羊结状神经节中的内脏感觉神经细胞。盆腔和腹腔内大多数脏器的内脏痛觉传入主要来自交感神经,迷走神经感觉纤维与血管运动反射、内分泌反射和特殊的内脏运动反射等有关系[38]。NG内的内脏传入神经细胞中枢突与脑干连接,与孤束核、三叉神经脊束核、迷走神经背运动核和网状巨细胞核等核团有广泛的联系[39]。内脏传入冲动可以引起内脏活动改变,还可引起内脏躯体反射(即引起躯体的活动)。因此,调整内脏活动主要受源于内脏感觉神经冲动的改变来完成。本试验采用免疫组织化学SP法发现山羊NG中催产素受体广泛分布表达,表明山羊NG具有OT对内脏器官进行内分泌调节和通过自主神经对内脏器官进行神经调节两者相互协调作用节点的条件,但催产素通过自主神经对内脏器官活动产生什么样的影响,还有待进一步研究。
4 结 论
OTR免疫阳性产物在山羊NG中广泛表达分布,且主要分布在NG内脏感觉神经细胞内,提示催产素有可能作用于NG内脏感觉神经细胞,从而通过内脏反射弧的传入神经纤维影响内脏器官活动,而NG可能成为OT对内脏器官进行内分泌调节和通过自主神经对内脏器官进行神经调节两者相互协调作用的节点。
猜你喜欢
妇女之友(2020年9期)2020-10-26
理化检验-化学分册(2020年5期)2020-06-15
37°女人(2019年4期)2019-04-20
猪业科学(2018年4期)2018-05-19
大科技·百科新说(2017年8期)2017-09-21
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中国医学科学院学报(2015年5期)2015-03-01
发明与创新(2015年1期)2015-02-27
现代检验医学杂志(2014年5期)2014-02-02
