
互花米草入侵对漳江口红树林保护区的影响研究
2023-06-25 18:48:40黄央央杨毕铖戴桂香
海洋开发与管理 2023年2期
黄央央 杨毕铖 戴桂香

关键词:漳江口红树林保护区;互花米草;入侵过程
中图分类号:X171.1;X36;P745 文献标志码:A 文章编号:1005-9857(2023)02-0115-10
0 引言
互花米草(Spartinaalterniflora)源于北美东海岸和墨西哥湾,是多年生的草本植物,属禾本科、米草属,适宜生活在潮间带和沼泽地。互花米草根系发达,在潮间带泥沙的快速沉降和游积过程中可以发挥重要的作用,由于无意带入或有意引种,互花米草已扩张到世界范围内诸多滨海和河口地区,威胁全球的海滨湿地本土物种,作为全球性入侵种引起广泛关注和研究[1]。我国于1979年12月从美国引进互花米草,在人为推广和自然力量的扩散下广为传播,因其强适应性和扩散能力,互花米草从引种地迅速蔓延,占据中国沿海北起辽宁、南至雷州半岛的广大滩涂[2],已成为我国潮间带分布最为普遍的盐沼植物[3],2003年被列入16 种中国首批外来入侵种名单。鉴于互花米草的入侵性和危害性,其对我国海岸带的入侵现状及生态效应已成为近十数年来的研究热点[2,4]。
福建漳江口红树林国家级自然保护区是中国红树林自然地理分布最北的重要湿地类型保护区,也是福建省最重要的湿地生态系统类型的国家级自然保护区[5],主要红树林类型有海榄雌林、桐花树林、海榄雌+ 桐花树、秋茄林、秋茄+桐花树林、木榄林。根据已有研究,2005—2017年漳江口保护区内互花米草面积增加超过1倍,主要分布于中部和西部滩涂[6]。本研究通过漳江口红树林保护区互花米草区和对照组(红树林区、光滩)互花米草生长状况、沉积物环境和大型底栖生物群落差异,结合历史遥感影像数据分析互花米草的入侵过程,以期为自然保护区科学管控提供重要信息,为预测互花米草潜在分布区域提供科学基础。
1 材料与方法
1.1 断面设置
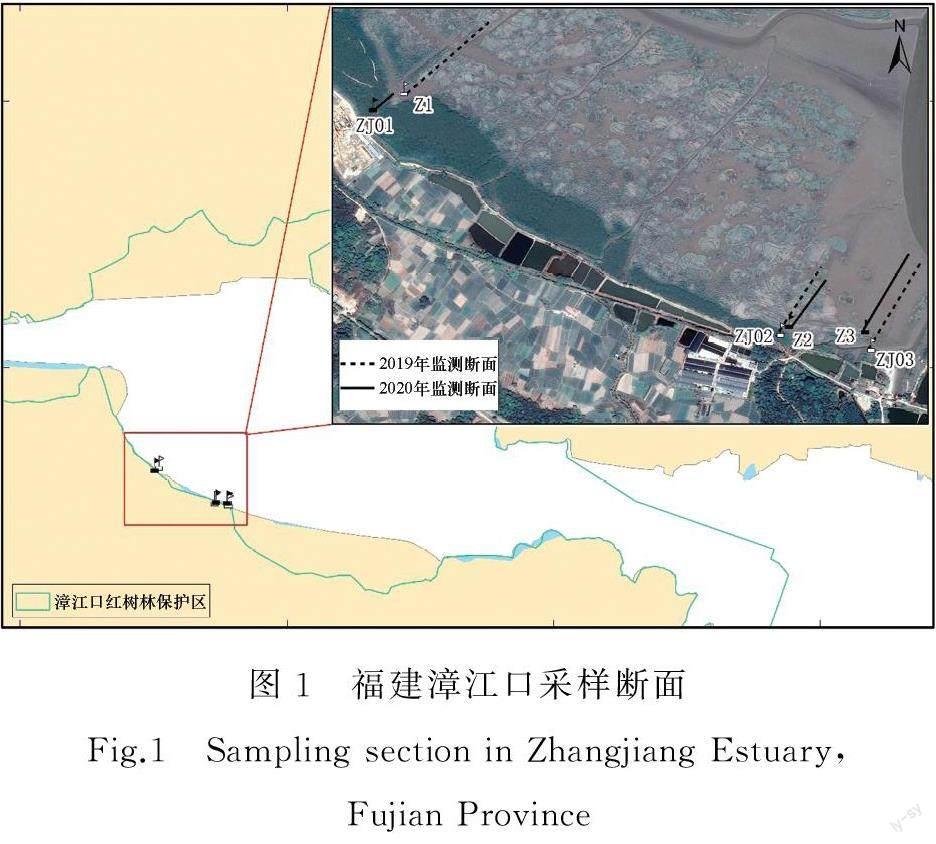
2019年、2020年10月分别在漳江口红树林保护区选择3 条较为典型的断面。其中,Z1~Z3 为2019年调查断面,ZJ01~ZJ03为2020年调查断面。其中,ZJ01位于红树林区(本研究断面位于竹塔村附近,红树林区主要为多种红树组成的群落区),Z3和ZJ03位于光滩,其余断面位于互花米草区。2019年互花米草、光滩调查断面各采集高、中、低潮带样方数量各1个,开展互花米草和沉积物监测;2020年互花米草调查断面采集高、中潮带样方数量各1个,开展互花米草和沉积物监测。监测断面如图1所示。
1.2 样品采集与分析
1.2.1 植被
植被监测选择在10月进行采样,主要调查互花米草区域植被种类组成、高度、数量、盖度等要素。其中,盖度调查方法采用目视法;高度用米尺从地面測量至样格内优势种顶端,取3株代表性强植物的平均值,再次记录样格内植物种类,最后进行样格内植株数量调查,现场记录样格内各类盐沼植物的数量。具体调查采样主要按照《海岸带生态系统现状调查与评估技术导则第4 部分:盐沼》(T/CAOE20.4-2020)相关要求执行。
1.2.2 沉积物
沉积物监测采样时须将环刀口刃向下垂直压入土中,直至环刀筒中充满土样为止,取出已经充满土的环刀,削去两端多余的土壤,将环刀内的土壤装入自封袋内封闭好带回实验室。现场记录沉积物类型、测定沉积物氧化还原电位,实验室分析有机碳、全盐含量、总氮、总磷等指标。氧化还原电位使用便携式pH 计(型号:pH3210)测定,具体调查采样主要按照《海洋调查规范》(GB/T12763-2007)相关要求执行;全盐含量调查采样主要按照《土壤检测》(NY/T1121-2006)相关要求执行;有机碳、总氮、总磷调查采样主要按照《海洋监测规范》(GB17378-2007)相关要求执行。
1.2.3 潮间带底栖生物
潮间带底栖生物调查在植被样方的高潮带、中潮带各设置1个站位,采集底栖生物定量和定性样品。定量样品在植被调查样方内采集,调查位置与植被调查和沉积物调查相一致;定性样品在样方周边采集,并尽可能捕捉全部类型生物。采集的样品均加甲醛溶液固定。具体调查采样主要按照《海洋监测规范》(GB17378-2007)和《海洋调查规范》(GB/T12763-2007)相关要求执行。
1.3 数据分析
1.3.2 ABC曲线绘图及W 值
用PRIMER6软件绘制漳江口各植被样方的潮间带底栖生物丰度和生物量的k-优势曲线(ABC曲线),分析生物群落受污染或其他因素扰动的情况[8],将群落分为未干扰、中等干扰和严重干扰3种状态[9]。其中,生物量优势度曲线在数量优势度曲线上方时,群落处于未干扰状态,以生长慢、性成熟晚的大型物种为主;生物量、数量优势度曲线相交时,群落处于中等干扰状态;生物量优势度曲线在数量优势度曲线下方时,群落处于严重干扰状态,以生长快、性成熟早的小型物种为主[10-11]。W 的取值范围为[-1,1],当W 值为1时预示着物种的丰度值均匀,但生物量是单一生物占优势,当W 值为-1时则相反[12]。
1.3.3 遥感影像选取与数据分析
本研究从91卫图等相关遥感软件选取漳江口区域2011 年4 月24 日、2015 年10 月17 日、2017年10月27日、2019年10月和2021年9月的高分辨率遥感影像数据(空间分辨率小于1m,投影坐标系统为GCS_ WGS_1984,中央经线选取117.5°),运用ArcGIS10.6软件对高分辨率遥感影像上的互花米草区进行人机交互识别,形成矢量化数据,分析互花米草扩散过程,计算其扩散面积。
2 结果与分析
2.1 互花米草群落特征
2019年、2020年漳江口互花米草植被调查结果见表1,互花米草的平均高度为1.27~1.43m,植株密度为188~378株/m2。从调查结果来看,漳江口高潮区互花米草的生长状况总体比中潮区好,但高潮区的株高低于中潮区。
2.2 沉积环境质量对比分析
2019年和2020年漳江口不同植被样方区域沉积物要素对比见图2(2020年低潮区互花米草区无数据)。2019年和2020年漳江口互花米草区沉积物的氧化还原电位由高到低依次为高潮区、中潮区、低潮区,光滩的氧化还原电位在各潮区变化不大。2019年漳江口互花米草区沉积物的全盐含量由高到低依次为低潮区、中潮区、高潮区,各潮区全盐含量均低于光滩;2020年漳江口互花米草区沉积物全盐含量由高到低依次为中潮区、高潮区,光滩沉积物中全盐含量由高到低依次为中潮区、低潮区、高潮区;与2019年相比,2020年互花米草区沉积物全盐含量降低26.6%~37.0%。
2019年漳江口互花米草区沉积物的硫化物、总磷含量由高到低依次为高潮区、中潮区、低潮区,光滩则由高到低依次为高潮区、中潮区、低潮区;2020年互花米草区沉积物的总磷含量则呈现中潮区高于高潮区,但总体互花米草区各潮区总磷含量相差不大。2019年和2020年漳江口互花米草区和光滩沉积物的总氮含量由高到低依次为中潮区、高潮区、低潮区,互花米草区沉积物总氮含量高于光滩。2019年漳江口互花米草区沉积物的有机碳含量由高到低依次为高潮区、中潮区、低潮区,光滩则由高到低依次为低潮区、中潮区、高潮区,这与王爱军等[13]研究结果一致;2020年漳江口互花米草区沉积物的有机碳含量中潮区略高于高潮区,但相差不大。与2019 年相比,2020年漳江口互花米草区高潮区和中潮区总磷含量减少68.1%~76.0%,总氮含量增加40.8%~50.5%,有机碳含量增加20.4%~25.2%。
另外,课题组研究了2019年泉州湾互花米草区和光滩的硫化物含量情况,发现泉州湾互花米草区硫化物含量由高到低依次为中潮区、高潮区、低潮区,光滩则呈现相反规律,2个研究区域的互花米草区沉积物硫化物含量显著高于光滩,其中漳江口的差异更明显,尤其是高潮区[14]。由于互花米草有较高的硫养分累积能力和养分归还能力,互花米草入侵后,湿地土壤的沉积环境、酸碱度、有机质、可溶性盐和土壤水分等发生重大改变,会导致土壤硫库增加,这种变化在入侵初期会更加显著[15-16],由此可见2019年漳江口的互花米草仍处于高速扩张期。
2.3 潮间带底栖生物多样性和群落结构稳定性
2.3.1 种类组成
2019年和2020年研究区域互花米草区、红树林区和光滩的潮间带底栖生物种类组成见图3。潮间带底栖生物种类数由高到低依次为红树林区、光滩、互花米草区。2019年漳江口互花米草区的潮间带生物主要类群未见多毛类,生物类群单一;2020年研究区域互花米草区的潮间带生物出现多毛类。
2.3.2 总密度、总生物量
2019年和2020年研究区域的潮间带底栖生物栖息密度和生物量变化如图4所示。2個航次研究区域的潮间带底栖生物的栖息密度由高到低依次为红树林区、光滩、互花米草区;潮间带底栖生物的生物量由高到低依次为互花米草区、光滩、红树林区,与栖息密度呈现相反的规律。
2.3.3 优势种
互花米草区、光滩、红树林区的潮间带底栖生物优势种类群均有所不同,具体见表2。2019年互花米草区主要优势种均为甲壳动物,2020年互花米草区优势种增加多毛类,其中2019年宁波泥蟹占较大优势(Y =0.27),2020年褶痕相手蟹占较大优势(Y=0.28)。光滩的优势种以甲壳动物、软体动物和多毛类为主,2019年和2020年光滩潮间带底栖生物优势种组成略有不同,均以甲壳动物占较大优势,其中2019年畸形鎚肢虫占较大优势(Y=0.37),2020年秀丽长方蟹占较大优势(Y =0.52)。红树林区优势种以甲壳动物、软体动物和多毛类为主,但优势种的优势度差距较小,Y 值为0.16~0.02,物种间分布较均匀。
2.3.4 物种多样性指数
2019年和2020年漳江口不同植被样方区域潮间带底栖生物物种多样性指数变化如图5 所示。2019年漳江口湿地互花米草区多样性指数和丰度指数均低于光滩。2020年研究区域的多样性指数和丰度指数由高到低依次为红树林区、互花米草区、光滩。2020年互花米草区多样性指数和丰度指数的变化幅度较2019年大。
2.3.5 群落结构稳定性
应用丰度生物量比较法对2个航次漳江口光滩、互花米草区和红树林区的潮间带底栖生物群落结构进行分析(图6)。结果表明:2019年漳江口光滩和互花米草区潮间带底栖生物的生物量曲线起点均位于丰度曲线下方,光滩在中部交叉,互花米草区在尾部交叉,表明2019年研究区域的底栖生物受到中度干扰,互花米草区W 值为-0.1508,干扰程度更深。2020年除光滩外,互花米草区和红树林区潮间带底栖生物的生物量曲线地点均位于丰度曲线上方,且互花米草区生物量起始位置与丰度起始位置相隔较2019年远,表明研究区域潮间带底栖生物受到的干扰较小。
2.4 入侵过程
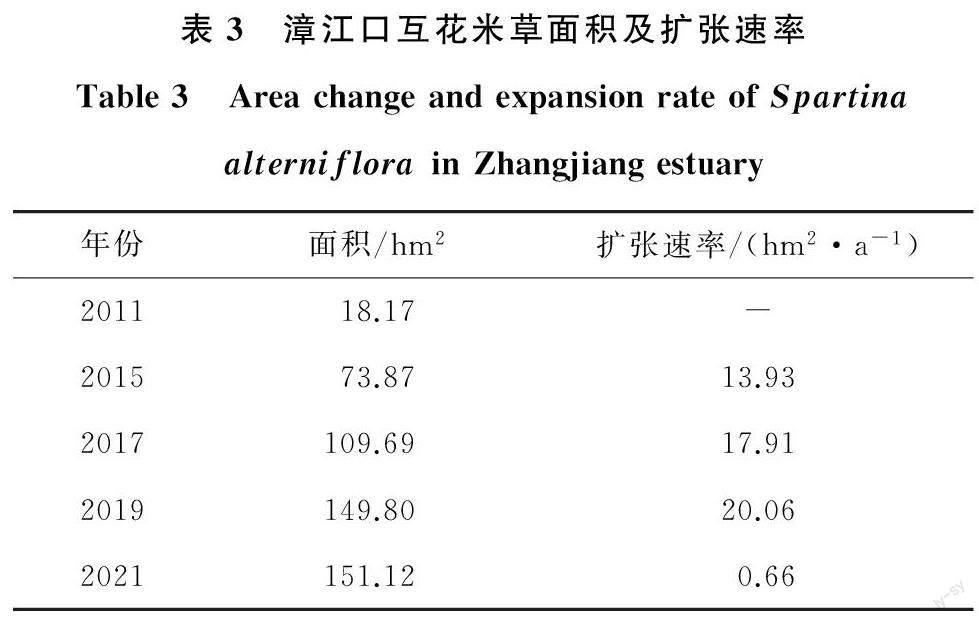
互花米草扩张过程见图7,互花米草面积及扩张速率见表3。
2011年研究区域的大部分区域为光滩,随后互花米草保持较高的扩张速率迅速占领光滩,2021年互花米草的面积是2011年的8.3倍,其扩张方式以外部隔离扩张[17]为主,破碎化程度高。李屹等[18]研究表明,互花米草从2002 年左右出现在云霄红树林保护区之后,经历“快速扩张-减缓扩张-快速扩张”的过程,2010—2014年的面积年平均增长为4.57hm2/a。而本研究表明到2015年,互花米草扩张速率已达13.93hm2/a,2011—2021年扩张速率为12.09hm2/a,可见漳江口互花米草在此期间保持较高的速率扩张,其中2019年互花米草的扩张速率达到最高值20.06hm2/a,但2021年互花米草面积与2019 年相差不大,扩张速率仅为0.66hm2/a,这与漳江口近年开展的互花米草清理整治有关。根据漳江口红树林保护区管理局统计,自2019年以来漳江口红树林保护区陆续开展互花米草除治,除治面积约17hm2,主要位于漳江口保护区竹塔村后港区域滩涂。
3 讨论
3.1 漳江口不同植物生境沉积物、潮间带底栖生物差异
漳江口研究区域互花米草区沉积物中总氮、硫化物和有机碳含量高于光滩,互花米草区沉积物中全盐含量低于光滩。漳江口研究区域潮间带底栖生物种类数、栖息密度由高到低依次为红树林区、光滩、互花米草区,生物量由高到低依次为互花米草区、光滩、红树林区,多样性指数和丰度指数由高到低依次为红树林区、互花米草区、光滩。根据丰度生物量比较法,2020年互花米草区的潮间带底栖生物群落结构较2019年稳定。
3.2 互花米草与红树林的竞争关系
2019年、2020年漳江口互花米草的平均高度为1.27~1.43m,高潮区的株高低于中潮区,这可能与漳江口高潮带生长的互花米草与红树林生长区域相邻、互相竞争有关。林清贤[19]在2001年调查漳江口红树林时发现该区域红树林主要分布于中潮带,红树植物主要为秋茄、桐花树和白骨壤,高度为1.5~6.0m 不等;主要红树植物中秋茄最高,在核心区高度可达6m,桐花树2~3m,白骨壤1.5~3m,红树林林相郁闭度高。可见漳江口红树植物平均株高略高于同潮区互花米草的平均株高,因此在密集的红树林区,互花米草没有竞争优势,无法形成遮阴效应,无法向红树林区域扩张,更倾向于向光滩扩张,类似于向外扩张的“先锋植物”,即促淤的同时降低盐度、增加有机质,为红树林的进一步扩张提供更好的立地条件。与此同时,互花米草区中的底栖生物量高于红树林区,底栖生物密度略低于红树林区,其对底栖生物多样性的干扰,与红树林相比并不明显。但在红树林生长区即红树林幼苗较多的区域,互花米草则存在较强的竞争优势,通过抑制红树林幼苗生长来抑制红树林的扩张。林清贤[19]2001年调查时红树林群落外为泥质滩涂,滩涂分布的其他植物有卡开芦、短叶茳芏和铺地黍等。根据2015年卫星遥感影像,红树林外光滩已逐渐被互花米草占据。据了解,本地盐沼植被目前仅大洲岛上有局部发现,其他本地盐沼植被已很少被发现,这也必然会对红树林生长区产生更大威胁。如何更好地利用互花米草的“先锋优势”为红树林生长服务,可在今后做进一步的研究。
3.3 互花米草入侵过程
互花米草入侵过程分析结果表明,2011—2019年漳江口互花米草保持较高的速率扩张,由于互花米草清理整治,2020年互花米草扩张进入稳定期。Luiting等[20]和洪荣标等[21]的研究结果表明,互花米草区大型底栖动物的栖息密度、种类多样性、丰度比相邻的普通滩涂低。本研究中2019年(快速擴张期)互花米草潮间底栖生物数据支持上述论述。仇乐等[22]认为新生互花米草生境大型底栖动物的物种数和丰富度较高,但随着时间的推移及互花米草的生长,互花米草生境中的大型底栖动物的物种数及多样性都会下降。本研究认为互花米草盐沼在不同的发育时期和不同的扩展时期,大型底栖动物群落的组成、物种丰富度和多样性指数等均发生变化。因此,研究互花米草入侵对滨海湿地大型底栖动物群落的影响,除调查方法及时空因素外,还应考虑样地互花米草入侵时间的长短[21-22],以及样地互花米草入侵的不同扩张时期。
