
降香黄檀根系性状对石灰岩石砾的适应响应
2023-06-20 12:27:38覃桂丽玉舒中
西南林业大学学报 2023年3期
覃桂丽 玉舒中
(广西大学林学院,广西 南宁 530004)
根系是植物摄取、输导与贮存土壤水分和养分的关键功能器官[1-2],也是最先感受到土壤逆境胁迫的器官。不同环境条件下,同种植物(根系)形态、解剖结构常表现出明显的趋异现象[3]。异质土壤生境中,植物会往资源丰富区域的根系分配更多来促进该区域根的生长以便提高资源的获取[4]。植物的根系形态与结构是指根系在生长介质中的空间构型分布、形态、解剖结构等,反映植物获取土壤资源的方式与能力[5]。随着全球气候变化加剧,高温、干旱等极端事件发生频率不断增加,良好的根系形态与结构有利于植物在逆境胁迫中抵抗力,所以提高根系形态稳定性和资源利用效率是提高植物成活率的关键因子。因此,植物根系形态与结构在生态学上研究的重要性不言而喻,目前已成为国内外农林领域关注的热点[6]。广西、云南和贵州是喀斯特石漠化程度最为严重的地区。该喀斯特石漠化区域异质性强,地高水低、石多土少、土薄地漏、保水性差,季节性与结构性干旱及养分匮乏[7],是其植被恢复困难的主要制约因子。石漠化治理所选用的植物需要具有很强的适应性。喀斯特石漠化生境中的适生植物根据当地土壤条件进化出了不同的适应性机制,确定这些植物适应机制,特别是深根系的高大乔灌木,对喀斯特石漠化治理有着非常重要的作用。
降香黄檀(Dalbergia odorifera),是一种适生于喀斯特石漠化地区半落叶高大木本植物,是研究根系性状对石漠化环境适应性的理想树种。但目前关于降香黄檀根系适应贫瘠石漠化环境的机理少有研究。刘震[8]对降香黄檀幼苗在不同石砾含量土壤的适应性硏究中表明,降香黄檀根冠比、根瘤数会随着土壤石砾含量的增加而逐渐増大,在石砾含量达到50%时,根表面积和体积均达到最大且根尖在与石砾接触部分出现膨大现象。倪周游[9]以分层和扇形柱2 种类型模拟石漠化,发现降香黄檀根系具有很好的生理生态适应能力。过高或过低的表土层厚度都会增加根冠比,从而将生物量更多的向三级根分配,以及增加根表面积以及根尖数为根系更好的吸收养分,并通过改变根系生物量分配和增加根系中细根的比例适应石砾土壤环境。关于根系构型与石漠化的研究有黄同丽等[10]和苏樑等[11]研究的喀斯特植被均通过延长根系连接长度以获得更多的水分和养分,同时拓展生长空间,增强固持土壤的能力,防止水土流失等。目前研究喀斯特地区植被根系生态学特征还存在许多问题,仍需继续深入的探索。在今后的研究中,应加强喀斯特地区根系生态学研究,进行植被生境室内模拟实验,构建和完善根系原位收集方法以及加强根系生态学在多重条件下的探究,如时空尺度、植被演替、氮沉降、碳循环等[12]。本研究采用全根和两室裂根系统研究降香黄檀根系性状对石灰岩石砾基质和局部石灰岩石砾基质的适应策略,探讨降香黄檀根系与地上部分性状之间的协调关系,为喀斯特石山地区造林推广和生态治理提供参考依据。
1 材料与方法
1.1 试验材料
本试验在广西南宁广西大学林学院教学实验基地(22°48′N,108°22′E)进行。典型亚热带季风气候,年均降雨量达1 300 mm,年均气温21.6 ℃左右,7—8 月份为最高温[13]。栽培基质黑褐色石灰土和石灰岩碎石采自广西壮族自治区石漠化地区百色市凌雷村(23°33.578′N,106°50.315′E),石灰岩碎石颗粒大小为0.5 cm3,基本理化性质见表1。

表 1 2 种基质的理化性质Table 1 Physicochemical properties of 2 substrates
1.2 试验设计
试验于2021 年3—8 月进行。选择采自同一棵树长势较一致的1 年生实生苗,苗高为(42 ±0.7)cm,地径为(3.6 ± 0.9)mm,去除主根后栽种于图1 所示的全根和两室裂根系统的栽培容器中。两室裂根系统中,按降香黄檀幼苗根数平均分为2 部分[14];同时采用有机玻璃阻断黑褐色石灰土和石砾基质中根系之间养分、水分等物质的交流。每个栽培容器栽植1 株,每5 株为1 组,每个处理4 组,合计栽植60 株。各处理间温度、光照、水分等环境因子保持一致,定期除草,对苗木不做任何施肥处理。栽培120 d 后测定各处理的生物量及根系形态解剖参数。
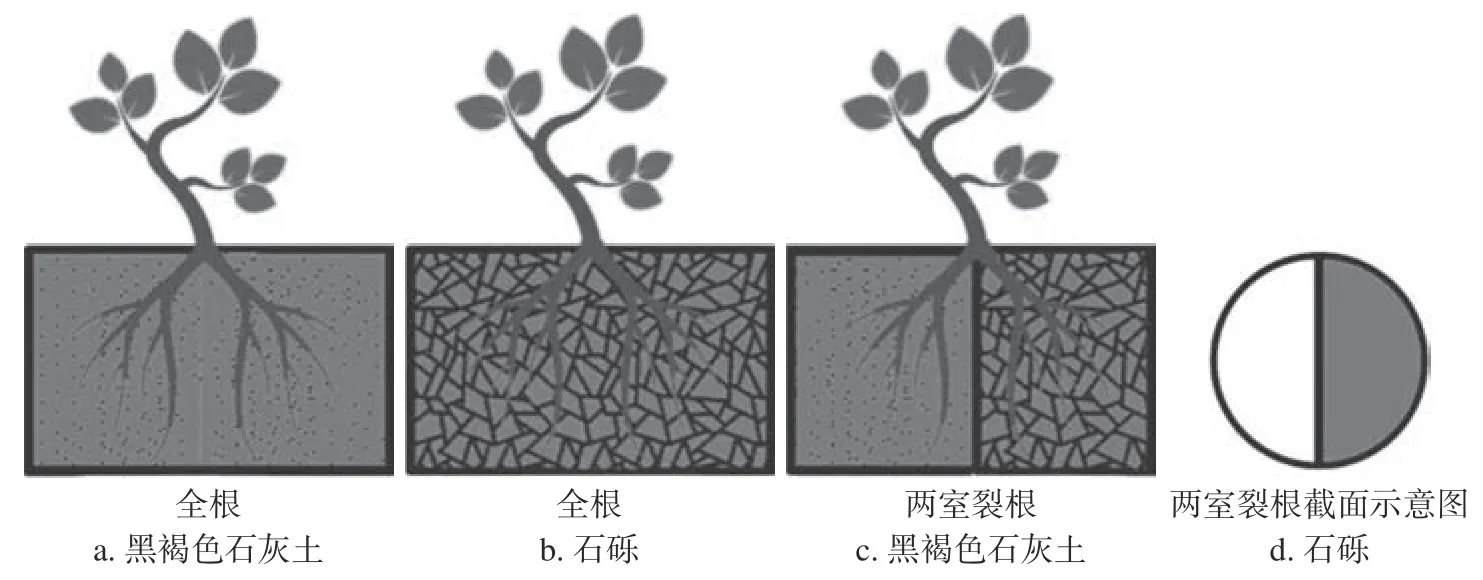
图 1 全根和两室裂根系统示意图Fig. 1 Schematic diagram of whole root and 2 compartment split root system
1.3 测定指标及方法
1.3.1 降香黄檀生长指标及生物量的测定
苗高、地径、高径比参考潘陆荣等[15]的方法。用皮尺(精确到0.1 cm)测定幼苗株高,用数显游标卡尺(精确到0.01 mm)测定地径,每株重复测定3 次,取其平均值,采用烘干称质量法[16]测定生物量。并计算高径比、根冠比和苗木质量指数[17]。
1.3.2 降香黄檀根系形态和根尖解剖结构测定
根系形态和结构参考根系形态和苗木生长指标[18-19]。每个处理随机选取5 株,使用 EXPRESSION 1 680 根系扫描仪(爱普生,日本)测定各项根系指标;每个处理选取3 株降香黄檀幼苗破坏取样,测定其根系总根长、总根表面积、根体积、根尖数量、分枝数量等根系指标,并计算出比根长、比根表面积、分枝密度和根组织密度。
根尖解剖结构参考《植物显微技术》[20]。每个处理采集多个完整根尖,用冰盒保存带回实验室进行。首先用去离子水后切取根尖(0~0.5 )cm,迅速固定于FAA 中,抽气,24 h 后进行一系列梯度酒精脱水,包埋,然后进行横、纵切,番红固绿染色,ICC50W 无线显微镜成像系统(徕卡,德国)镜检照相。每个处理选取 5 个根段作为重复,每个重复制 3 个样片,每个样片观察 2 个视野。利用软件imagej 2018 进行测定根尖横切面直径、基本分生组织厚度、形成层直径、根冠长度。并计算原形成层直径与根尖横切面直径的比值。
1.4 分析方法
利 用Microsoft Excel 2003 整 理 汇 总,运用SPSS 24.0 进行分析,以 Duncan 法多重比较,使用Origin 8.5 软件制图,使用PhotoShop 进行图片排版。
2 结果与分析
2.1 降香黄檀幼苗根系形态对石灰岩碎石基质的响应
由表2 可知,在相同系统下,总根长、根平均直径、分枝数、根尖数、分枝密度差异显著(P<0.05)。全根系统中,黑褐色石灰土基质生长的植物根系总根长、根体积、根平均直径、根尖数和分枝数均显著大于生长在石砾基质中的根系(P<0.05)。在不同系统中,全根系统中的黑褐色石灰土基质下植物的总根长、根平均直径、根尖数、分枝数均显著大于两室裂根系统下(P<0.05)。而全根系统下,石砾基质中生长的植物总根长、总根表面积和根体积显著低于两室裂根系统(P<0.05),其总根长、总表面积、总体积分别比下降了86.24%、76.45%、72.46%。
由表3 可知,在相同系统下,与生长在石砾基质中的植物相比,生长在黑褐色石灰土的比根长、比表面积显著增大(P<0.05)。在不同系统下,两室裂根系统中植物的比根表面积显著高于生长在石砾基质植物,而根分枝密度则显著大于生长在黑褐色石灰土基质中的植物(P<0.05)。两室裂根系统通过增加根系比表面积,增大根分枝密度来适应石灰岩石砾的胁迫环境。
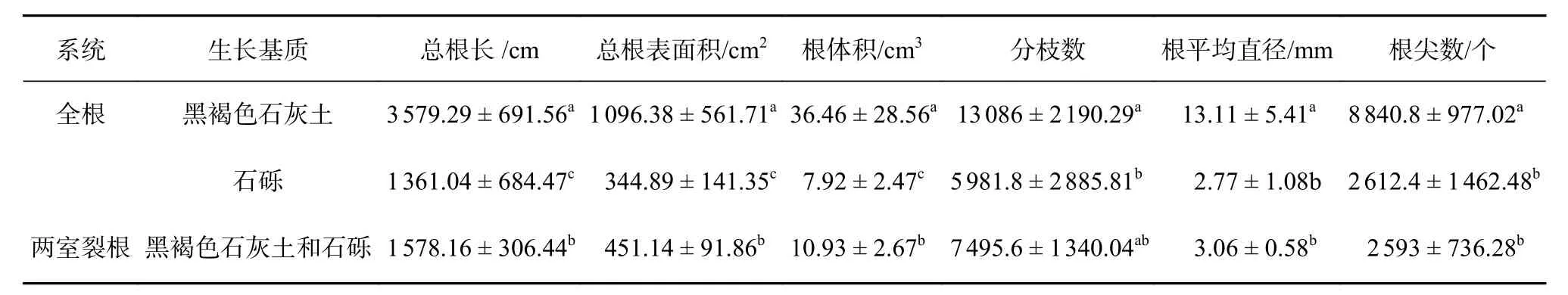
表 2 降香黄檀幼苗根系形态对石灰岩碎石基质的响应Table 2 Response of root morphology of D. odoriferaseedlings to limestone gravel media
2.2 降香黄檀幼苗根尖解剖结构对石灰岩碎石基质的响应
由图2~3 可知,降香黄檀根尖从外到内分别为根冠、原表皮、基本分生组织和原形成层。从形态学下端到上端分别是根冠(静止中心)分生区、伸长区等。从根尖的横切面看(图2),两室裂根系统中植物根尖的原表皮细胞比全根系统中的更大,各个系统中的原生韧皮部均有3 ~ 4 层筛管且原生木质部均为五原型。其中,全根系统的石砾基质和两室裂根系统的黑褐色石灰土基质中植物的原生木质部有逐步分化为六原型的趋势,但石砾基质中根尖细胞较黑褐色石灰土基质中分化的程度更明显,出现了空腔,液泡化程度强,且石砾基质中根尖的后生木质部扩的口径和细胞更小。从根的纵切面来看(图3),全根系统中根冠呈圆锥型,两室裂根系统则为锥子型,且全根系统的根尖原形成层的紫红色着色深度较两室裂根系统更深,说明其原生质的浓度大,细胞分裂旺盛。一直到伸长区,细胞分裂结束,出现液泡,颜色减淡。与其余各基质比,全根系统的石砾基质中生长的植物根尖细胞分化比较早且分化迅速。说明在石砾基质的环境中,根尖在不停的分化、伸长。
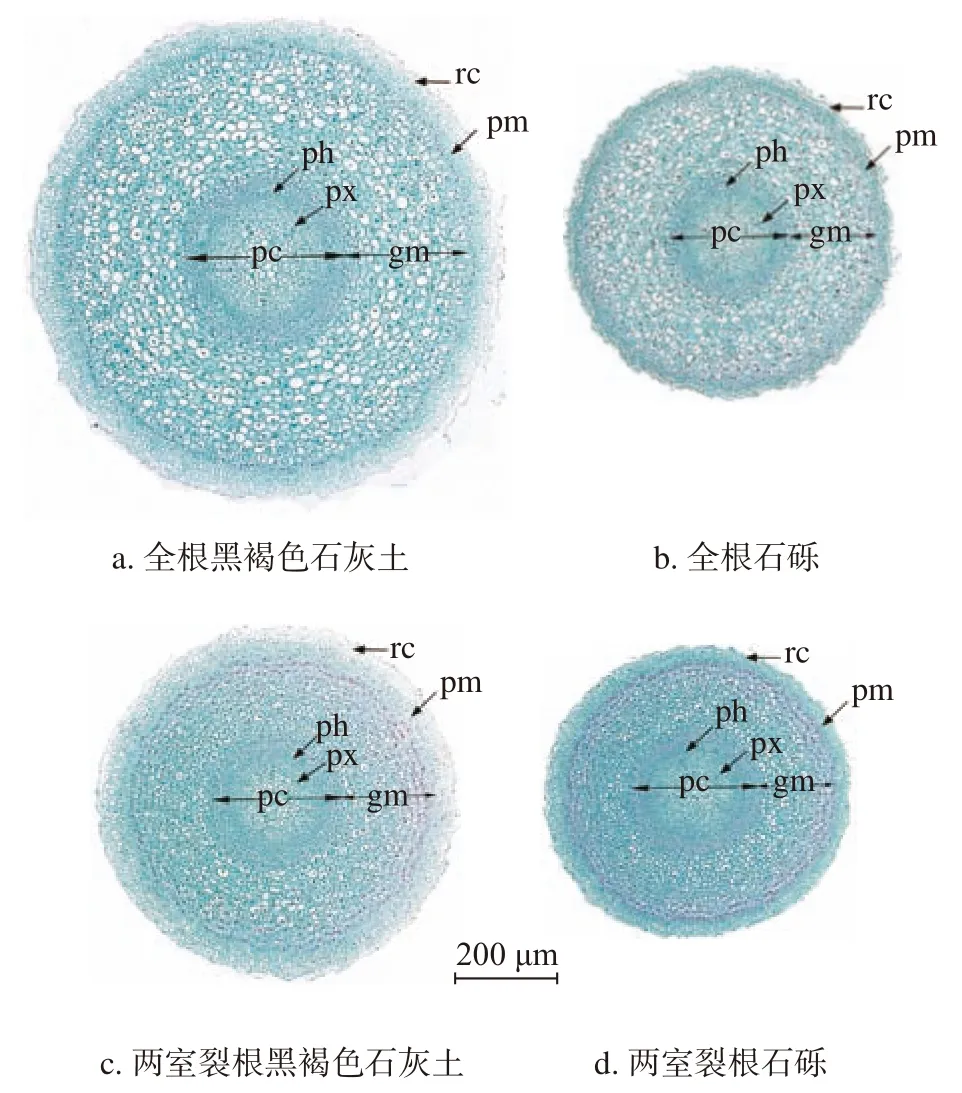
图 2 降香黄檀幼苗根尖的横切面图(4×,距根冠大约5 mm)Fig. 2 Cross section of root tip of D. odoriferaseedlings(Multiple 4×, about 5 mm from root crown)
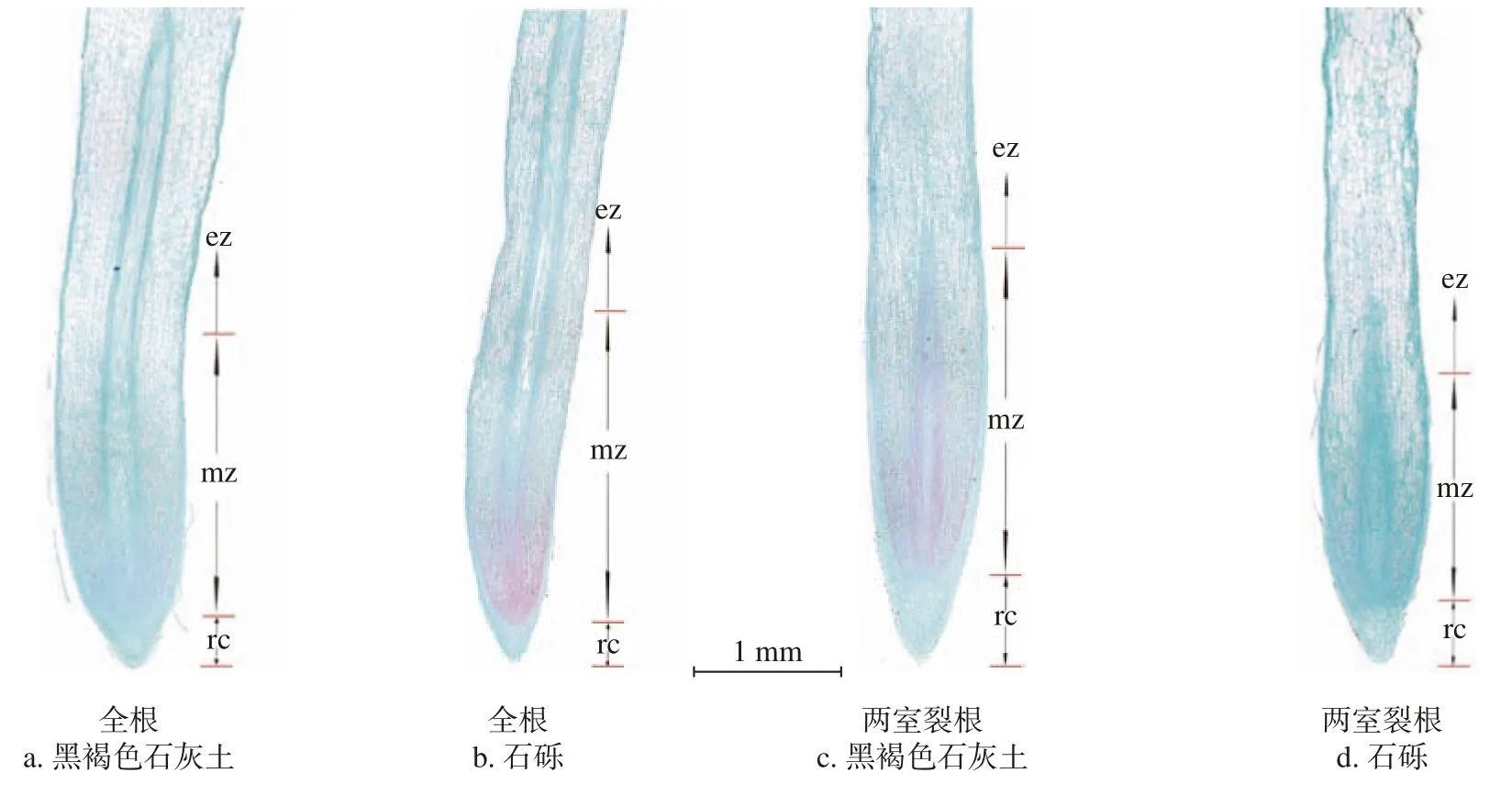
图 3 降香黄檀幼苗根尖的纵切面图(10×)Fig. 3 Longitudinal section of root tip of D. odoriferaseedlings(Multiple 1×)
由表4 可知,在相同系统中,降香黄檀根尖形态解剖结构指标差异显著(P<0.05)。全根系统中,黑褐色石灰土基质下生长的植物根尖剖面直径、基本分生组织厚度、原形成层直径显著大于石砾基质中生长的植物(P<0.05),且石砾基质中植物根尖直径、基本分生组织厚度、原形成层直径降幅最大,分别下降了38.51%、45.17%、39.08%。在两室裂根系统中,黑褐色石灰土基质中植物根冠细胞长度显著高于生长于石砾基质中的根尖大小(P<0.05)。其余指标中,根尖直径和基本分生组织厚度分别减小10.95%、18.47%,原形成层直径增加0.5%,原形成层直径与根尖直径比显著增加9.76%。在相同基质下,黑褐色石灰土基质下的植物根尖剖面直径、基本分生组织厚度、原形成层直径为全根系统显著大于两室裂根系统,而根冠细胞长度则相反,两室裂根系统显著高于全根系统(P<0.05)。生长在石砾基质中的根尖原形成直径为全根系统显著低于两室裂根系统(P<0.05)。由此可知,石砾基质中生长的植物根尖主要通过减小根尖直径和原形成层直径,适应石灰岩石砾的胁迫环境;而两室裂根系统下,则通过增加根冠厚度来保护根尖细胞。
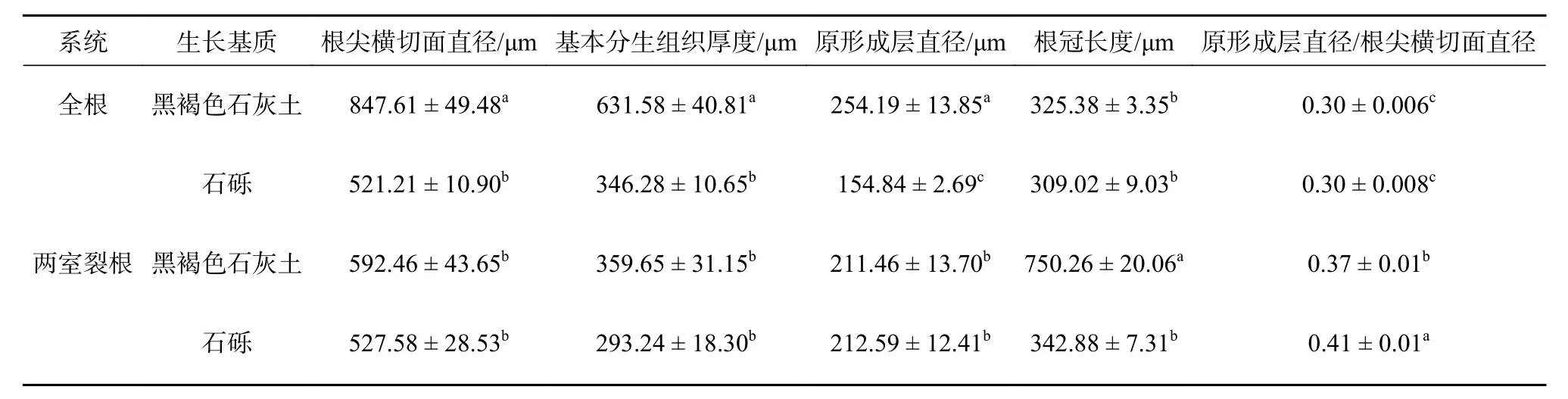
表 4 降香黄檀幼苗根尖解剖结构对石灰岩碎石基质的响应Table 4 Response of root tip anatomical structure of D. odoriferaseedlings to limestone gravel media
2.3 降香黄檀幼苗生长及生物量配对石灰岩碎石基质的响应
由表5 可知,不同系统中降香黄檀幼苗的苗高、地径相对增长率,地下和地上干质量、根冠比指标差异显著(P<0.05)。相比于生长在全根系统的黑褐色石灰土基质下的植物,生长于全根系统的石砾基质和两室裂根系统中苗高和地径相对增长率、地上和地下干质量显著减小(P<0.05),高径比无显著差异,根冠比显著增大(P<0.05)。其中植物在全根系统的石砾基质中,地下和地上生物量分别降低了29.21%、63.33%,降幅最大,根冠比增加了79.17%,增幅最大;另外,生长于全根系统的黑褐色石灰土基质中的苗木质量指数最大达2.49,石砾基质中最小为1.15。
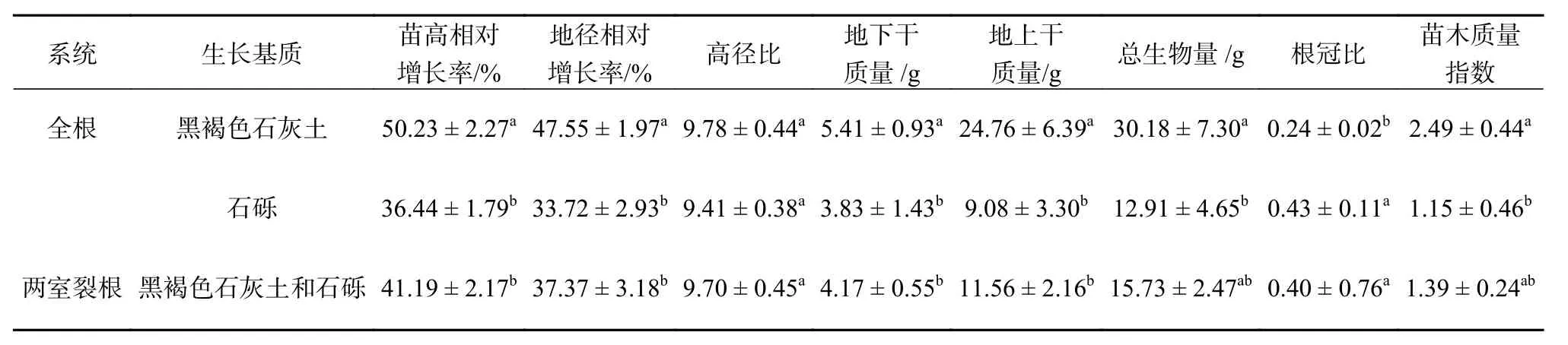
表 5 降香黄檀幼苗生长及生物量配对石灰岩碎石基质的响应Table 5 Response of growth and biomass of D. odoriferaseedlings to limestone gravel media
3 结论与讨论
3.1 根系性状对石灰岩石砾的适应性变化
近些年,根系研究在许多领域(如形态学、生理学、根系架构学、生物力学和解剖学等)取得了长足的发展[21],从单一的根经济学谱进入了一个多维的根系经济学谱空间[22]。但与叶经济学谱相比,目前对根性状与功能关系的认识还相当有限的。因此,为了更好地了解降香黄檀根系变异对土壤环境变化的对策,本研究的分析聚焦于8 月份降香黄檀最活跃时期内的几个关键根系(根尖)性状指标(根平均直径、根组织密度、根分枝数、根分枝密度、根横切直径、原形成层直径)变异。
根系的大小、形态结构以及分布范围等直接决定着植物对土壤资源的吸收和利用能力[23-24]。在植物生长阶段的环境因子对根系功能性状产生重要影响,处于不同环境资源的植物根系外部形态会发生变化,导致根系同一性状的特征有所差异,从而表现出不同的生长方式和适应策略[25-26]。其中根长、表面积、直径以及结合生物量的综合评价如比根长、比表面积、根组织密度和分枝密度是衡量根生长的重要指标[27-28]。本研究中,与生长在全根系统下黑褐色石灰土基质中的植物根系相比,两室裂根系统根系根长、比根表面积逐渐减小,意味着降香黄檀分根后对养分和水分等资源有着很强的竞争力能保存较多的稀缺营养且细根较多,从而,有利于降香黄檀在严酷的生境中生存[29]。同时,这一现象说明,在不同石灰岩石砾胁迫下,降香黄檀并不是被动忍受逆境胁迫,而是主动调节其根系生长以提高对水分和养分的吸收能力,减缓逆境伤害。根组织密度是描述资源获取和保存的另一个关键特征,被认为与根寿命显著相关,组织密度越大寿命越长[30-31]。一项对13 种热带树种的研究发现,根系呼吸随着根组织密度的增加[32]。植物生长在不同系统和基质中的根组织密度差异不大,但生长石砾基质的根组织密度相对而言较大,而黑褐色石灰土基质中较小。因此推定,降香黄檀根组织密度与整株受迫程度有关且根系所在的区域资源受限越大,该区域根组织密度就越大。验证了高根组织密度是反映对贫瘠土壤适应的最一致的根系性状的结论[33]。另外,随着石灰岩石砾胁迫的增加,降香黄檀的生长受到了抑制,分枝数也随之减少。分枝密度的变化是生长在全根系统下黑褐色石灰土基质中的植物最低,两室裂根系统中最高。即整株根系处于石灰岩石砾的局部胁迫(石砾基质)时,根分枝密度有所增大,转变为增加勘探土壤体积的对策。说明区域越贫瘠分枝密度越高,根系收到胁迫时,根系趋同采取增大分枝密度提高勘探精度提高获取的对策,幼苗根系具有向深处延伸生长的趋势。
3.2 根尖对石灰岩石砾的适应性变化
在不同逆境条件下,植物对环境的响应是有所不同的,除了地上部分外观有所显现之外,植物根系的解剖结构对环境的响应会更加明显[34]。本研究中降香黄檀的根尖细胞仍在进行不断的分裂、生长和分化。降香黄檀在不同系统和基质下的根尖解剖结构表现为植物生长在全根系统下黑褐色石灰土基质中的根尖横切面直径、基本分生组织厚度、原形成层直径显著大于处理石砾基质和两室裂根系统中根尖的生长。其中生长在全根系统下石砾基质中的原形成层直径小于两室裂根系统。表明在整体和局部石灰岩石砾胁迫下,主要通过降低根尖基本分生组织厚度和原形成层直径来提升根系的吸收效率及养分运输能力[35-36]。从降香黄檀根尖的纵切面来看,两室裂根系统较全根系统的根冠两侧的角度更小,变为尖锥弧形状,分生区和伸长区的宽度较窄,细胞间隙减少,根冠细胞层数增多。同时在石砾基质下,植物根尖出现弯曲。说明石砾基质中的根尖比黑褐色石灰土基质受到的阻力更大,根冠随根尖的增细变长,由椭圆弧形变为尖锥弧形,根尖的角度变得更加倾斜,且推动力更强。这也可能是由于在石砾的挤压和摩擦下以及石砾孔隙大的原因,使得根尖利于向前延伸和发展而逐渐形成的。可知,在石灰岩石砾基质和局部石灰岩石砾基质的胁迫时,根尖的形状和大小均发生不同的变化,主要是通过减小根直径和增加细根的数量,从而加大与土壤接触面积来适应。
3.3 降香黄檀幼苗生长及生物量分配对石灰岩碎石基质的响应
植物的地下部分(根系)的生长状况很大程度上决定了地上部分的生长。当植株生长受到养分等环境因子限制时,会通过调节各器官的光合产物分配比例来应对环境胁迫。本研究表明不同系统对降香黄檀幼苗的生长指标和生物量分配均具有显著影响。相较于植物生长在全根系统下黑褐色石灰土基质中,生长在全根系统下石砾基质和两室裂根系统的降香黄檀苗高和地径增长率均表现出下降趋势且在石砾基质下降的幅度最大。因此,生长在全根系统下的黑褐色石灰土基质中的降香黄檀幼苗状况最佳,而局部胁迫(两室裂根系统)的苗木生长质量比整体胁迫(全根系统下的石砾基质)中更佳。主是由于在两室裂根系统中降香黄檀根系生长在局部石灰岩石砾基质中,两室的营养物质不同,在营养的吸收上会通过吸收的营养物质多的一侧来弥补营养物质不足的一侧,以保证其存活[37]。本研究中,整株植物生长于黑褐色石灰土基质中的地上、地下生物量含量最多,其次是局部胁迫的两室裂根系统的生物量高于整体胁迫下的石砾基质中生长的植物且两者相差较大。表明降香黄檀通过改变根系(地下部分)生物量的分配来适应复杂的生长空间,同时,在石灰岩石砾基质的胁迫下,地上和地下部分的生长是协同的关系。这符合功能平衡假说,植物通过改变地下生物量的分配来获取限制植物生长的环境资源[38],当受到胁迫时,植物通过增加地下生物量的分配以保证水分的供应。苗高和地径的平衡关系通常用高茎比来体现,表征着植物抗性的强弱。高径比越小,表明植物越健壮,抗性越强。各系统之间降香黄檀的高径比无显著差异,说明在石灰岩石砾的基质(全根系统下石砾基质)和局部石灰岩石砾(两室裂根系统)胁迫下,对降香黄檀地上部分各器官之间的协调性影响不大;其次,植物地下部分生长和地上部分生长之间的平衡关系通常用根冠比来表征。根冠比越大,越有利于植物在严苛的不利条件下保持养分与水分平衡及存活。随着胁迫增加,降香黄檀的根冠比显著提高,降香黄檀在全根系统下石砾基质和两室裂根系统中的根冠比分别是生长在全根系统下黑褐色石灰土基质中的1.79 倍与1.67 倍。由此可知,降香黄檀幼苗为了生存,通过降低苗木的高度而矮化植株并将更多生物量分配到地下,增加地下生物量的分配模式来达到增强对不利环境适应性。该结论和Mokany等[39]和龙毅等[40]的研究结果一致。
综上所述,地下部分(根系、根尖)和地上部分共同影响着降香黄檀幼苗各器官对水分和养分的吸收利用。局部石灰岩石砾胁迫对降香黄檀幼苗的生长和物质分配具有促进效应。同时,对降香黄檀根系的生长(总根长、根表面积、分枝数)、分枝密度和根冠长度具有不同程度的促进作用,而整体石灰岩石砾胁迫则具有抑制作用。降香黄檀幼苗生物量分配对石灰岩石砾的响应表现为:局部石灰岩石砾胁迫增加促使地下生物量和根冠比显著增加,而整体胁迫则使地下生物量减少和根冠比增加,植物以此来调整各器官物质分配进而优化自身对有限资源的获取。石灰岩石砾胁迫还影响着降香黄檀根尖解剖结构的变化,随着胁迫程度增强,根尖横切面直径和基本分生组织厚度随之减小。
猜你喜欢
林业与生态(2022年5期)2022-05-23 01:16:51
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
天然产物研究与开发(2018年5期)2018-06-13 03:23:40
现代农业科技(2017年16期)2017-09-22 14:47:46
山东林业科技(2017年1期)2017-06-29 07:53:52
广东药科大学学报(2016年1期)2016-04-11 05:28:57
广西林业科学(2016年2期)2016-03-20 05:53:32
中国继续医学教育(2015年4期)2016-01-07 07:38:13
江苏农业科学(2014年1期)2014-07-18 21:21:34
