
扎龙湿地藻类优势种生态位及其种间联结性动态分析
2023-06-14 08:17:14范亚文杨应增
生态学报 2023年10期
李 雪,夏 伟,范亚文,杨应增
1 茅台学院,遵义 564507
2 哈尔滨师范大学生命科学与技术学院,黑龙江省水生生物多样性研究重点实验室,哈尔滨 150025
湿地生态系统,是物质循环与能量转换的重要生态载体,对于区域气候调节、生态环境具有重要作用[1]。受气候变暖和城市化进程加快等因素影响,湿地自然生态问题日益突出,湿地功能和可持续发展面临重大威胁[2]。藻类是水体中的初级生产者,也是水域生态系统的重要组成部分及湿地环境变化的重要标志[3]。藻类的群落结构的变化与人类活动和自然环境的季节性变化密切相关[4—5]。目前,对于湿地藻类的研究主要集中在环境因子对生物群落结构影响方面,而基于生态位和群落演替等方面则较少。
生态位研究在了解群落结构及功能、物种多样性、群落动态演替和种群进化等方面有重要的参考作用[6]。生态位理论是通过生态位宽度和重叠程度来揭示在特定环境中物种的竞争能力,从而解释其资源利用能力和分布情况[7—8]。物种间关联研究则是探究物种之间分布格局的关联性[9]。结合生态位理论和物种间关联研究,对探讨群落结构和功能、群落种间关系、动态演替等有重要作用[10]。目前,生态位的研究已应用于陆地动植物生态学[11—15]、水生动物生态学领域[16—19]。该方面,浮游植物生态位研究多限于空间生态位研究[20—21],湖泊[22]、水库[23]以及河流[24],利用藻类生态位理论预测水华暴发的预警系统已取得较好效果[25],但对典型内陆性高寒湿地浮游植物时空生态位分析则少有报道。
本研究以黑龙江省扎龙国家级自然保护区为对象,通过其藻类群落调查,分析生态位测度,结合MFG藻类功能群划分[26],对其藻类优势种开展分析,评估其优势种的生态位和相互关系,为湿地水生态系统的保护提供理论参考。
1 材料和方法
1.1 区域概况及采样点布设
扎龙国家级自然保护区(46°52′—47°32′N,123°47′—124°37′E)位于黑龙江省西部、齐齐哈尔市东南部松嫩平原、乌裕尔河下游湖沼苇草地带,总面积2100 km2,是以芦苇沼泽为主的内陆性湿地和水域生态系统[27]。该研究于2012年的5月(春季)、7月(夏季)、9月(秋季)对扎龙湿地进行藻类植物进行调查和样品的采集。取样点分布见图1。

图1 扎龙湿地取样点示意图Fig.1 Sampling sites schematic diagram in Zhalong wetlandS:取样点Sampling point;1—9:取样点编号
根据生态环境的分布特点,将研究区域划分为4个采区域,根据区域特点设置9个取样点:沼泽开阔型水体(S1),位于保护区上游区域,属于芦苇沼泽中大型开放水域,水面开阔;小型封闭水体(S2),丹顶鹤养殖区域附近小水域,水域面积小,水体封闭,且受丹顶鹤粪便等影响较大,水体有机污染较为严重;湖泊边缘半封闭水体(S3—S7),芦苇丛生,水位较浅,流速较缓;湖泊型水体(S8—S9),沼泽湿地主要汇水区域,水面开阔,水位较深,芦苇分布较少。
1.2 样品的采集与鉴定
藻类样品采集采用有机玻璃采水器,在表层0.5m处采集1L水样,现场加入15mL鲁格试剂固定保存。带回实验室后,静置沉淀48h后,基于虹吸原理,去除上清液,将其浓缩至30mL,取0.1mL浓缩样品于浮游生物计数框中,在MoticBA200显微镜下进行种类鉴定并计数。藻类种类鉴定参照《中国淡水藻类-系统、分类及生态》及《淡水微型生物图谱》等[28]。
1.3 数据分析
数据分析使用R语言平台中的spaa、vegan等函数包,数据处理和绘图均在Microsoft Office、R语言平台中完成。
1.3.1藻类物种优势度
利用Mcnaughton优势度指数(Yi)确定优势种,计算公式如下:
(1)
式中,ni为样点中第i种浮游植物的个体数,N为样点中所有浮游植物的总数,fi为该种属在各样点中出现的频率;其中Yi≥0.02的藻类确定为优势藻种,Yi>0.1为绝对优势种。
1.3.2生态位特征
采用改良后的Levins公式计算生态位宽度(Bi),公式如下:
(2)
式中,Bi为物种i的生态位宽;度Pij=nij/Ni为物种i在第j个资源状态(即采样点)下的个体数占该种总数的比例;r为采样点数。
采用Pianka指数公式确定生态位重叠指数(Oik),公式如下:
(3)
式中,Oik为生态位重叠指数,重叠值介于0—1;Pij和Pkj分别为物种i和k在第j个资源状态下的个体数占该种所有个体数的比例;r为采样点数。当Oik>0.6 时视为重叠度较高,0.3≤Oik≤0.6 视为重叠度较低,Oik<0.3 视为重叠度低。
1.3.3总体联结性检验
物种间总体的关联性测定采用Schluter的方差比率法(VR),计算公式如下:
物种相对多度的方差
(4)
相对多度频率
(5)
物种数的方差
(6)
样方中物种数的平均数
(7)
方差比率
(8)
统计量W
W=VR×N
(9)

1.3.4种间联结性检验
种间联结性采用χ2统计量定性分析。选取各季节优势种进行种对间联结性及相关性分析。构建种对间的2×2列联表,对a、b、c、d的值进行统计。a为物种A和物种B共同占据的样方数;b为只出现物种A的样方数;c为只出现物种B的样方数;d为物种A和物种B都不出现的样方数;n为所有采样点数。将b和d值加权为1以避免分母值为0,采用Yates的连续校正公式计算卡方值,进行卡方检验,公式为:
(10)
式中,N为取样点数。当ad>bc时,种对间为正联结;当ad
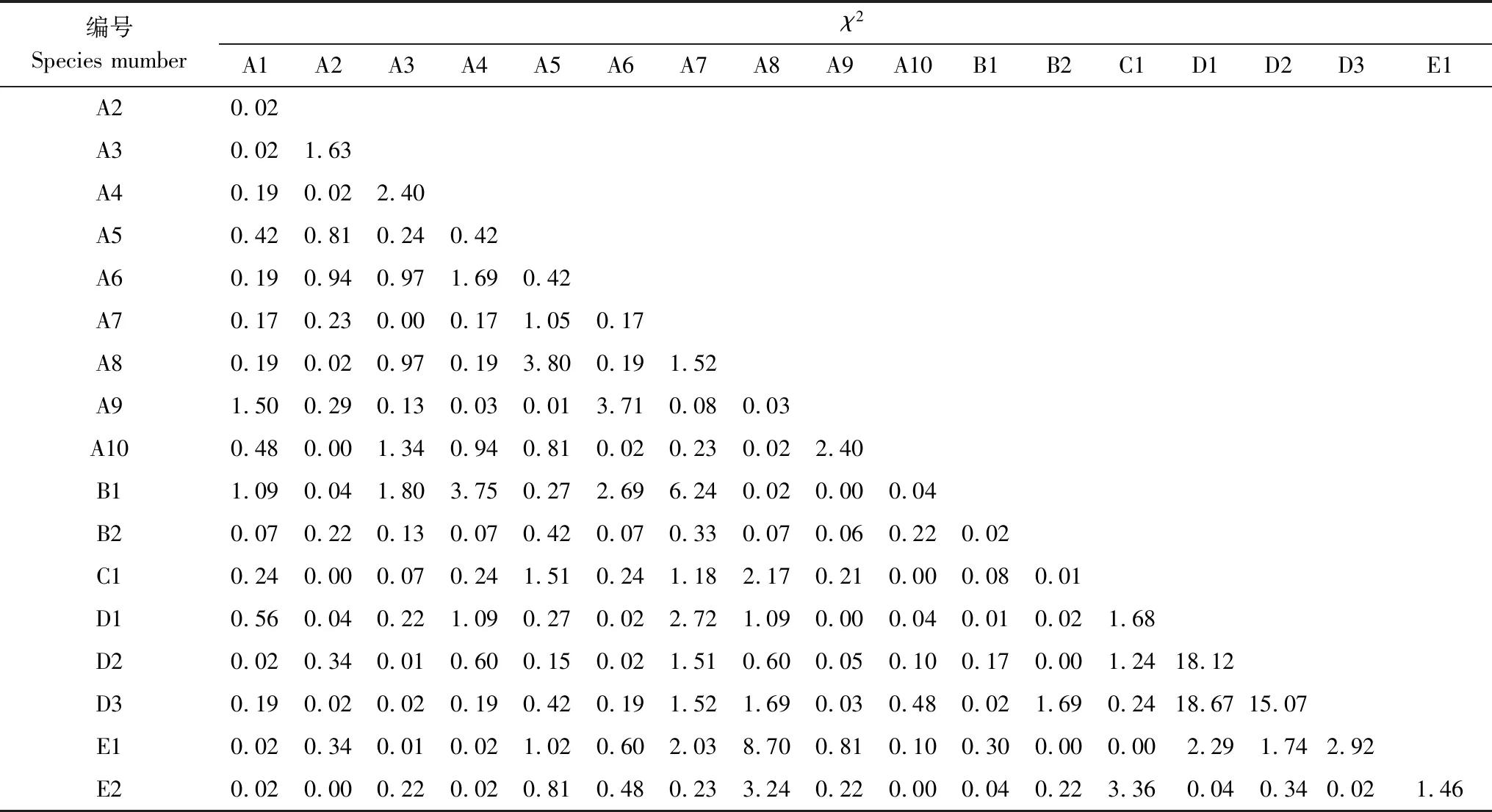
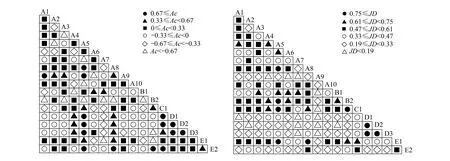
χ2值不能定量区分连结强度。因此,需要结合联结系数(Ac)、Jaccard(JD)等指数对中间联结程度进行进一步分析。若ad≥bc,则Ac= (ad-bc)/[(a+b)(b+d)];若bc>ad且d≥a,则Ac= (ad-bc)/[(a+b)(a+c)];若bc>ad且d 共鉴定出藻类 8门311种。其中绿藻门161种、硅藻门53种、蓝藻门38种、裸藻门36种,四者占总种数的91.77%。其它门类相对较少,黄藻门8种,金藻门6种,甲藻门11种,隐藻门4种,四者仅占总种数的8.23%。不同季节物种丰富度存在一定差异,其中夏季最高达255个分类单元、春季224个分类单元、秋季227个分类单元,见图2。 图2 藻类植物物种组成和密度季节分布Fig.2 The composition of species and seasonal distribution of algae density 全年藻类平均密度为2.27×105个/L,各门藻类种类密度季节性变化明显。如图2所示,春季,藻类各门密度的变化范围为(0.008—10.853)×104个/L,其中金藻门密度占比最大(39.48%),其次是绿藻门和隐藻门(33.51%和13.30%)。夏季,藻类各门密度的变化范围为(0.022—15.085)×104个/L,其中绿藻门密度占比最大(50.74%),其次是硅藻门和隐藻门(18.98% 和12.29%)。秋季,藻类各门密度的变化范围为(0.044—9.767)×104个/L,其中绿藻门密度占比最大(52.38%),其次是硅藻门和隐藻门(19.19%和14.69%)。各门藻类密度的变化趋势为:绿藻、硅藻、蓝藻以及甲藻密度均呈现夏高秋低的分布特征;金藻门仅在春季具有较高密度,夏秋季节密度极低;裸藻密度从春季开始,逐季下降;黄藻全年密度维持在较低水平。 2.2.1丰度和优势度 根据优势度计算结果(Y≥0.02),扎龙湿地藻类优势种5门18种,结果如表1所示。 表1 优势种的密度(D,×104个/L)、优势度(Y)和生态位宽度(B)Table 1 Density (D,×104cells/L),dominance(Y)and niched width(B)of dominant algal species 全年优势种种类数较多,优势度在不同季节差异较大:春季优势种有8种,以金藻门和绿藻门的优势种为主,优势种群显著;夏季和秋季以绿藻门和隐藻门的优势种为主,无绝对优势种。绿藻门的狭形纤维藻和隐藻门的啮蚀隐藻为全年优势种,而部分种属仅在某一季节为优势种属,如金藻门的密集锥囊藻、长锥形锥囊藻在春季为绝对优势种,丰度值最高。 2.2.2优势种的生态位宽度 根据生态位宽度的大小将优势种大致分为广生态位(Bi>0.700)、中生态位(0.500≤Bi≤0.700)和窄生态位(Bi<0.500)三大类群。扎龙湿地藻类优势种生态位宽度值如表1所示,春季优势种的生态位宽度值变化为0.197—0.971,其中属于广生态位的有狭形纤维藻、长锥形锥囊藻和密集锥囊藻,属于中生态位的有卷曲纤维藻、啮蚀隐藻和群聚锥囊藻,属于窄生态位的有镰形纤维藻和旋转囊裸藻。夏季优势种的生态位宽度值变化为0.301—0.504,均为窄生态位种。秋季优势种的生态位宽度值变化为0.155—0.754,其中属于广生态位为啮蚀隐藻,属于中生态位的有四尾栅藻、尖尾蓝隐藻,其它属于窄生态位。 根据不同季节生态位宽度将扎龙湿地藻类优势种分为3类:第一类,在某一季节高生态位宽度和优势度。只在某一个季节成为绝对优势种并具有较高的生态位宽度值,这一类优势种可利用某些季节特定的生态因子(如温度)迅速成为优势种,在当季具有强的竞争力,如金藻门的3个种:长锥形锥囊藻、密集锥囊藻和群聚锥囊藻,春季除小型封闭水体区域外,其它采样区域均为广生态位优势种;第二类,在多个季节均具有较高的生态位宽度值,此类物种受季节因素限制小,环境适应能力较强,具有较广的生态位幅度,但竞争和利用资源的能力一般,能在多个季节成为优势种,优势度值介于0.02—0.1之间,如狭形纤维藻、啮蚀隐藻、尖尾蓝隐藻和旋转囊裸藻,各采样区域均有分布;第三类,在某一季节成为优势种,但生态位宽度和优势度值低于前两类。此类物种只在某一季节成为优势种,对生态因子的依赖性强,竞争能力相对前两类较弱,水体生境的变化易对其产生影响,如绿藻门的珍珠角星鼓藻,只在夏季湖泊型水体区域成为优势种。 2.2.3优势种生态位重叠 生态位重叠是指两个或两个以上生态位相似的物种生活于同一空间时分享或竞争共同资源的现象。春季优势种空间生态位重叠指数的变化范围为0.084—0.893(表2),其中狭形纤维藻和密集锥囊藻生态位重叠值最大,旋转囊裸藻和群聚锥囊藻生态位重叠值最小。夏季优势种空间生态位重叠指数的变化范围为0.089—0.939(表3),其中小形月牙藻和尖尾蓝隐藻生态位重叠值最大,尖尾蓝隐藻和肘状针杆藻生态位重叠值最小。秋季优势种生态位重叠指数变化范围为0.004—0.902(表4),其中尖尾蓝隐藻和啮蚀隐藻生态位重叠值最大,湖生卵囊藻和弯棒杆藻生态位重叠值最小。 表2 春季藻类优势种生态位重叠指数(Oik)Table 2 Niched overlaps(Oik)of dominant algal species in spring 表3 夏季藻类优势种生态位重叠指数(Oik)Table 3 Niched overlaps (Oik)of dominant algal species in summer 表4 秋季藻类优势种生态位重叠指数(Oik)Table 4 Niched overlaps (Oik)of dominant algal species in autumn 优势种的生态位重叠指数大于0.600的组数,春季为12组(42.85%)、夏季为6组(28.57%)、秋季为10组(27.78%),季节间生态位重叠差异较大,总体上春季高于夏秋两季。 表5 扎龙湿地藻类优势种总体关联性Table 5 General interspecific associations of dominant algal species 种间联结指不同种类在时间或空间分布上的相互关联性。单一的检验方法可能会导致结果存在误差,因此以卡方检验为基础,结合联结系数、Jaccard指数进一步检验卡方结果联结性不显著的种对,以达到更客观判断种对间联结关系。扎龙湿地藻类优势种种间联结性检验结果χ2检验结果如表6所示。在171个种对中,4个种对表现出极显著正联结关系(χ2>6.635),分别是在春季成为优势种的3个种对,密集锥囊藻与群聚锥囊藻(18.12)、密集锥囊藻与长锥形锥囊藻(18.67)、长锥形锥囊藻与群聚锥囊藻(15.07),该3个种对在除小型封闭水体区域外的其它采样区域均有分布。此外,在秋季成为优势种的1个种对,弯棒杆藻与针形纤维藻(8.70),该种对主要出现在湖泊型水体区域,其它种对联结关系不显著(χ2<3.841),Jaccard指数的结果也印证了这一结论。 表6 藻类优势种的χ2统计量检验Table 6 χ2 correlation test of the dominant algal species 从图3可以看出171种对中有15组种对间联结程度较高(Ac≥0.67),表现出强的正联结性,占总数的8.77%;0.33≤Ac<0.67的有17对,表现出正联结程度一般,占总数的9.94%;-0.33≤Ac<0.33的有106对,种对间趋于独立,占总数的61.99%;-0.67≤Ac<-0.33的有17对,表现出负联结程度一般,占总数的9.94%;Ac<-0.67的有16对,表现出强的负联结性占总数的9.36%。从Jaccard(JD)结果可以看出JD≥ 0.75的种对有8对,表现出非常紧密的联结性,占比4.68%;JD<0.19的种对有13个。表现出无联结性,占比7.60%。 图3 藻类优势种种间联结系数(AC)和Jaccard指数(JD)半矩阵Fig.3 Semi-matrix of association coefficient (AC) and (JD) or dominant algal species 扎龙湿地春季优势种中,广生态位种为绿藻门的狭形纤维藻、金藻门的长锥形锥囊藻和密集锥囊藻。狭形纤维藻为普生性种类,广泛存在各类水体中,因而对水体指示性不强,而广生态位种金藻种类根据MFG功能群划分解1a/2a(有鞭毛混合营养型金藻门种类大个体/小个体)功能群,一般情况下有利于它们生长繁殖的环境条件主要是温度较低,透明度较大,溶解有机质含量低的水体,并且对温度变化感应灵敏[29]。表明春季水体环境为贫营养的小型浅水水体。夏季与春季优势种差异较大,长锥形锥囊藻和密集锥囊藻夏季不在是优势种,绿藻门、硅藻门小个体优势种增多,且生态位均为窄生态位种。杨文焕等[30]研究表明,窄生态位种出现在特定的时空条件,是敏感型指示种。夏季优势种根据MFG功能群划分为8b/9d(无鞭毛自养型其他藻类单细胞大个体/小个体)功能群,表明此时水体环境处于贫营养混合浅水状态,该状态为更多藻类提供了发展空间。秋季啮蚀隐藻成为唯一的广生态位种,优势种根据MFG功能群划分为2d(有鞭毛混合营养隐藻门小个体)功能群,指示水体秋季状态为中营养的浅水静水水体。监测周期内,扎龙湿地水体环境由春季“贫营养的小型浅水水体”向夏季“贫营养混合浅水体”和秋季“中营养的静水浅水水体”进行季节性转变,这与于洪贤等[31]的结果一致。 春季扎龙湿地优势种重叠值高的种对在3个季节中最多为12对,除与旋转囊裸藻配对的种对外,其它种对间生态位重叠值较高,尤其是出现狭形纤维藻的种对,表明各优势种在空间资源分布上的同域性强,对资源的竞争也最激烈。原因可能是春季随着气温回升和光照时间、强度的增加,适合贫营养混合浅水的单细胞种类,争夺资源的趋势加剧,导致竞争激烈。夏季优势种重叠值与春季相差较大,具有高重叠值的狭形纤维藻、尖尾蓝隐藻种对均为3对、小型月牙藻和珍珠角星鼓藻1对,表明夏季物种对资源的利用共性小,竞争态势相对均衡,种群间趋于稳定发展。原因可能是经历了春季激烈竞争后,一些种类逐渐衰退(如金藻门种类),季节因素影响下,各类资源较为充足,水体环境向混合营养型转变,各优势种占据各自的空间发展,减小了种间竞争。秋季重叠值高的种对比例进一步降低,其中狭形纤维藻已无高重叠值种对,种对间重叠值最高的为啮蚀隐藻和尖尾蓝隐藻种对。可能受秋季温度下降、光照时长和强度降低等因素影响,部分种类逐渐进入衰退状态,竞争能力下降,大量有机质中营养元素的释放,水体开始向有机质丰富的中营养状态转变,为啮蚀隐藻和尖尾蓝隐藻快速发展创造了机会。同时从全年优势种狭形纤维藻与啮蚀隐藻生态位重叠值变化规律看,两者呈现相反的变化趋势。狭形纤维藻与其它优势种呈现春季向秋季逐渐降低的趋势,啮蚀隐藻反之,这可能与水体温度和有机质变化趋势有关,啮蚀隐藻在温度较低的有机质丰富的水体中竞争力更强[32]。优势种生态位重叠值受季节因素影响较大,各季节间存在较大差异。春季优势种间资源的竞争最为突出,以金藻门种类为主,夏季达到相对稳定状态,绿藻门等优势种占据主导,秋季隐藻门竞争开始加剧。 卡方检验结果表明,种间联结性最为显著的是金藻门的3个种对,表现为只在春季成为优势种,且广泛分布于除小型封闭水体区域外的其它采样区域,这与金藻门耐低温、对温度变化敏感以及水质要求清洁密切相关。此外,弯棒杆藻与针形纤维藻种对种间联结性也极为显著,表现为春、夏两季广泛分布于各采样区域,但生物量低,秋季在湖泊型水体区域成为优势种,湖泊区域作为湿地的营养物质的汇集区,秋季较为肥沃的水体为其发展提供了条件。总体而言,171个种对中仅4对呈显著相关,其它167个种对种间联结性不显著,种对种之间趋近于相对独立,这一结果可能与湿地水域宽阔,水流平缓,易形成局部小环境,导致整体关联度下降有关。但卡方检验只能对种间联结性做定性判断,无法区分其大小,且在检验中联结性表现不显著的种对也可能存在关联。联结系数Ac显示处于显著正联结(32对)和显著负联结(33对)对数接近,一般来说,负关联比较多的群落处于演替初期,随着演替的进行物种间逐渐趋于正关联,而正负关联比越高,群落结构越趋于稳定[33]。显然扎龙湿地水环境中藻类群落尚未达到稳定阶段,与包头南海湖的发展状况相类似,这可能与较长的冻融期对藻类的影响有关[34]。 1)本研究共检出藻类8门311种,表明扎龙湿地水域藻类物种丰富度较高,尤其在夏季。藻类群落中绿藻门种类占据优势,全年共鉴定藻类优势种18种,以绿藻门种类为主,全年优势种为狭形纤维藻和啮蚀隐藻。其优势种类随季节变化明显,春季金藻门种类优势度明显、夏季绿藻门优势种为主,秋季隐藻门优势度进一步体现。 2)优势种的季节性变化反映了扎龙湿地水体环境的变化趋势:由春季“贫营养的小型浅水水体”向夏季“贫营养混合浅水体”和秋季“中营养的静水浅水水体”变化,因此利用生态位宽度较大的优势种来指示扎龙湿地水体有可靠的生态学意义。 3)生态位重叠值大小可以较好反映优势种间不同季节的竞争关系情况,春季优势种间资源的竞争最为突出,以金藻门种类为主,夏季达到相对稳定状态,绿藻门等优势种占据主导,秋季隐藻门竞争开始加剧。 4)种间联结性分析结果表明,扎龙湿地水体中藻类种间总体联结上呈正关联关系,但是藻类群落中绝大多数优势种间联结性不显著,种间联结较为松散,种与种之间相对独立。 扎龙湿地藻类群落结构生态位测度均存在明显的季节波动,并且较好反映出水环境季节变化特征,运用生态位测度来反映扎龙湿地水体水环境的变化,具有参加价值和生态学意义。2 结果与分析
2.1 扎龙湿地藻类群落结构特征

2.2 藻类优势种生态位




2.3 优势种总体关联性分析


2.4 优势种种间联结性
3 讨论
3.1 优势度和生态位宽度
3.2 生态位重叠值
3.3 优势种种间联结性
4 结论
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
青年文学家(2023年4期)2023-05-30 10:48:04
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
伙伴(2020年4期)2020-05-11 06:15:11
浙江农林大学学报(2019年4期)2019-07-24 08:19:16
经济技术协作信息(2018年22期)2019-01-19 02:59:58
中国真菌学杂志(2018年2期)2018-05-17 01:00:17
青年文学家(2017年28期)2017-11-28 17:27:53
中国真菌学杂志(2016年5期)2016-12-12 10:05:26
小品文选刊(2016年21期)2016-11-27 00:09:57
