
利用高密度遗传图谱定位水稻耐低氧萌发QTL
2023-06-07 03:44闫晓霞朱满山柳武革李金华黄永相刘迪林
广东农业科学 2023年4期
闫晓霞,朱满山,王 丰,柳武革,李金华,霍 兴,黄永相,刘迪林
(1.广东海洋大学滨海农业学院,广东 湛江 524088;2.广东省农业科学院水稻研究所/广东省育种新技术重点实验室/广东省水稻工程实验室/农业农村部华南优质稻遗传育种重点实验室,广东 广州 510640)
【研究意义】水稻(Oryza sativaL.)是我国最重要的粮食作物之一,也是世界上近60%人口的主食[1-2]。随着城镇化发展和农村劳动力转移,水稻机械化和轻简化生产成为一种趋势。与传统的移栽模式相比,水稻直播栽培省却了育秧和移栽环节,可以节约资源和劳动力投入,实现高效生产[3-5]。水稻直播可分为旱直播、湿直播和淹水直播,其中淹水直播有利于控制杂草,减少鸟害和鼠害,但是对品种的淹水发芽能力要求很高[6-7]。“全苗难”是水稻直播面临的核心问题,也是直播稻产量不高不稳的首要因素[7]。因此,挖掘耐低氧萌发相关基因,培育耐低氧萌发的专用品种成为水稻直播生产的迫切需求。【前人研究进展】目前已定位到一些控制水稻低氧萌发的QTL[5,8-19]。侯名语等[8]利用81 个Kinmaze//DV85 重组自交系群体检测到5 个低氧发芽相关QTL,表型贡献率为10.5%~19.6%。Jiang 等[9]利用USSR5/N22 的F2群体,检测到2 个低氧发芽力相关QTL,表型贡献率为15.51%和10.99%。王洋等[10]利用重组自交系群体检测到6 个QTL,表型贡献率在5.8%~16.2%之间。Angaji 等[5]利用Khao Hlan On/IR64 回交群体,发现5 个芽期耐涝性QTL,解释17.9%~33.5%的表型变异。陈孙禄等[11]以R0380/RP2334 回交自交系群体,检测到影响低氧萌发特性的4 个QTL,贡献最大的是qGS2.2,贡献率为17.34%。Septiningsih 等[12]采用IR42/马占红群体定位到6 个QTL,其中7 号染色体上的QTL 贡献率达31.7%。Baltazar 等[13]利用IR64/Nanhi 的F2:3群体中,在7 号染色体上检测到1 个来自Nanhi 的厌氧发芽主效QTLqAG7,解释表型变异的22.3%。Yang 等[14]用YZX/02428 衍生的重组自交系群体,检测到25 个与低氧发芽能力相关的QTL。Baltazar 等[15]从IR64/Kharsu 80A F2:3群体中定位到4 个与厌氧发芽相关的QTL,解释表型变异的8.1%~12.6%。牛世朋等[16]在Asominori/IR24 重组自交系群体中检测到3 个与中胚轴伸长相关的QTL,效应值最大的QTL 贡献率为11.07%。Ghosal 等[17]利用两个分离群体,发现5 个淹水存活率相关QTL 和4 个控制苗高的QTL。张所兵等[18]采用扎西玛/南粳46 重组自交系群体,检测到qAG-12,表型贡献率为11.24%。Liu 等[19]利用93-11 背景的片段导入系,发现两个与胚芽鞘伸长有关的QTLqAGP1和qAGP3,贡献率均达到15%。通过全基因组关联分析,也发现一些和水稻低氧萌发性显著关联的位点[20-23]。例如,孙凯等[20]采用200份水稻种质进行关联分析,分别鉴定到8 个活力指数相关QTL 和15 个与胚芽鞘相关的QTL。此外,克隆了4 个低氧萌发基因[24-27]。其中,OsTPP7是首个通过正向遗传学途径分离的耐低氧萌发基因,通过调控海藻糖6-磷酸的代谢,增强水稻厌氧萌发的耐受性[24]。OsCBL10是淹水条件下低氧信号通路的负调控因子,其启动子序列差异决定了基因效应,在耐淹型水稻品种中OsCBL10的表达水平极低[25]。利用全基因组关联分析鉴定到控制缺氧萌发的基因CLSY1,可能通过甲基化途径参与低氧萌发性的调控[26]。最近,从杂草稻中克隆到控制低氧萌发出苗的关键基因OsGF14h,编码一个14-3-3 蛋白,导入栽培稻后可以显著提高直播萌发率和成苗率[27]。【本研究切入点】目前尽管在水稻耐低氧萌发基因挖掘方面取得一些进展,但仍存在一些突出问题。一是田间表型受环境影响大,导致基因克隆难度大;二是很多研究采用了SSR 和RFLP 等传统低通量标记,遗传图谱的标记密度不高,导致QTL 定位区间较大。因此,适合育种利用的基因仍然十分缺乏,亟需挖掘更多新的芽期耐低氧基因/QTL。【拟解决的关键问题】针对这些问题,本研究利用外引直播稻品种Francis 和我国南方稻区大面积应用的骨干恢复系R998 创制RIL 群体,构建高密度遗传图谱,并在室内可控环境下开展水稻低氧萌发相关QTL 定位,为芽期耐低氧基因克隆和分子育种提供支撑。
1 材料与方法
1.1 试验材料
以直播型温带粳稻品种Francis 为母本,多穗型优质恢复系R998 为父本进行温汤杂交,从F2代起采用单粒传法构建含有214 个家系的RIL 群体。
1.2 试验方法
2022 年4—12 月在广东广州市天河区广东省农业科学院水稻研究所开展试验。从重组自交系群体各家系选取20 粒健康饱满的种子,自来水冲洗3 次,放入体积为460 mL 的透明塑料杯中,杯中加入10 cm 深的灭菌水,杯口盖上培养皿,在28 ℃、相对湿度75%的黑暗条件下淹水7 d,保持淹水深度不变,以此来模拟低氧萌发环境。淹水处理7 d 后,将萌发的种子取出,观察萌发和生长情况,用直尺测量幼苗的胚芽鞘长(coleoptile length,CL)、芽长(shoot length,SL)和最大根长(root length,RL)。
1.3 遗传图谱构建
采用CTAB法提取Francis、R998 及214 个家系的基因组DNA,检测合格的DNA 样品经过酶切、加测序接头、纯化、PCR 扩增等步骤完成文库制备。构建好的文库通过Illumina HiSeq 测序平台进行测序,委托华大基因完成。测序数据根据亲本间SNP 过滤后得到SNP 位点。采用窗口滑动的方法,每15 个SNP 为1 个窗口,最终获得3 106 个bin 标记,利用MSTMap 软件构建重组自交系群体的高密度遗传连锁图谱[28]。
1.4 水稻低氧萌发QTL 定位
整合表型数据和基因型数据,采用WinQTL Cart 2.5 进行QTL 扫描[29],检测低氧萌发性状相关QTL,LOD 阈值设定为2.5。QTL 的命名遵循McCouch 命名原则[30]。
2 结果与分析
2.1 双亲及群体低氧萌发表型分析
在淹水条件下发芽7 d 后,对水稻亲本Francis和R998 的低氧萌发性进行调查,测量其胚芽鞘长、芽长和最大根长。结果发现,Francis 的胚芽鞘长、芽长和最大根长的均值分别为3.03、3.40、0.74 cm,R998 的胚芽鞘长、芽长和最大根长的均值分别为1.99、2.73、0.57 cm。Francis 的胚芽鞘长度和最大根长显著高于R998(P<0.05,图1、表1),但两者芽长差异不显著。
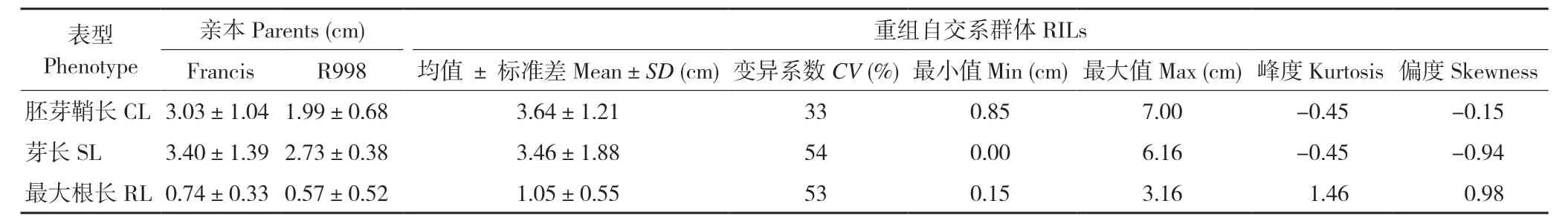
表1 Francis/R998 RIL 群体低氧萌发表型统计Table 1 Statistics of phenotype among the RIL population of Francis/R998 under hypoxia germination
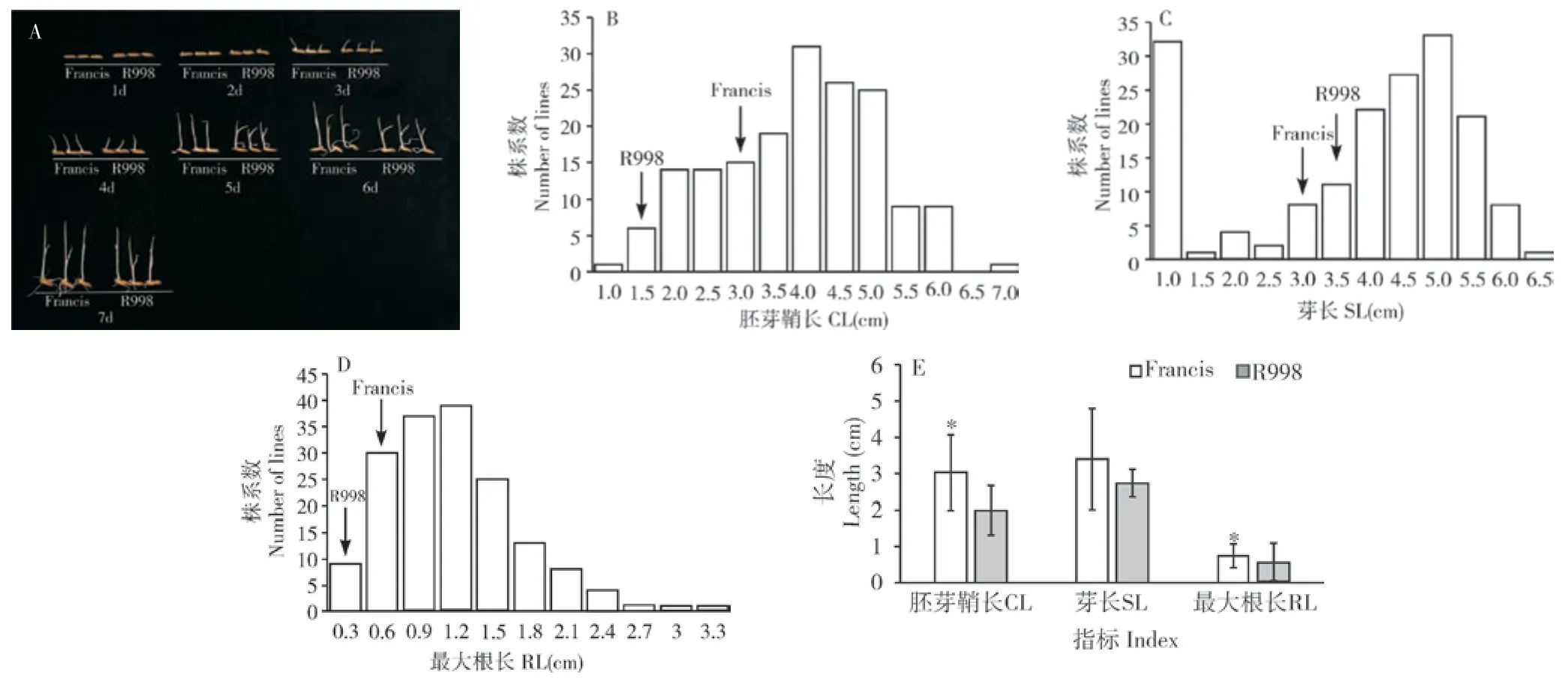
图1 Francis/R998 RIL 群体低氧萌发特性的表型分布Fig.1 Phenotypic distribution of the RIL population of Francis/R998 under hypoxia germination
在重组自交系群体中,各家系芽长和最大根长均值分别为3.90 cm 和0.62 cm,中位数分别为3.86 cm 和0.66 cm,胚芽鞘长的平均值为2.97 cm,中位数为3.31 cm,最大值为7.00 cm,最小为0.85 cm,变异系数为0.28。群体中有多个家系的胚芽鞘长度表现出超亲现象。因此,Francis/R998的RIL 群体各家系间在低氧萌发性上具有较大的表型变异。从表型频率分布看,3 个表型在群体内均呈现出正态分布,除了最大根长的峰度为1.46,其余的峰度和偏度绝对值都小于1,适合进行QTL 作图(图1 B~D)。
2.2 水稻低氧萌发的标记连锁图谱构建
利用低倍基因组重测序数据,构建了水稻Francis/R998重组自交系群体的高密度遗传图谱。根据窗口滑动的方法,得到每个家系的基因型并生成bin 图,最后上图的bin 标记总数为3 106 个,其中12 号染色体标记数最少,1号染色体标记数最多,分别为174 个和389 个;总图距3 646.2 cM,4 号染色体遗传图距最长,而9 号染色体最短,分别为399.6 cM 和220.8 cM(图2)。整张遗传图谱的标记平均图距为1.21 cM,平均图距范围是0.68~1.84 cM,大于5 cM的Gap比例为0.36%。因此,构建的遗传图谱标记密度均匀,图谱质量较好,完全满足QTL 定位的要求。

图2 Francis/R998 RIL 群体耐低氧萌发性QTL 的染色体位置Fig.2 Chromosome locations of hypoxia germinationrelated QTLs in the RIL population of Francis/R998
2.3 水稻低氧萌发QTL 定位
利用软件WinQTL Cart 2.5 的ICM 法对水稻低氧萌发表型进行QTL 扫描。结果表明,在淹水条件下共检测到6 个与低氧萌发相关的QTL(表2、图2)。控制胚芽鞘长的QTL 有3 个(图3),其中qCL1位于1 号染色体标记区间bin364~bin369,LOD 值为2.62,贡献率为5.05%,加性效应-0.43;qCL9位于9 号染色体标记区间bin79~bin83,LOD 值为6.67,贡献率为13.39%,加性效应0.69;qCL12位于12 号染色体标记区间bin41~bin45,LOD 值为2.76,贡献率为4.93%,加性效应-0.43。qCL9的表型贡献率大于10%,是一个主效位点。qCL1和qCL12的加性效应是负值,表明增效等位基因均来自R998;qCL9加性效应为正值,表明增效等位基因来自Francis。

表2 Francis/R998 RIL 群体中检测到的耐低氧萌发性QTLTable 2 QTLs for hypoxia germination tolerance detected in the RIL population of Francis/R998

图3 Francis/R998 RIL 群体低氧萌发胚芽鞘长度的 QTL 扫描峰图Fig.3 QTLs peak of coleoptile length under hypoxia germination in the RIL population of Francis/R998
控制芽长的QTL 只有1 个,即位于5 号染色体上的qSL5,LOD 值为5.00,标记区间位于染色体上bin169~bin173,贡献率为10.78%,加性效应为-0.85。该位点的表型贡献率大于10%,也是主效位点,加性效应分析表明,增效等位基因来自R998。
控制根长的QTL 有2 个,均位于2 号染色体上,命名为qRL2-1、qRL2-2,LOD 值分别为3.31和2.84,位于2 号染色体标记区间bin261~bin267和bin202~bin210,贡献率分别为6.42%、6.47%,加性效应分别为-0.20、0.19。值得注意的是,这两个QTL 的效应值相当、作用方向相反,qRL2-1的加性效应是负值,表明增效等位基因来自R998,qRL2-2的加性效应是正值,增效等位基因来自Francis。
2.4 低氧萌发QTL 中候选基因筛选
从定位到的QTL 中,我们选取效应值最大的两个位点qCL9和qSL5作进一步分析,在目标区间上下游500 kb 范围内筛选可能的候选基因。其中胚芽鞘长位点qCL9位于9 号染色体12.63~13.04 Mb 范围,距该区间上游约400 kb 的12.25 Mb 处存在1 个已克隆基因OsTPP7,该基因编码1 个海藻糖-6-磷酸磷酸酶,也是首个通过图位克隆分离到的低氧萌发条件下控制胚芽鞘伸长的基因[24]。因此,OsTPP7可能是胚芽鞘长相关位点qCL9的候选基因。此外,从已有的转录组数据中,在芽长位点qSL5区间发现对淹水胁迫具有响应的9 个基因,其中3 个下调,6 个上调(表3)。
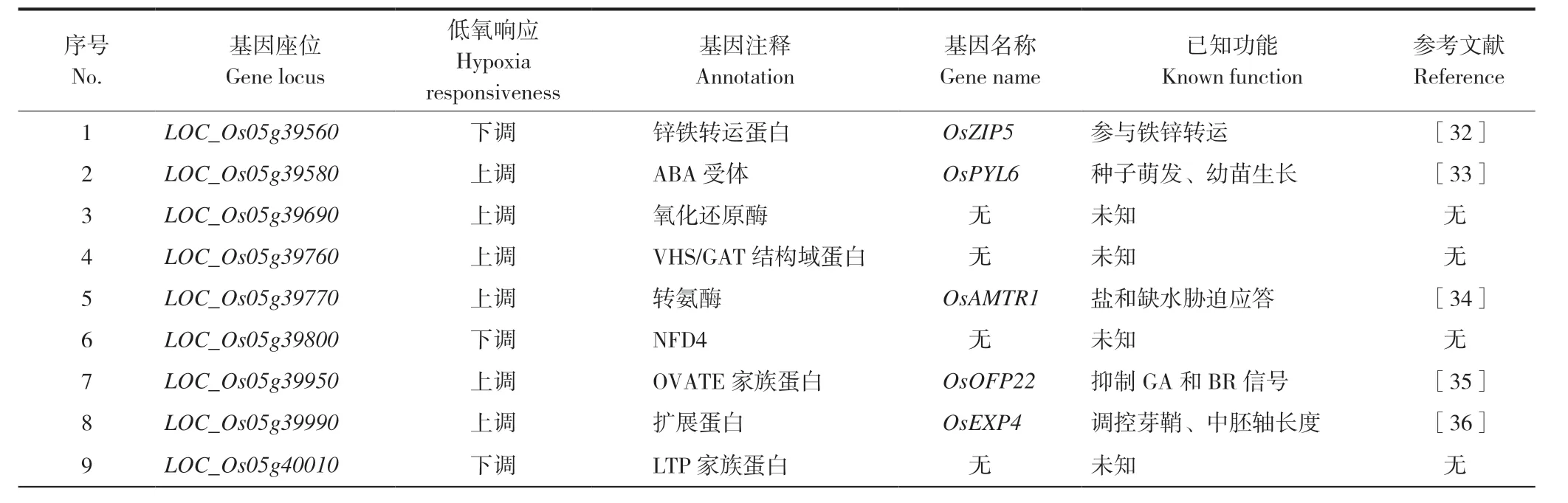
表3 qSL5 位点的可能候选基因Table 3 Potential candidate genes for the locus qSL5
从功能相关性看,最紧密的是LOC_Os05g 39580和LOC_Os05g39990,LOC_Os05g39580可能为ABA 受体,主要影响种子萌发和幼苗早期生长,LOC_Os05g39990 则编码一个扩展蛋白,过表达植株会促进胚芽鞘和中胚轴的伸长[33,36]。其余7 个基因分别编码锌铁转运蛋白、氧化还原酶、VHS/GAT 结构域蛋白、转氨酶、NFD4 蛋白、OVATE 家族蛋白和LTP 家族蛋白(表3)。
3 讨论
随着农村劳动力转移和农业科技进步,水稻生产方式正在发生重大变迁,机械化和轻简化生产势在必行,其中淹水直播是水稻轻简化生产的重要发展方向,不但具有省工、省时和高效等优势,还具有防草防鸟防鼠的功效,是南方地区理想的直播生产方式。在直播稻生产中,淹水带来的低氧条件下,水稻种子萌发和出苗受到影响,显著降低水稻的成苗率。低氧萌发成苗能力强的专用品种缺乏,成为影响直播稻发展的瓶颈。因此,加快淹水直播专用品种的培育,亟需对水稻芽期耐低氧萌发性基因和QTL 进行挖掘,为分子育种提供更多的有利基因资源。在芽期耐低氧萌发QTL 研究中,材料的选择十分关键。本研究采用的RIL 群体材料衍生自亲本Francis 和R998,其中F rancis 是国外的直播型温带粳稻品种,R998 是我国南方稻区大面积应用的优质多穗型恢复系,二者具有丰富的遗传差异,且在低氧萌发方面有显著差异,是开展耐低氧萌发QTL 研究的理想材料。
前人研究表明,水稻种子在低氧条件下萌发时,根和叶的生长受到抑制,但是胚芽鞘可以快速伸长到达有氧环境,为根和叶的生长提供足够氧气。一般认为,耐低氧萌发的水稻品种胚芽鞘伸长能力比不耐低氧萌发的品种强。在耐低氧萌发性的衡量指标方面,以往的研究普遍采用低氧条件下萌发的芽长[8]、胚芽鞘长[10]、缺氧反应指数[37]、淹水成苗率[38]等。本研究则采用淹水暗培养7 d 的芽长、胚芽鞘长和最大根长作为衡量指标。侯名语等[8]从籼粳分化角度比较,认为水稻品种的低氧发芽力存在籼粳差异,粳稻的低氧发芽力一般比籼稻强。类似地,Rauf 等[39]比较了来自6 个水稻亚群的185 份水稻材料,发现低氧萌发条件下粳稻的发芽率和胚芽鞘长度整体优于其他亚群,温带粳稻的平均胚芽鞘长最大。本研究中采用的温带粳稻Francis 胚芽鞘伸长能力显著好于籼稻R998。因此,从粳稻中筛选耐低氧萌发的种质,值得引起重视。
前人多项研究表明,水稻低氧萌发性是多基因控制的数量性状[8-15]。本研究采用高密度遗传图谱,定位到6 个低氧萌发相关的QTL。其中有3 个QTL 与前人结果处于相同或相近位置,如9号染色体上的胚芽鞘长度QTLqCL9(物理位置:12.63~13.04 Mb),与已克隆的OsTPP7位置临近(12.25 Mb 处)[24]。尽管二者是否由同一基因控制还需确认,但是这类在不同研究中重复鉴定、效应稳定的QTL,在耐低氧遗传机制和育种应用中均有较大价值。
2 号染色体上控制根长的QTLqRL2-1(物理位置:30.38~30.63 Mb),与Septiningsih 等[12]检测到的淹水发芽QTLqAG2临近(29.9 Mb 处);2 号染色体上控制根长的QTLqRL2-2,位于25.78~26.09 Mb 区间,与控制淹水成苗率的qSES2存在区间重叠(24.04~26.5 Mb)[31],它们可能是相同的QTL。
此外,本研究定位到的QTL 有3 个暂未见报道。胚芽鞘长度QT LqCL1和qCL12,分别位于1 号染色体41.55~42.02 Mb 和12 号染色体36.06~37.09 Mb 区间;芽长QTLqSL5,位于5 号染色体23.18~23.59 Mb 区间,这3 个位点附近暂未见低氧萌发相关QTL 的报道,可能是控制低氧萌发性的新位点。在qSL5区间筛选到9 个可能的候选基因,其中LOC_Os05g39580和LOC_Os05g39990可能性最大。LOC_Os05g39580是潜在的ABA 受体;LOC_Os05g39990编码一个扩展蛋白,过表达植株会促进胚芽鞘和中胚轴的伸长[33,36]。
不同水稻品种的低氧萌发能力不同。从品种选育角度看,耐低氧萌发QTL 位点的发掘对于耐低氧萌发水稻品种的定向培育具有重要意义。一般认为,贡献率在10%以上的主效QTL 具有应用价值。前人克隆的OsTPP7已经用于直播稻品种的改良,定向导入栽培稻品种后,耐低氧萌发能力有不同程度的提升[40]。本研究定位的QTL中效应值最大的是胚芽鞘位点qCL9和芽长位点qSL5,效应值均在10%以上,增效等位基因分别来自Francis 和R998,在育种中可以利用分子标记进行前景选择将二者进行聚合,创制耐低氧萌发能力显著提升的新种质。此外,良好的低温发芽力也是直播品种所需的关键性状[41-42],本研究下一步也将采用该群体进行低温发芽性状的遗传分析。
4 结论
淹水直播是水稻生产的重要发展方向。然而,低氧萌发性强的专用品种缺乏,成为影响直播稻发展的瓶颈。加快淹水直播专用品种的培育,亟需对水稻耐低氧萌发性基因和QTL 进行挖掘。本研究以Francis 和R998 衍生的重组自交系群体为材料,检测到6 个低氧萌发相关的QTL,其中3个可能是新位点。两个主效QTLqCL9和qSL5贡献率均大于10%,在分子育种中具有应用潜力,胚芽鞘长度QTLqCL9的增效等位基因来自外来种质Francis,芽长QTLqSL5的增效等位基因来自骨干恢复系R998,在分子育种中可将这些位点通过杂交、回交和分子标记辅助选择导入新品系中,提高水稻品种的耐低氧萌发能力。在qSL5区间筛选到9 个候选基因,可进一步鉴定这些候选基因的表达和功能,为解析水稻淹水直播机制奠定基础,为培育直播稻新品种提供新的基因资源。
猜你喜欢
中学生物学(2022年8期)2022-10-13
天津医科大学学报(2021年3期)2021-07-21
军事运筹与系统工程(2020年2期)2020-11-16
生物学通报(2020年10期)2020-08-13
浙江农业学报(2019年2期)2019-03-05
军事运筹与系统工程(2018年3期)2018-03-26
教学考试(高考生物)(2016年2期)2016-08-11
国外医药(抗生素分册)(2016年5期)2016-07-12
中亚信息(2016年10期)2016-02-13
实验动物与比较医学(2014年5期)2014-02-28
