
梨园土壤真菌群落垂直分布及功能预测分析
2023-05-30 22:11:13冀明辉高丽娟李龙飞徐金涛郝宝锋
山东农业科学 2023年3期
冀明辉 高丽娟 李龙飞 徐金涛 郝宝锋
关键词:梨园:真菌:土壤:垂直分布:差异物种:功能预测
中图分类号:S661.2:Q938.1 文献标识号:A 文章编号:1001-4942(2023) 03-0083-12
土壤、微生物、植物間相互联系紧密,土壤是植物吸收营养的重要场所,直接影响植物的生长发育,对微生物群落结构及多样性影响显著。通过对不同深度土壤微生物的研究,能更确切地了解微生物在土壤中的变化。土壤硝态氮、有机碳能显著影响土壤真菌群落结构。不同类型土壤微生物群落组成各不相同,作用方式复杂多样。陈娟等研究发现潮土相对于稻田土和黄棕土抗病性更强。微生物为生态系统中重要的一环,通常真菌与细菌通过相互作用来调节土壤生态系统。大多数真菌生活在土壤或植物残体中,可增强土壤微生物酶活性。Li等发现通过接种外生菌根能提高土壤基质可用性,减轻酸雨对土壤中氮循环微生物的影响。但真菌对植物也有不利影响,土壤连作可增加真菌数量,降低土壤质量,抑制植物生长。研究发现Cortinarius acutu s.l.会导致表土中局部碳储存降低33%。子囊菌门(Asco-mycota)会导致植物枯萎,担子菌门(Basidiomyco-ta)会引起小麦锈病、玉米黑穗病等多种病害,壶菌门(Chytridiomycota)会诱发植物猝倒病、癌肿病,镰刀菌属(Fusarium,)通常作为植物病原菌存在。细菌或真菌病害往往伴随着土壤微生物群落的变化。黄伟雄等研究发现,荔枝在感染根腐病后,土壤中真菌群落多样性下降,种类减少,均匀度上升。感染根结线虫的土壤和健康土壤在真菌门类上有明显差异。韩凤等发现根腐病改变了多花黄精根际土壤的真菌组成。目前在改善微生物群落多样性方面的研究较多。刘业萍、程分生等研究表明,生草能显著提高土壤理化性质,改善土壤微生物群落结构,提高土壤中有机碳含量。通过轮作可降低真菌丰度,改善真菌群落组成,调节微生物功能。除此之外,一些学者通过土壤消毒或生物菌剂可达到平衡微生物的目的。赵晓东、陈利达等采用熏蒸处理可抑制土壤镰孢病菌,改善土壤结构,调节真菌群落。刘亮亮等通过土壤强还原消毒增加了有益真菌的相对丰度,降低西瓜枯萎病的发病率。王辉等利用黄柄曲霉可提高土壤营养状况,降低辣椒疫病带来的影响。木霉菌剂能使茉莉根际土壤微生物明显变化,提高其生长速率。氯吡嗪与多唑醇交替使用,能有效改善土壤理化性质、微生物群落,提高草莓产量。随后,绿色、可持续的研究理念逐渐增强。屈忠义、姚彤等利用生物炭来调节土壤理化性质,可改变真菌丰度,增加土壤肥力。刘莉等指出,枝条粉碎还田能提高土壤有机质含量,增加微生物数量。有学者利用水稻和鱼类共生系统来减少化肥农药的施用,可提高土壤质量,调节优势真菌丰度。
目前,土壤真菌群落研究主要集中在通过轮作、生草、消毒、共生等方式改善真菌群落结构,提高土壤质量,而忽略了土壤垂直分布微生物群落差异。基于此,本试验以梨园土壤为研究对象,通过对不同深度土壤进行理化性状测定及ITS测序分析,对真菌群落在土壤中的垂直分布情况及功能进行预测分析,以期为梨园土壤微生物群落的改善等提供理论参考。
1材料与方法
1.1样品采集
2021年4月,于河北省农林科学院昌黎果树研究所孔庄示范基地梨园(2009至今,土壤管理模式为清耕)进行土壤样品采集。由于距梨树主干Im左右,根系主要分布在距地面10~30 cm的土层内,所以本试验选取距主干1m左右、深度为5~35 cm土壤,采用“S”形取样法,分别取5~15cm(A处理)、15~25 cm(B处理)、25~35cm(C处理)三种不同深度土壤各5份,共计15份,将采集到的不同深度土样分别混匀,采用四分法留样。样品分为两份,一份用于土壤理化性质测定,另一份用于真菌ITS1-5F区测序分析。
1.2土壤理化性质测定
土壤有机质含量的测定采用重铬酸钾容量法(外加热法)。土壤碱解氮含量的测定采用碱解扩散法,土壤速效磷含量的测定采用碳酸氢钠提取-钼锑抗比色法,土壤速效钾含量的测定采用乙酸铵提取-原子发射光度法。
1.3土壤DNA的提取与分析
采用CTAB法对土壤样品进行DNA提取。采用Phusion High-Fidelity PCR Master Mix试剂盒(New England Biolabs)进行PCR扩增,引物序列为:ITS5-1737F(5′-GGAAGTAAAAGTCGTAA-CAAGG-3′),ITS2-2043R(5′-GCTGCGTTCT-TCATCGATGC-3′)。PCR扩增程序:98C预变性1min;98℃变性10s,50℃退火30s,72℃延伸30s,共30个循环,720C延伸5 min。随后进行文库构建、上机测序、下机数据拆分,整理得到有效数据。最后以97%的一致性将序列聚类成为OTUs,同时筛选OTUs的代表序列并进行物种注释分析。利用诺禾云平台(https://magic. novogene.com/custom-er/main#/login)基于有效数据进行OTUs聚类和物种注释、α多样性分析、Venn图绘制等。
1.4数据分析
所有数据经Microsoft Excel 2019进行处理,采用SPSS 20.0进行单因素方差分析。土壤理化性质与真菌群落的相关性采用Pearson相关系数法进行分析。
2结果与分析
2.1土壤理化性质
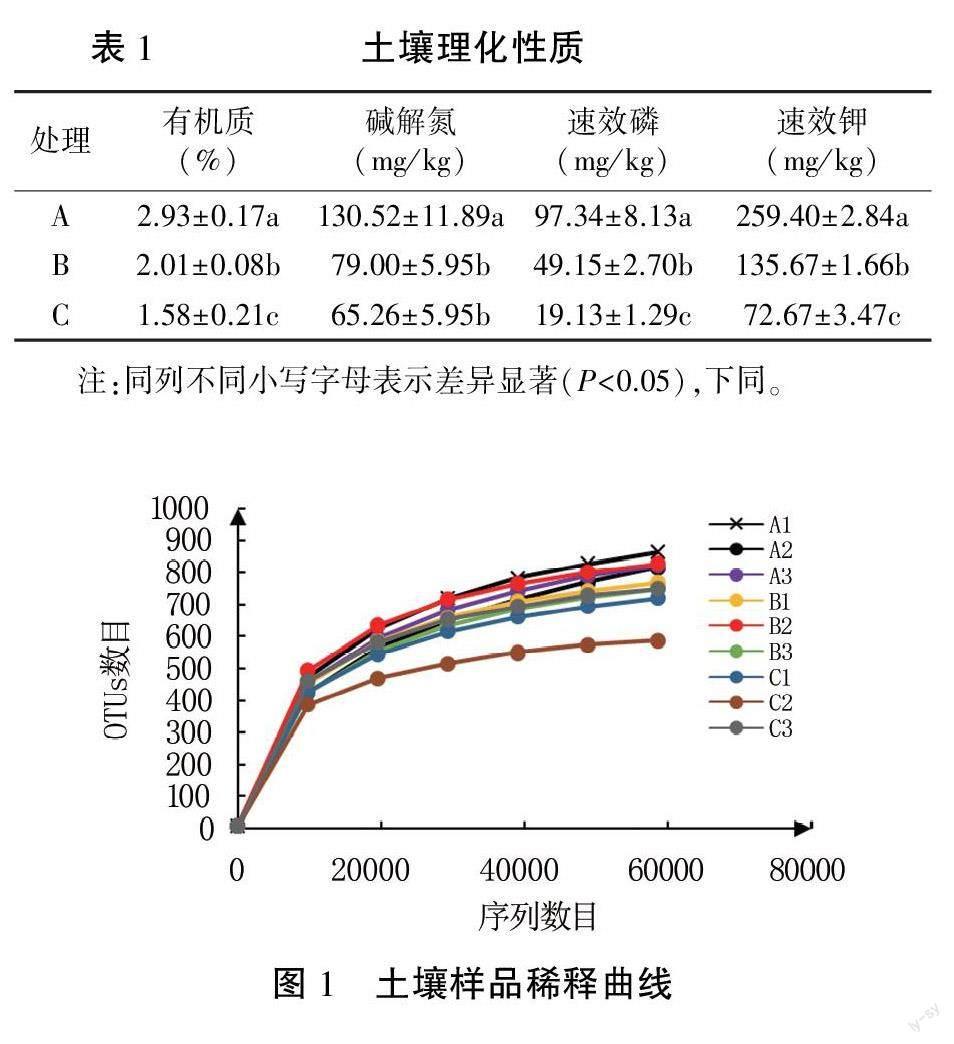
由表1可知,有机质(SOM)、碱解氮(AN)、速效磷(AP)、速效钾(AK)均随土壤深度的增加而降低,且降低速率逐渐减缓。其中SOM、AP、AK在处理A、B、C中均呈显著性差异。A处理SOM含量为B处理的1.46倍、C处理的1.85倍;A处理AN含量为B处理的1.65倍、C处理的2.00倍;A处理AP含量为B处理的1.98倍、C处理的5.09倍:A处理AK含量为B处理的1.91倍、C处理的3.57倍。所以,四种土壤养分随土壤深度的变化速率为SOM
2.2土壤样品测序深度分析
为保证测序数据的准确性,对土壤样品测序数据进行分析。如图1所示,曲线逐渐趋于平缓,说明测序数据合理。A处理共获得Raw Tags91172条,拼接和质控后,得到Clean Tags 90 983條,嵌合体过滤后,得到Effective Tags 65 252条;B处理共获得Raw Tags 93 131条,拼接和质控后,得到Clean Tags 88 402条,嵌合体过滤后,得到Effective Tags 65 029条;C处理共获得Raw Tags 95 239条,进行拼接和质控,得到Clean Tags 94 674条,嵌合体过滤后,得到Effective Tags63 748条。
2.3不同土壤深度真菌群落多样性分析
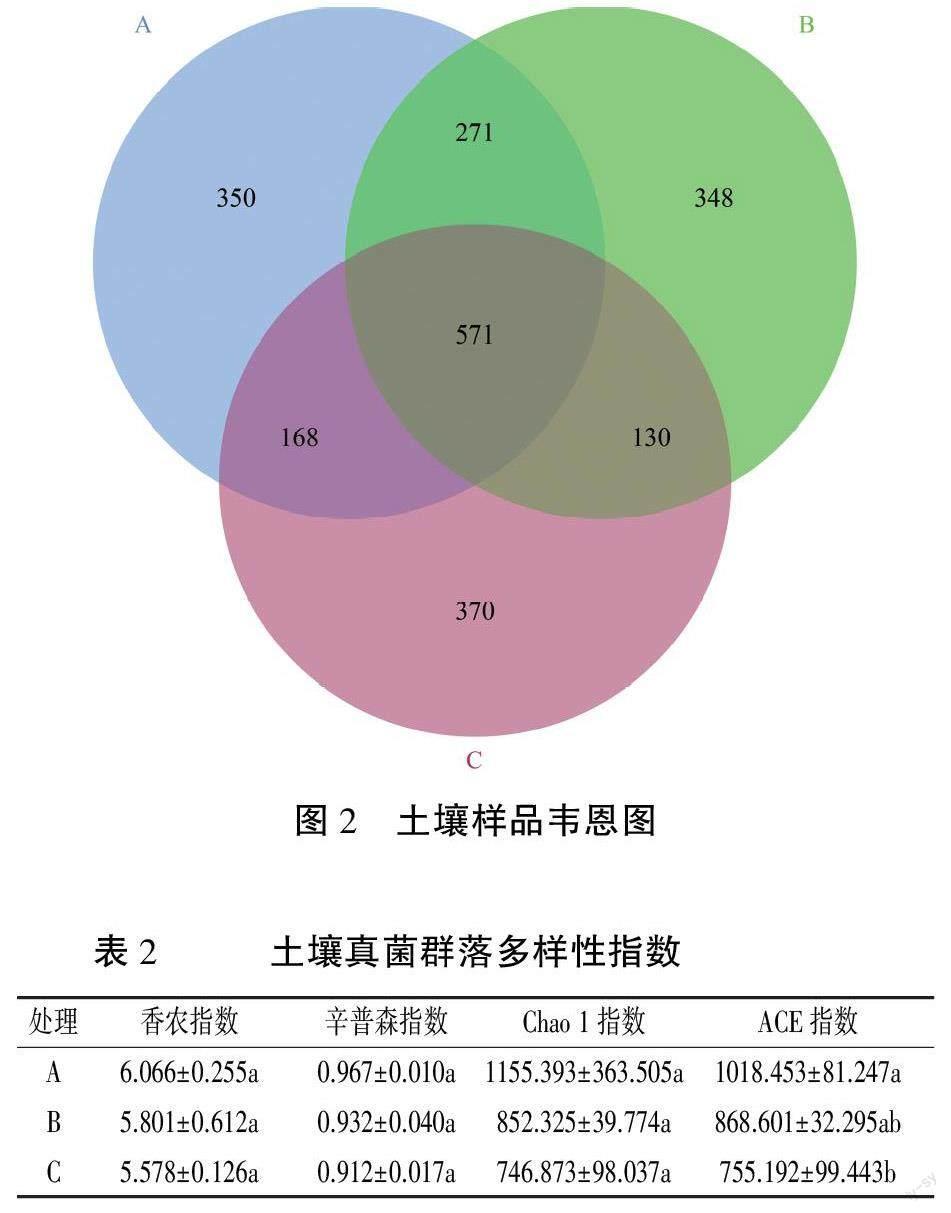
由图2可知,A处理共有OTUs 1360个,B处理共有OTUs 1 320个,C处理共有OUTs 1 239个。A、B处理相同OTUs为842个,分别占A、B处理的61.91%、63.79%;B、C处理相同OTUs为701个,分别占B、C处理的53.11%、56.58%;A、C处理相同OTUs为739个,分别占A、C处理的54.34%、59.64%。A、B、C处理相同OTUs为571个,分别占A、B、C处理的41.99%、43.26%、46.09%。A、B、C处理各自特有的OTUs为350、348、370个,分别占A、B、C处理的25.74%、26.36%、29.8%。
如表2所示:随着土壤深度的增加,香农指数和辛普森指数均逐渐降低,说明物种多样性逐渐降低,但各处理均无显著差异。Chao 1指数和ACE指数也随土壤深度的加深逐渐降低,说明物种的丰度逐渐减少。
2.4不同土壤深度对真菌群落结构的影响
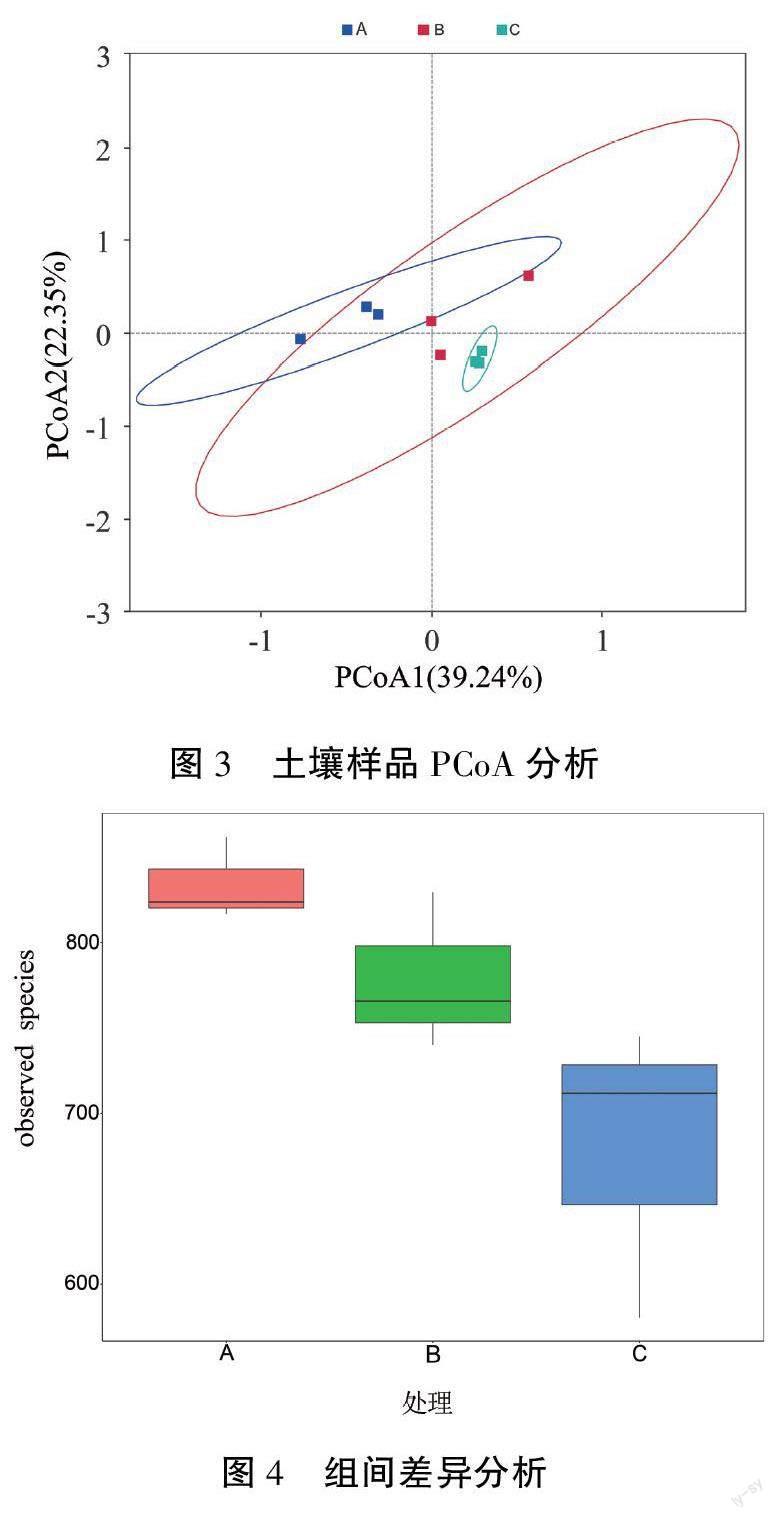
采用weighted_unifrac算法对土壤样品进行PCoA分析,横、纵坐标表示两个主成分,百分比表示其贡献值;样本距离越接近,物种组成结构越相似。由图3可知:第一主成分各处理间A与C的距离最远,B与C之间的距离最近;第二主成分各处理间A、B间距离较小。说明第一主成分A、C处理群落结构差异性大于A、B处理与B、C处理,第二主成分上A处理与B处理几乎无差异,均与C处理有较小差异。综合第一、第二主成分分析发现:随着土壤深度加大,土壤真菌结构逐渐差异化,A、C处理间的群落差异大于A、B处理和B、C处理。
通过MRPP检验可知:A与B处理、A与C处理、B与C处理组间差异均大于组内差异。为了解组间差异是否显著,绘制图4所示箱形图。箱子的中位数A>B>C,说明A处理物种多样性大于B、C处理。箱子的上下边缘代表95%置信区间,A箱子最扁,其次是B、C,说明A处理均一性好于B、C处理。通过Tukey检验可知:A与B、A与C、B与C处理的P值分别为0.433、0.038(P<0.05)、0.202,说明A、C处理间群落多样性差异显著,A与B、B与C处理间无显著性差异。
2.5不同土壤深度优势种群分析
为进一步了解不同土层间物种差异,对真菌种类进行分析。通过OUT注释分析得,A、B、C处理共有门类16个,纲类57个,目类135个,科类282个,属类470个,种类677个。取相对丰度前10的门类和属类进行分析。如图5a所示:土壤真菌门水平以子囊菌门(Ascomycota)、被孢霉门(Mortierellomycota)、担子菌门(Basidiomycota)为主。其中子囊菌门在A、B、C处理中的占比分别为51.98%、51.57%、61.81%,为丰度最高的门类;被孢霉门在A、B、C处理中的占比分别为10.96%、16.02%、10.33%;担子菌门在A、B、C处理中的占比分别为12.63%、13.63%、7.14%。子囊菌门在C处理中含量最多,担子菌门、被孢霉门在B处理中含量最多,这说明真菌主要门类大部分分布在15~35 cm的土层中。如图5b所示:属水平上,主要由镰刀菌属(Fusarium,)、被孢菌属(Mortierella)、产油菌属(Solicoccozyma)组成。其中镰刀菌属在A、B、C处理中的占比分别为11.01%、18.51%、28.75%;被孢菌属在A、B、C处理中的占比分别为4.39%、13.81%、9.94%;产油菌属在A、B、C处理中的占比分别为9. 58%、4.72%、2.98%。
2.6不同土壤深度差异物种分析
通过对门水平处理间进行t-test差异物种分析,发现B、C处理间无差异物种。A处理罗兹菌门(Rozellomycota)相对丰度极显著大于B处理(图6a);A处理担子菌门(Basidiomycota)相对丰度显著大于C处理,罗兹菌门相对丰度极显著大于C处理,子囊菌门( Ascomycota)相对丰度显著小于C处理(图6b)。其中罗兹菌门在A与B、A与C处理间的相对丰度均呈显著性差异,说明其对土壤的敏感性较高。
属水平上,A、B处理差异物种共有13种,其中以产油菌属(Solicoccozyma)为主,A处理极显著高于B处理(图7a)。A、C处理差异物种共有18种,主要为镰刀菌属(Fusarium)、产油菌属。A处理产油菌属极显著高于C处理:C处理镰刀菌属(Fusarium)极显著高于A处理(图7b)。B、C处理差异物种共有8种,以瓶毛壳属(LophDtrichus)、姆拉克酵母菌属(Mrakia)为主。B处理瓶毛壳属、姆拉克酵母菌属显著高于C处理(图7c)。综上,差异物种数量随着土壤深度增加逐渐减少。
2.7功能多样性预测分析
为探究不同土壤深度真菌功能是否存在差异,选取土壤中真菌生态功能前10类进行统计。由图8可知:随着土壤深度的逐渐增加,未分配(Un-assigned)的类别逐渐减少。属于腐生营养的真菌分别占A、B、C处理的23.89%、34.39%、26.48%,分别为未界定腐生菌(Undefined_Saprotroph,A:21.66%、B:28.14%、C:24.27%)、外生菌根(Ectomycorrhizal,A:0.42%、B:4.99%、C:1.14%)、未界定腐生菌-木质腐生菌(Undefined_Saprotroph-Wood—Saprotroph,A:1.80%、B:1.26%、C:1.07%),此三种生态功能为真菌中主要的功能;其次为病原-腐生营养型,分别占A、B、C处理的11.87%、21.41%、31.99%,具体功能为植物病原菌-土壤腐生菌-木质腐生菌(Plant_Pathogen-Soil_Saprotroph-Wood _Saprotroph,A:11.01%、B:18.51%、C:28.75%)、动物病原菌-土壤腐生菌(Animal_Pathogen-Soil_ Saprotroph,A:0.73%、B:2.77%、C:1.86%)、动物病原菌-内生菌-真菌寄生菌-植物病原菌-木质腐生菌(Animal_Pathogen-Endophyte-Fungal_P arasite-Plant_Pathogen-Wood_Saprotroph,A:0.13%、B:0.12%、C:1.38%);病原营养型分别占A、B、C处理的8.29%、2.26%、6.25%,具体功能为植物病原菌(Plant_Pathogen,A:7.00%、B:2.23%、C:6.24%)、动物寄生菌-真菌寄生菌(Animal_Parasite-Fungal_Parasite,A:1.29%、B:0.03%、C:0.01%);丰度最少的为共生-腐生营养型的动物共生菌-未界定腐生菌(Animal_Endosymbiont-Undefined_Saprotroph),分别占A、B、C处理的0.05%、0.07%、1.43%。
由图9可以看出,A、B处理显著差异功能为动物病原菌-土壤腐生菌(Animal_Pathogen-Soil_Saprotroph),且B处理显著大于A处理;A处理在未分配(Unassigned)类型上极显著大于C处理,在植物病原菌-土壤腐生菌-木质腐生菌(Plant_Pathogen-Soil_Saprotroph-Wood_Saprotroph)上极显著小于C处理,在粪腐生菌-未界定腐生菌-木质腐生菌(Dung_Saprotroph-Undefined_Saprotroph-Wood_Saprotroph)、外生菌根(Ectomycorrhizal)上显著小于C处理。B处理中的动物病原菌(Ani-mal_Pathogen)显著大于C处理,叶片腐生菌(Leaf_Saprotroph)极显著小于C处理。
2.8土壤理化性质与真菌门类及其功能的相关性
为了解土壤理化性质与真菌间的关系,对其进行相关性分析。真菌丰度前10的门中,担子菌门(Basidiomycota)、油壶菌门(Olpidiomycota)与土壤有机质、碱解氮、速效磷、速效钾含量呈正相关关系;芽枝霉门(Blastocladiomycota)与土壤有机质、碱解氮、速效磷含量呈正相关关系,与速效钾含量呈显著正相关关系;罗兹菌门(Rozellomy-cota)与土壤有机质、碱解氮、速效磷、速效钾含量呈极显著正相关关系。子囊菌门(Ascomycota)、被孢霉门(Mortierellomycota)、壶菌门(Chytridio-mycota)、蛙粪菌门(Basidiobolomycota)、隐孢子菌门(Aphelidiomycota)、捕虫霉门(Zoopagomycota)与土壤有机质、碱解氮、速效磷、速效钾含量呈负相关关系(表3)。
如表4所示:未分配功能(F1)与罗兹菌门(Rozellomycota)呈极显著正相关关系。未界定腐生菌(F2)与子囊菌门(Ascomycota)显著负相关,与被孢霉门(Mortierellomycota)极显著正相关。外生菌根(F3)与担子菌门(Basidiomycota)、蛙粪菌门(Basidiobolomycota)、捕虫霉门(Zoopagomy-cota)显著正相关。植物病原菌-土壤腐生菌-木质腐生菌(F5)与罗兹菌门极显著负相关。动物病原菌-内生菌真菌-寄生菌-植物病原菌-木质腐生菌(F7)与隐孢子菌门(Aphelidiomycota)显著正相关。动物寄生菌-真菌寄生菌(F9)与油壶菌门(Olpidiomycota)极显著正相关。动物共生菌-未界定腐生菌(F10)与壶菌门(Chytridiomycota)极显著正相关。
3讨论
3.1不同土壤深度下真菌群落多样性
随土壤深度逐渐加深,有机质、碱解氮、速效磷、速效钾含量降低,且速率由高到低依次为速效磷、速效钾、碱解氮、有机质,说明速效磷含量对土壤深度反应敏感。25~35 cm土壤主要养分下降了1.85~5.09倍,推测梨园养分主要集中在5~25cm土层中。与土壤理化性质趋势相同,随土壤深度加深,OTUs、真菌多样性指数逐渐降低,但不同土层相同OTUs占40%以上,说明在5~35cm土层中真菌群落同质性较大;真菌多样性指数降低速度放缓,说明土壤深度越大,真菌微生物群落多样性及丰富度变化趋势减小,这与Xue等的研究结果相同。A与C处理间群落差异大于A与B、B与C处理,且经过Tukey检验A与C处理间群落差异具有显著性,同时A处理的多样性及均一度均大于B、C处理。由此可以猜测:群落丰富度越高,群落越稳定,真菌均一性越强;随土壤深度加大,群落丰富度及多样性减小,物种差异增加,群落多样性降低且速率减缓,不稳定性增加。通过相关性分析发现:土壤真菌多样性与有机质、碱解氮、速效磷、速效钾等土壤理化性质关系密切,这与靳海洋、Mayer等的研究结果一致。
3.2不同土壤深度与真菌物种关系
通过对不同土壤深度真菌进行OTUs注释分析,发现5~15 cm的物种最丰富。土壤真菌门类以子囊菌门(Ascomycota)、被孢霉门(Mortierello-mycota)、担子菌门(Basidiomycota)为主,这与满百膺、罗正明等的研究结果一致。其中子囊菌门在25~35cm相对丰度最大且在5~35 cm土层中均达到50%,被孢霉门、担子菌门在15~25 cm土层相对丰度最大,其他类型(物种相对丰度排名10以外的物种)5~15 cm丰度最高,这也从侧面说明5~15 cm的物种多样性较高。属类以镰刀菌属(Fusarium,)、被孢菌属(Mortierella)、产油菌属(Solicoccozyma)为主,分别属于子囊菌门、被孢霉门和担子菌门,随着土壤深度增加,镰刀菌属逐渐增多,产油菌属逐渐降低,被孢菌属为15~25 cm最多。排名前10的属中子囊菌门包含5种,被孢霉门、担子菌门各2种,罗兹菌门(Roz-ellomycota)1种,说明子囊菌门不仅相对丰度大且物种多样性高。
在门水平上,A与B处理差异门类为1种,A与C处理为3种,B与C间无差异菌门;在属水平上,A与B处理差异物种有13种,A与C处理为18种,B与C处理有8种。A、B、C处理无共同的差异物种,说明随着土壤深度加大,真菌多样性差异明显。
在门类与土壤理化性质相关分析中,相对丰度前10的物种,有6类与土壤理化性质呈负相关,4类呈正相关。其中相对丰度大于50%的子囊菌门与土壤理化性质呈负相关,而土壤理化性质越高说明土壤越肥沃,越有利于植物生长,所以推测子囊菌门可能对植物的危害性大于益处。且随着土壤深度增加,子囊菌门相对丰度增加,由此可推测:土壤越深,对植物生长越不利,果树深层根系生长会受到抑制,这可能是梨树根系主要分布在树冠内浅层土中的原因。罗兹菌门与有机质、碱解氮、速效磷、速效钾均呈极显著正相关关系,说明罗兹菌门对土壤养分反应敏感,这与A和B处理、A和C处理间的差异菌门均包含罗兹菌门的结果相互得到验证。
3.3不同土壤深度真菌生态功能分析
真菌中营养类型以腐生、病原-腐生、病原为主,且随着土壤深度增加腐生类型呈先增后减趋势,病原呈先减后增趋势,病原-腐生逐渐增加。这说明15~25 cm土层适合腐生营养型真菌生存而不适宜病原真菌生存,5~25 cm为病原类型相对较少的土层深度,适合植物生存、生长,这与3.2分析中植物适合生活在浅层土壤中结果一致。
由t-test功能差異检验可得:A与B处理的差异功能营养类型为病原-腐生.B与C处理间为病原、腐生,说明随着土壤深度的增加,营养类型逐渐由兼性(病原-腐生)变为单性(病原、腐生),功能性逐渐单一化。原因可能为:土壤越深,土壤环境变化越趋于稳定,且微生物种类越少,竞争越小,真菌营养类型稳定性越强,变化越少,单一的营养类型足以支撑微生物生存;当土壤越浅时,微生物种类增多,竞争越强,需要足够多的营养类型才能够生存。研究指出病原菌、腐生菌与土壤有较强的相关性,这与本试验结果一致。
4结论
(1)土壤深度与土壤理化性质、真菌群落关系紧密,深度增加,土壤养分含量、群落多样性降低。土壤养分主要集中在5~25 cm土层中,且此土层病原类真菌少,适合植物生存。
(2)随土壤深度增加,真菌群落差异增加,差异变化速率、群落均一性、稳定性降低。
(3)真菌群落门水平以子囊菌门(Ascomyco-ta)、属水平以镰刀菌属(Fusarium,)为主,主要分布在15~35 cm土层中。罗兹菌门对土壤变化反应敏感。
(4)土壤真菌以腐生营养型为主,随着土壤加深,营养类型由兼性向单性转化。
猜你喜欢
红领巾·探索(2021年2期)2021-08-26 15:02:02
河北果树(2020年4期)2020-11-26 06:05:46
华人时刊(2020年13期)2020-09-25 08:21:58
科学(2020年2期)2020-08-24 07:56:56
学生天地(2020年34期)2020-06-09 05:50:36
中华戏曲(2020年2期)2020-02-12 05:18:28
当代陕西(2019年11期)2019-06-24 03:40:26
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
北方音乐(2017年16期)2017-10-21 02:07:43
作文通讯·高中版(2017年12期)2017-02-06 05:56:42
