
球孢白僵菌和中华甲虫蒲螨对冷杉梢斑螟杀虫作用及其相关酶活性影响1)
2023-05-23 14:41:46刘行聂振业迟德富贺丽敏
东北林业大学学报 2023年5期
刘行 聂振业 迟德富 贺丽敏
(东北林业大学,哈尔滨,150040) (河北省农林科学院昌黎果树研究所)
冷杉梢斑螟(Dioryctriaabietella)是针叶林的破坏性害虫,广泛分布于朝鲜半岛、欧洲、北美和中国东北[1]。在欧洲,种子园中有很大比例的球果被冷杉梢斑螟侵害,导致种子产量大幅下降[2]。冷杉梢斑螟幼虫主要以球果为食,对球果、种鳞和种子造成危害。幼虫在林下枯枝落叶层中越冬,成虫在每年6月和7月上树产卵,会对红松(Pinuskoraiensis)、赤松(Pinusdensiflora)和云杉(Piceaasperata)等造成危害[3-4]。近年来,该害虫在我国东北大面积发生,严重危害红松球果种实,造成很大的经济损失。因此,控制冷杉梢斑螟种群具有较大的经济意义。
球孢白僵菌(Beauveriabassiana)是一种虫生真菌,在自然界分布广、寄主多,对很多农林害虫致病力强,可侵染15目149科的700余种昆虫[5-7]。对梢斑螟具有较高的毒力和防治潜力[8]。中华甲虫蒲螨(Pyemoteszhonghuajia)作为目前国内重点研究和开发的寄生螨品种,对螟蛾科(Pyralidae)昆虫也表现出了较好的寄生和防治能力[9]。白僵菌一般是配成孢子液,直接喷施到目标生物上,然后通过孢子的侵染作用杀死害虫[10],但防治球果内部的害虫时,害虫因不容易接触到孢子液而成为“漏网之鱼”[11],从而降低了白僵菌的防治效果。中华甲虫蒲螨通过主动搜索的方式寄生寄主[12],从这个角度来说,二者的联合应用可以起到互补作用。本文采用浸虫法和室内接种蒲螨的方法,对2株球孢白僵菌和中华甲虫蒲螨进行室内毒力和杀虫力测定,并从2株球孢白僵菌中筛选出毒力更高的菌株。单独或联合使用球孢白僵菌和中华甲虫蒲螨,研究其对冷杉梢斑螟幼虫僵虫率和死亡率的影响,并测定幼虫被球孢白僵菌和中华甲虫蒲螨侵染或寄生后的谷胱甘肽-S-转移酶(GSTs)、乙酰胆碱酯酶(AChE)2种解毒酶以及多酚氧化酶(PPO)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)3种保护酶的活性变化,旨在为球孢白僵菌与中华甲虫蒲螨联合防控冷杉梢斑螟提供参考。
1 材料与方法
试验材料:冷杉梢斑螟由黑龙江省牡丹江市海林市海林镇森宝源天然食品有限公司提供。培养条件:使用人工气候箱(HPG-400HX),设置温度(25±1)℃,相对湿度(65±5)%,光周期16 h光照、8 h黑暗,利用人工饲料饲养[13]。
供试菌株,2株球孢白僵菌CFCC81428和CFCC83116由北纳生物公司提供。利用马铃薯葡萄糖琼脂(PDA)培养基(配方:马铃薯200 g、葡萄糖20 g、琼脂15 g,蒸馏水1 L)培养。
中华甲虫蒲螨由河北省农林科学院昌黎果树研究所提供。置于直径2.0 cm、长10.0 cm的玻璃管内,使用人工气候箱培养,温度设置为(25±1)℃,相对湿度(50±5)%,用天牛幼虫供其寄生繁育[14]。
球孢白僵菌和中华甲虫蒲螨的毒(杀虫)力测定:将球孢白僵菌孢子接种在PDA固体平板培养基上,在25 ℃黑暗条件下培养15 d[15],将分生孢子溶于体积分数为0.05%的无菌聚山梨酯-80中,用无菌脱脂棉过滤掉菌丝,将孢子液稀释为105、106、107、108孢子/mL 4个孢子浓度梯度,菌株孢子萌发率均超过90%,用于毒力测定。选取冷杉梢斑螟的老熟幼虫,逐头浸入不同孢子浓度的球孢白僵菌分生孢子悬浮液中15 s后取出,用滤纸拭干虫体表面水分,放入养虫盒中,再盖上已扎多个直径1 cm小孔的养虫盒盖,置于人工气候箱中饲养,饲养条件设置为温度(25±1)℃,相对湿度(65±5)%,光周期16 h光照、8 h黑暗。每15头冷杉梢斑螟幼虫为1个重复,每个孢子浓度重复3次,以体积分数为0.05%的无菌聚山梨酯-80作为对照。连续观察记录10 d,每天记录幼虫死亡数,幼虫僵直不动视为死亡,计算出毒力数据[16],并将死虫挑出保湿培养,记录僵虫数。
将中华甲虫蒲螨在玻璃管中用天牛幼虫饲养,挑出5个数量梯度的雌螨[17],分别为20、40、60、80、100头,用于杀虫力测定,选取冷杉梢斑螟老熟幼虫,置于35 mm×12 mm的培养皿里,将不同数量梯度的雌螨挑取到培养皿里。每15头冷杉梢斑螟幼虫为1个重复,每个中华甲虫蒲螨数量设置3次重复,以未接种蒲螨处理的作为对照。连续观察3d,用针扎不动视为死亡,每天记录死亡数,计算得出半数致死量等杀虫力数据。
将球孢白僵菌和中华甲虫蒲螨联合使用,以孢子浓度为108孢子/mL的球孢白僵菌和数量为20、40、60、80、100头的中华甲虫蒲螨联合使用,以使用中华甲虫蒲螨和体积分数为0.05%的无菌聚山梨酯-80为对照,记录冷杉梢斑螟幼虫的死亡数;将数量为80头的中华甲虫蒲螨和孢子浓度为105、106、107、108孢子/mL的球孢白僵菌联合使用,以单独使用球孢白僵菌为对照,记录冷杉梢斑螟幼虫僵虫数。
虽然RBM模型有许多优势,但是现有的流行数值分析软件并没有RBM模型,使得应用RBM变得异常困难。使用基于有限元原理的Abaqus-6.11来进行对不同几何形状的直根的拔出模拟[1]。首先是用Abaqus找到一种可行的模拟根土相互作用的模型;其次将对不同树根模型进行拔出分析,得到几何模型的变化如何影响树根的拔出强度。
亚致死浓度(量)处理:根据球孢白僵菌和中华甲虫蒲螨对冷杉梢斑螟幼虫的致死情况,以球孢白僵菌6 d后的亚致死浓度(LC25)和中华甲虫蒲螨2 d后亚致死数量(LD25)对幼虫进行侵染和寄生处理,50头冷杉梢斑螟幼虫为一个重复,每个处理重复3次,以体积分数为0.05%的无菌聚山梨酯-80处理为对照,在12、24、36、48、60、72 h时对每组幼虫进行谷胱甘肽-S-转移酶(GSTs)、乙酰胆碱酯酶(AChE)、多酚氧化酶(PPO)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)的酶活力测定。
酶原制备方法:分别在菌和螨处理后的12、24、36、48、60、72 h进行取样,从处理过的每组幼虫中挑取活力较高的3头幼虫,分别加入1 mL提取液,进行冰浴匀浆。12 000 r/min、4 ℃离心15 min,取上清,置冰上待测。
GSTs、AChE、PPO、SOD、CAT活力测定:参照测试盒说明书(购自苏州格锐思生物科技有限公司)测定。
数据处理:根据试验所获数据计算各处理的死亡率、校正死亡率和僵虫率。
死亡率=(死亡虫数/总虫数)×100%。
校正死亡率=[(处理死亡率-对照死亡率)/(100%-对照死亡率)]×100%。
僵虫率=(僵虫数/总虫数)×100%。
2 结果与分析
2.1 球孢白僵菌对冷杉梢斑螟幼虫的侵染效果
2株球孢白僵菌对冷杉梢斑螟幼虫的室内毒力测定结果见表1。由表可知,2 d时CFCC83116菌株对幼虫的亚致死浓度(LC25)和半数致死浓度(LC50)分别为4.79×108、1.41×1011孢子/mL,CFCC81428菌株对幼虫的LC25和LC50分别为2.14×108、2.19×1010孢子/mL;6 d时幼虫死亡率上升明显,此时CFCC83116菌株对幼虫的LC25和LC50分别为6.92×106、1.05×109孢子/mL,CFCC81428菌株对幼虫的LC25和LC50分别为7.41×105、3.02×107孢子/mL;10 d时,CFCC83116菌株对幼虫的LC25和LC50分别为1.62×105、1.29×107孢子/mL,CFCC81428菌株对幼虫的LC25和LC50分别为2.04×104、7.24×105孢子/mL。2~10 d,球孢白僵菌CFCC81428的LC25、LC50均小于CFCC83116菌株。由表2可知,同一菌株随着孢子浓度的升高,其对冷杉梢斑螟幼虫的累计校正死亡率也逐渐升高。幼虫被孢子浓度为105、106孢子/mL的菌株CFCC83116侵染10 d后的累计校正死亡率,小于菌株CFCC81428侵染10 d后的累计校正死亡率;幼虫被孢子浓度为107、108孢子/mL的菌株CFCC83116侵染10 d后的累计校正死亡率,显著小于菌株CFCC81428侵染10 d后的累计校正死亡率(P<0.05)。由以上数据可知,菌株CFCC81428对冷杉梢斑螟幼虫的毒力大于菌株CFCC83116,故选取菌株CFCC81428做后续试验。

表1 不同饲养时间2株球孢白僵菌对冷杉梢斑螟幼虫的致死情况
CFCC81428菌株侵染冷杉梢斑螟幼虫并使其死亡后,在其表面产生菌丝,在体式显微镜下观察发现,冷杉梢斑螟染菌后48 h,虫体已有白色菌丝产生,分布在头部下侧和节间褶处,此时冷杉梢斑螟已经死亡,身体柔软(图1A);染菌后144 h,虫体被白色菌丝全部覆盖,营养耗尽,身体僵硬萎缩(图1B)。

A为接种后48 h的虫体形态;B为接种后144 h的虫体形态。

表2 冷杉梢斑螟幼虫被2株球孢白僵菌侵染10 d后的累计校正死亡率
2.2 中华甲虫蒲螨对冷杉梢斑螟幼虫的寄生效果
中华甲虫蒲螨对冷杉梢斑螟幼虫的室内杀虫力测定见表3。1 d时,蒲螨对幼虫的LD25和LD50分别为36.31、91.20头;2 d时,幼虫死亡率明显上升,蒲螨对幼虫的LD25和LD50分别为16.60、27.54头;3 d时,蒲螨对幼虫的LD25和LD50分别为16.60、25.12头。
冷杉梢斑螟幼虫被蒲螨寄生后,膨腹体逐渐变大。幼虫被寄生24 h后,大量蒲螨附着于虫体,此时虫体颜色加深,已被寄生致死(图2A);幼虫被寄生144 h后,蒲螨产生膨腹体,膨腹体上爬动着许多雄螨,等待子代雌螨从膨腹体中产出,与其交配,此时幼虫营养已经耗尽,虫体严重萎缩(图2B)。
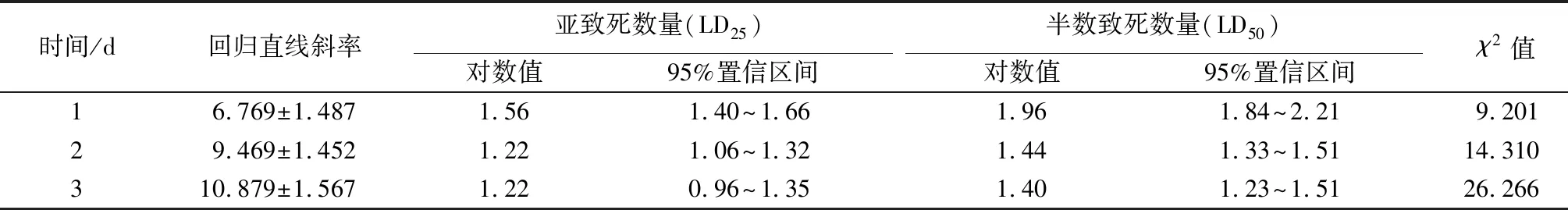
表3 不同饲养时间中华甲虫蒲螨对冷杉稍斑螟幼虫致死情况

A为寄生后24 h虫体形态;B为寄生后144 h虫体形态。
2.3 冷杉梢斑螟幼虫被球孢白僵菌和中华甲虫蒲螨联合处理后的作用效果
冷杉梢斑螟幼虫被中华甲虫蒲螨和球孢白僵菌联合处理3d后的累计校正死亡率见表4。由表可知,单独使用中华甲虫蒲螨20、40、60、80、100头时,对应的冷杉梢斑螟幼虫累计校正死亡率为51.11%、55.56%、80.00%、100.00%、100.00%。单独使用蒲螨寄生幼虫时,使用数量20、40头和使用数量60头的幼虫累计校正死亡率差异显著(P<0.05);使用数量60头和使用数量80、100头的幼虫累计校正死亡率差异显著(P<0.05)。说明冷杉梢斑螟幼虫的累计校正死亡率随着蒲螨数量的增加而增加。中华甲虫蒲螨80头和100头对应的冷杉梢斑螟幼虫累计校正死亡率差异不显著,说明冷杉梢斑螟幼虫被寄生超过80头蒲螨后,其累计校正死亡率不会显著增加。联合使用时,中华甲虫蒲螨20、40、60头对应的冷杉梢斑螟幼虫累计校正死亡率显著高于未联合使用的(P<0.05),说明中华甲虫蒲螨和球孢白僵菌联合使用对冷杉梢斑螟幼虫杀虫力更高。

表4 冷杉梢斑螟幼虫被球孢白僵菌和不同数量的中华甲虫蒲螨联合处理3 d后的累计校正死亡率
根据中华甲虫蒲螨对冷杉梢斑螟幼虫的致死情况,选取数量为80头的中华甲虫蒲螨和孢子浓度为105、106、107、108孢子/mL的球孢白僵菌联合使用,查看冷杉梢斑螟幼虫的僵虫率,以单独使用球孢白僵菌处理为对照,结果见表5。由表可知,冷杉梢斑螟幼虫被孢子浓度为105、106孢子/mL的球孢白僵菌和数量为80头的蒲螨联合处理后的僵虫率为22.22%和33.33%;冷杉梢斑螟幼虫被孢子浓度为105、106孢子/mL的球孢白僵菌单独侵染后的僵虫率为11.11%和17.78%。在球孢白僵菌孢子浓度为105、106孢子/mL时,联合处理与未联合处理冷杉梢斑螟幼虫的僵虫率差异显著(P<0.05)。冷杉梢斑螟幼虫被孢子浓度为107、108孢子/mL的球孢白僵菌和数量为80头的蒲螨联合处理后的僵虫率为35.56%和53.33%;幼虫被孢子浓度为107、108孢子/mL的球孢白僵菌单独侵染后的僵虫率为31.11%和48.89%。球孢白僵菌孢子浓度为107、108孢子/mL时,冷杉梢斑螟幼虫被联合处理的僵虫率均高于未联合处理。幼虫被孢子浓度为105、106孢子/mL的球孢白僵菌侵染后的僵虫率与被孢子浓度为107、108孢子/mL侵染后相比差异显著(P<0.05)。由表可知,同一处理下的冷杉梢斑螟幼虫僵虫率会随球孢白僵菌孢子浓度升高而不同程度的升高。

表5 冷杉梢斑螟幼虫被中华甲虫蒲螨和不同孢子浓度的球孢白僵菌联合处理后的僵虫率
中华甲虫蒲螨和球孢白僵菌单独或联合使用对冷杉梢斑螟幼虫谷胱甘肽-S-转移酶(GSTs)活性的影响见表6。由表可知,幼虫在被菌侵染12 h后GSTs活性均高于对照组,36 h时达到峰值,此时GSTs活性是对照组的3.6倍,显著高于对照组(P<0.05),说明单独使用球孢白僵菌侵染对冷杉梢斑螟幼虫GSTs活性有诱导作用;幼虫GSTs活性在被蒲螨寄生后12 h达到峰值,是对照组的1.2倍,被蒲螨寄生72 h后,GSTs活性低于对照组,处于被抑制状态,表明幼虫GSTs活性在被蒲螨寄生后的前60 h处于被诱导状态,72 h后处于被抑制状态;幼虫GSTs活性在被菌和螨联合处理后12 h达到峰值,是对照组的1.2倍,之后逐渐下降,前48 h,联合处理组高于对照组,幼虫GSTs活性处于被诱导状态,处理60 h后,联合处理组低于对照组,幼虫GSTs活性处于被抑制状态,幼虫在被菌和螨联合处理后,除48、72 h外,其他取样时间,GSTs活性均低于另外2个处理组,与另外2个处理组相比呈现进一步下降趋势。
中华甲虫蒲螨和球孢白僵菌单独或联合使用对冷杉梢斑螟幼虫乙酰胆碱酯酶(AChE)活性的影响见表7。由表可知,幼虫AChE活性在被菌侵染后36 h到达峰值,达到对照组的6.8倍,显著高于对照组(P<0.05),球孢白僵菌处理组幼虫AChE活性在所有取样时间均高于对照组,说明单独使用球孢白僵菌侵染对冷杉梢斑螟幼虫AChE活性具有诱导作用;幼虫AChE活性在被蒲螨寄生后12 h达到峰值,达到对照组的1.8倍,显著高于对照组(P<0.05),处于被诱导状态,被螨寄生后72 h时,幼虫AChE活性显著低于对照组(P<0.05),处于被抑制状态;幼虫AChE活性在被菌和螨联合处理后12 h时到达峰值,达到对照组的1.3倍,处于被诱导状态,48 h后,幼虫AChE活性低于对照组,处于被抑制状态,与另外2个处理组相比,AChE活性呈现进一步下降趋势。
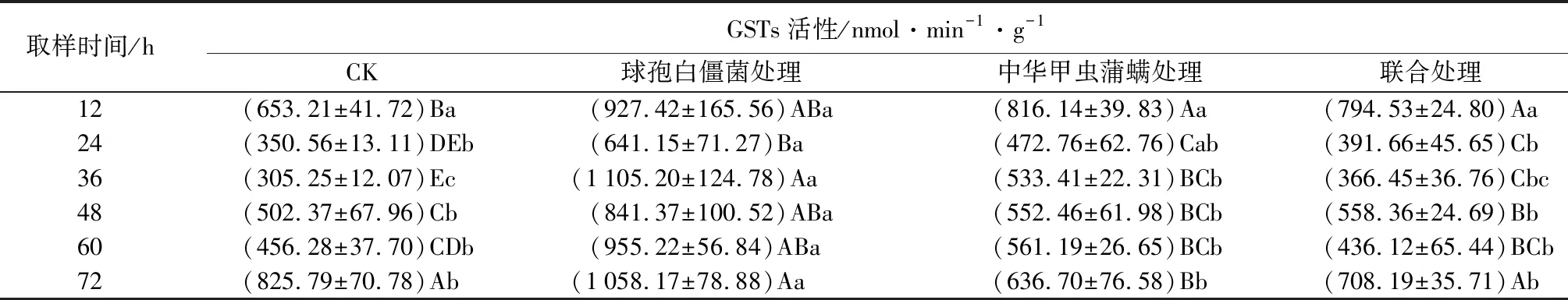
表6 亚致死浓度(数量)球孢白僵菌和中华甲虫蒲螨处理冷杉梢斑螟幼虫谷胱甘肽-S-转移酶(GSTs)的活性
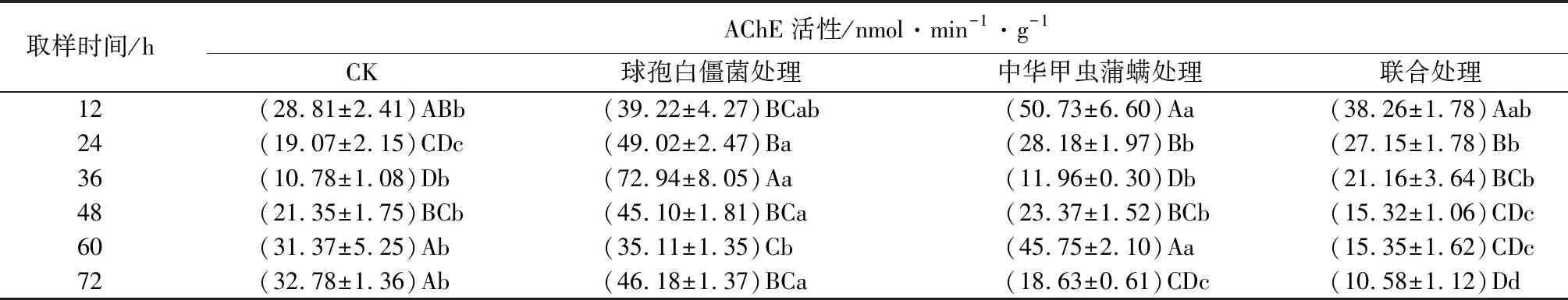
表7 亚致死浓度(数量)球孢白僵菌和中华甲虫蒲螨处理冷杉梢斑螟幼虫乙酰胆碱酯酶(AChE)活性
中华甲虫蒲螨和球孢白僵菌单独或联合使用对冷杉梢斑螟幼虫多酚氧化酶(PPO)活性的影响见表8。由表可知,幼虫PPO活性在被球孢白僵菌侵染12 h时低于对照组,处于未被激活状态,24 h时开始被激活,48 h时到达峰值,此时菌处理组PPO活性是对照组的2.4倍,显著高于对照组(P<0.05),幼虫PPO活性在被菌侵染24 h后处于被诱导状态;幼虫PPO活性在被中华甲虫蒲螨寄生后逐渐升高,48 h时到达峰值,达到对照组的1.5倍,显著高于对照组(P<0.05),PPO活性在被蒲螨寄生后的前60 h均高于对照组,处于被诱导状态,72 h后低于对照组,处于被抑制状态;幼虫PPO活性在被菌和螨联合处理后,仅24、48 h时高于对照组,其余时间均低于对照组,72 h时活性最低,显著低于对照组(P<0.05),证明此时抑制作用最明显。被菌和螨联合处理后,幼虫PPO活性与其他2个处理组相比呈现提前下降趋势,证明菌和螨联合使用对冷杉梢斑螟幼虫PPO活性抑制更强。
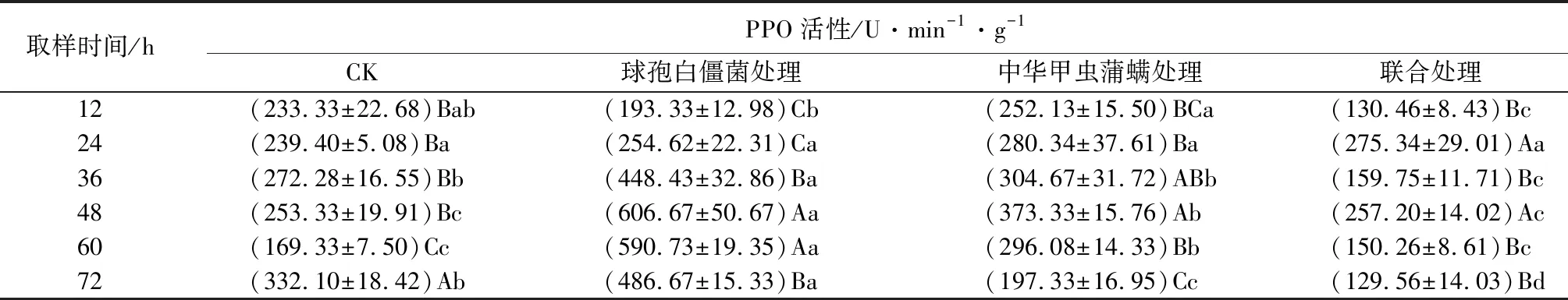
表8 亚致死浓度(数量)球孢白僵菌和中华甲虫蒲螨处理冷杉梢斑螟幼虫多酚氧化酶(PPO)活性
中华甲虫蒲螨和球孢白僵菌单独或联合使用对冷杉梢斑螟幼虫超氧化物歧化酶(SOD)活性的影响见表9。由表可知,幼虫SOD活性在被菌侵染36 h时到达峰值,是对照组的2.0倍,显著高于对照组(P<0.05),幼虫SOD活性12~72 h均处于被诱导状态;幼虫SOD活性在被蒲螨寄生前24 h低于对照组,从36 h开始被激活,36~72 h时蒲螨处理组SOD活性高于对照组,处于被诱导状态,幼虫SOD活性在被蒲螨寄生后36 h达到峰值,是对照组的1.5倍,显著高于对照组(P<0.05);在被菌和螨联合处理后,幼虫SOD活性始终低于对照组,证明菌和螨联合使用对冷杉梢斑螟幼虫SOD活性具有明显抑制作用。
中华甲虫蒲螨和球孢白僵菌单独或联合使用对冷杉梢斑螟幼虫过氧化氢酶(CAT)活性的影响见表10。由表可知,幼虫CAT活性在被菌侵染12 h时低于对照组,处于未被激活状态,在24 h开始被激活,36 h时到达峰值,达到对照组的1.9倍,显著高于对照组(P<0.05),幼虫CAT活性从24~72 h均高于对照组,处于被诱导状态;幼虫CAT活性在被蒲螨寄生前48 h高于对照组,处于被诱导阶段,36 h到达峰值,是对照组的2.1倍,显著高于对照组(P<0.05);幼虫CAT活性在被菌和螨联合处理后12 h到达峰值,是对照组的2.2倍,显著高于对照组(P<0.05),呈明显被诱导状态,72 h时低于对照组,处于被抑制状态。幼虫CAT活性与其他2个处理组相比呈现提前下降趋势。
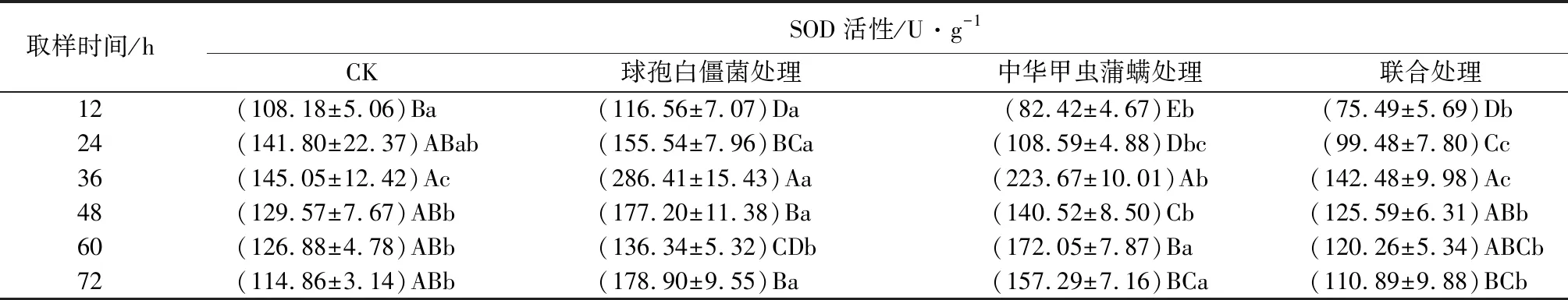
表9 亚致死浓度(数量)球孢白僵菌和中华甲虫蒲螨处理冷杉梢斑螟幼虫超氧化物歧化酶(SOD)活性

表10 亚致死浓度(量)球孢白僵菌和中华甲虫蒲螨处理冷杉梢斑螟幼虫过氧化氢酶(CAT)活性
3 结论与讨论
随着冷杉梢斑螟危害的加重,生物防治方法将会作为一种可靠且能与其他方法兼容的措施,在综合害虫防治中具有重要地位[11]。球孢白僵菌作为一种非常重要的昆虫病原真菌,对梢斑螟具有较高的毒力和防治潜力[18]。本研究从2株球孢白僵菌中筛选发现,CFCC81428菌株在孢子浓度为108孢子/mL时,侵染冷杉梢斑螟幼虫10 d时累计校正死亡率为77.30%、半数致死浓度为7.24×105孢子/mL,表明CFCC81428菌株毒性更强,故选取此菌株做后续试验。
中华甲虫蒲螨作为我国本土的一种寄生性害虫天敌,对鳞翅目昆虫表现出了较高的杀虫效果[19],将其与球孢白僵菌联合起来应用,可提升防治梢斑螟的效果,同时尽可能降低病原菌和天敌的用量,为本文拟探索的重点内容。本研究发现,中华甲虫蒲螨和球孢白僵菌联合处理组对冷杉梢斑螟幼虫的累计校正死亡率高于中华甲虫蒲螨处理组;联合处理组对冷杉梢斑螟幼虫的僵虫率也高于球孢白僵菌处理组。这是由于中华甲虫蒲螨寄生幼虫时,使幼虫体表产生了伤口,使球孢白僵菌更易侵染,导致其僵虫率更高。在实验室条件下,中华甲虫蒲螨的寄生,对冷杉梢斑螟幼虫的致死率可以达到100%。但这只是在实验室封闭条件下实现的,由于天气、温度等因素影响,在野外要达到室内条件下的杀虫效果,对其释放技术还有待进一步探索。
在昆虫体内,AChE和GSTs是2种重要的解毒酶,能够分解植物次生代谢物、真菌、细菌等产生的外源毒素[20],维持昆虫正常生理生化活动,提高昆虫的抗逆能力,在细胞的许多重要代谢过程中起着重要作用[21-24]。本研究发现,冷杉梢斑螟幼虫AChE和GSTs活性在被亚致死浓度下的球孢白僵菌侵染后36 h时到达峰值,其余各取样时间也均高于对照组,表现为被诱导状态。幼虫AChE和GSTs活性在被亚致死量下的中华甲虫蒲螨寄生后12 h到达峰值,然后逐渐下降,到72 h时低于对照组,处于被抑制状态。这是因为中华甲虫蒲螨产生毒素,导致冷杉梢斑螟幼虫难以维持正常的生理生化活动,生命力下降,从而解毒酶活性受到抑制。球孢白僵菌和中华甲虫蒲螨联合使用时,幼虫解毒酶活性与其他2个处理组相比呈现进一步下降趋势,这是由于中华甲虫蒲螨和球孢白僵菌的联合处理,产生了更多的毒素,幼虫酶活性受到抑制,从而导致幼虫解毒酶活性与其他2个处理组相比呈现进一步下降趋势。
当昆虫受到外源逆境胁迫时,PPO、SOD、CAT这3种保护酶能迅速做出应激反应,从而维持正常的生理生化活动[25-27]。保护酶可以维持一定的自由基水平,从而防止自由基对生物体的危害,一旦保护酶系遭到破坏,导致生物机体内氧自由基浓度过高,将破坏生物功能分子,使细胞功能受到威胁[28]。本研究发现,冷杉梢斑螟幼虫PPO、SOD、CAT这3种保护酶中,SOD和CAT活性在被球孢白僵菌侵染后36 h到达峰值,PPO活性在被侵染后48 h到达峰值,在被处理后的12 h,PPO和CAT活性略低于对照组,这是由于球孢白僵菌尚未侵染到冷杉梢斑螟幼虫体内,其活性还未被激活。其他各取样时间点,幼虫保护酶活性均高于对照组,表现出明显的被诱导状态。单独使用中华甲虫蒲螨处理后,幼虫的3种保护酶活性均呈现先升高再下降的趋势,这是由于中华甲虫蒲螨寄生幼虫后,产生的毒素导致其保护酶活性增加,但随着寄生时间的延长,幼虫生理功能受到破坏,保护酶活性也逐渐降低。幼虫SOD和PPO活性,被球孢白僵菌和中华甲虫蒲螨联合处理后,与其他2个处理组相比呈现提前下降趋势,这是由于中华甲虫蒲螨的寄生,导致冷杉梢斑螟幼虫体表产生伤口,加速了球孢白僵菌的侵染,导致幼虫SOD和PPO活性与另外2个处理组相比呈现提前下降趋势。
根据对冷杉梢斑螟幼虫的毒力测定,试验筛选出了对冷杉梢斑螟幼虫高效的球孢白僵菌菌株,通过对球孢白僵菌和中华甲虫蒲螨的联合试验,证明了两者联合使用的增效作用。根据幼虫解毒酶和保护酶活性的变化发现,病原菌开始侵染或天敌开始寄生时,幼虫为了维持其正常的生理活动,抵御外源毒素影响,从而使其解毒酶和保护酶活性升高,但随着侵染(或寄生)时间的延长,幼虫防卫能力逐渐瓦解,表现为解毒酶和保护酶活性下降,随着毒素的积累,细胞受到破坏,幼虫将无法正常活动最终导致死亡。
猜你喜欢
环境昆虫学报(2023年5期)2023-11-09 12:31:52
山西林业科技(2022年4期)2022-03-12 04:39:48
中国生物防治学报(2021年3期)2021-08-11 02:15:38
湖北林业科技(2019年2期)2019-05-05 05:36:12
花火A(2019年1期)2019-03-22 02:27:42
现代养生·上半月(2017年7期)2017-07-07 21:43:25
中成药(2017年5期)2017-06-13 13:01:12
上海农业学报(2016年2期)2016-10-27 00:49:41
广西林业科学(2016年4期)2016-03-16 05:44:50
生物灾害科学(2015年3期)2015-03-11 16:38:38
