
翘嘴鲌淀山湖和兴凯湖群体的胚胎发育及染色体核型比较
2023-05-23 03:28:04徐博文张国奇周陆张友良王军王成辉
水产科技情报 2023年3期
徐博文 张国奇 周陆 张友良 王军 王成辉
(1 上海海洋大学,农业农村部淡水水产种质资源重点实验室,水产科学国家级实验教学示范中心,上海水产养殖工程技术研究中心,上海 201306; 2 上海松江水产 良种场有限公司,上海 201616; 3 上海市松江区水产技术推广站,上海 201616)
翘嘴鲌(Culteralburnus)隶属于鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲌亚科(Cultrinae)、鲌属(Culter),是我国重要的经济鱼类,在水产养殖中占有重要的地位[1]。翘嘴鲌具有体形好、广温性、生长快及肉质鲜美等优良性状和较高的经济价值[2],深受养殖户和消费者喜爱。翘嘴鲌在我国分布范围广,从我国的台湾、珠江流域、长江干流、长江支流(淮河)一直到黑龙江流域等几大水系均有分布。翘嘴鲌不同的地理群体具有不同的种质遗传特性。史建伍等[3]基于随机扩增多态性(RAPD)方法对鄱阳湖、梁子湖以及太湖的3个不同翘嘴鲌地理群体进行了遗传多样性分析,发现鄱阳湖群体与梁子湖群体亲缘关系较近,与太湖群体稍远。伊西庆[4]用线粒体ND2基因分析了兴凯湖与5个翘嘴鲌地理群体间的遗传差异,结果表明,兴凯湖群体与其他群体间存在极显著的遗传分化(P<0.01)。王伟[5]应用扩增片段长度多态性(AFLP)方法、内部简单重复序列(ISSR)核DNA分子标记和线粒体COII、D-LOOP标记对我国太湖、兴凯湖、梁子湖、浮桥河水库、三溪口水库、南湾水库以及湖州人工养殖池塘等7个不同地理群体翘嘴鲌进行了遗传多样性比较分析,结果发现,兴凯湖群体、南湾水库群体和太湖群体的遗传多样性要高于其他群体,并根据各群体遗传特征呈现的差异性,首次提出将翘嘴鲌分为南、北两大区系。
我国在翘嘴鲌胚胎发育研究方面已有不少报道。顾志敏等[6]对太湖翘嘴鲌进行了胚胎及胚后发育观察,指出翘嘴鲌受精卵为圆球型,呈墨绿色、青灰色、黄色3种颜色,为黏性卵,并且对其胚胎发育特征进行了详细的描述。刘丹阳等[7]对兴凯湖翘嘴鲌的胚胎发育进行了观察,指出其受精卵呈淡黄绿色或青灰色,为半浮性卵,无黏性。上述两个研究表明,由于地理隔离,北方的兴凯湖群体与南方的长江流域群体,其受精卵呈现出了不同特性。然而,翘嘴鲌南、北方群体的受精卵差异是否是由于长期的环境适应性演化造成的,还有待进一步探讨。研究翘嘴鲌南、北方群体在同一环境下的受精卵和繁殖特性,将为后续翘嘴鲌的人工养殖以及遗传育种提供重要参考。
核型是指一个细胞内有丝分裂中期所有染色体的表型,如染色体数目、大小和形态特征等。核型分析研究不仅可以在分类系统中探讨物种地位,了解系统的演化过程,还可以为一些重要养殖种类的杂交育种及分子生物学研究等提供理论依据和实践指导[8]。按照Levan等[9]的分类标准,染色体可分为中着丝粒染色体(m)、亚中着丝粒染色体(sm)、端部着丝粒染色体(t)、亚端着丝粒染色体(st)4种类型。国内对翘嘴鲌染色体的核型分析已有一些报道,但存在较大差异。如王晓艳等[10]发现,洞庭湖翘嘴鲌染色体数目为2n=48(n为配子体的染色体个数),核型公式为18m+10sm+18st+2t,染色体总臂数(NF)为76条;杨晓芬等[11]报道的贵州地区翘嘴鲌的核型公式则为16m+26sm+6st,NF=90;而赵春霞等[12]研究发现,兴凯湖翘嘴鲌染色体数目为2n=48,核型公式为16m+16sm+12st。翘嘴鲌南、北方群体在染色体数目和核型上的较大差异需要进一步的研究和确认。
本研究对来自南方淀山湖以及北方兴凯湖的翘嘴鲌群体的胚胎发育过程进行了系统的观察比较,并通过染色体核型分析探究两个群体的遗传差异,旨在了解分布在南、北方不同区域翘嘴鲌群体间的变异程度及其繁育的环境适应性,为翘嘴鲌种质资源的进一步挖掘和利用提供必要的基础数据。
1 材料和方法
1.1 试验材料
翘嘴鲌北方群体样本采自黑龙江兴凯湖,南方群体样本采自长江水系(上海淀山湖)。两个群体的亲鱼培育均在上海市松江水产良种场进行,管理方法一致。本试验于2020年5月进行翘嘴鲌南方淀山湖群体、北方兴凯湖群体的人工繁育。
1.2 胚胎发育
1.2.1 授精与孵化条件
挑选性腺发育良好的亲鱼进行人工催产。雌性亲鱼注射促黄体生成素释放激素类似物LRH-A(15 U/kg)和地欧酮(1.5 U/kg);雄性亲鱼剂量减半。在注射催产剂12 h后,采用干法人工授精,然后将受精卵平铺于培养皿中进行孵化。通过温度计实时监控水温变化情况。
1.2.2 胚胎发育观察
参考王云等[13]的方法,在显微镜下观察胚胎各发育时期的特征,当半数胚胎发育至某时间点时作为该期的胚胎发育时间,记录并拍照。
1.2.3 繁育相关指标的统计分析
翘嘴鲌南方淀山湖群体、北方兴凯湖群体各取3个生物学重复,统计它们的相对繁殖力、卵径大小、受精率和出膜后体长。相对繁殖力采用李思发[14]的方法,即相对繁殖力=卵粒总和(粒)/体质量(g);受精率为原肠期正常受精卵数与总卵数之比[13];卵径和出膜后体长均在显微镜下观察并测量。相关数据用(平均值±标准差)表示,利用SPSS 21.0软件进行t检验统计学分析。
1.3 核型分析
1.3.1 染色体制备
参照孙远东等[15]的方法,在试验鱼的胸鳍基部注射植物血凝素(PHA)溶液(10 μg/g);12 h后在同一部位再次注射PHA(15 μg/g);作用3 h后,在相同部位第3次注射PHA(10 μg/g),同时在另一侧胸鳍基部注射秋水仙素(colchicine)(4~6 μg/g)。1.5~2.0 h后,断鱼尾放血2~3 min,随后取鱼肾脏,用手术剪剪碎后转移至10 mL离心管中,猛烈吹打制成细胞悬液;离心后收集细胞,弃上清液,加入KCl溶液(0.075 mol/L)低渗处理,随后加入新鲜配制的卡诺固定液(甲醇与冰醋酸体积比3∶1,并提前-20 ℃预冷);取预冷的干净载玻片,从载玻片高处进行滴片,之后将载玻片在酒精灯上过火15~20次,再用Giemsa染色液染色1~2 h,最后封片,镜检。
1.3.2 染色体观察与核型编排
在光学显微镜下观察翘嘴鲌南方淀山湖群体、北方兴凯湖群体的染色体形态,记录并拍照。使用Photoshop软件对图片进行观察和测量,首先确定着丝粒的位置,然后通过软件自带的测量功能,测量每条染色体着丝粒到染色体臂末端的距离,得到染色体的相对臂长。按照Levan等[9]的染色体分类标准,根据臂比值(长臂臂长/短臂臂长)将染色体分为m、sm、t、st共4组,并根据染色体长度及臂比值对其进行两两配对、编号并排序。
2 结果
2.1 翘嘴鲌南方淀山湖群体、北方兴凯湖群体的繁育性能比较
翘嘴鲌南方淀山湖群体、北方兴凯湖群体的相对繁殖力、卵径、受精率、出膜后体长情况见图1。南方淀山湖群体的相对繁殖力(相对怀卵率)略高于北方兴凯湖群体,但差异不显著(P>0.05),而北方兴凯湖群体的卵径为(1.21±0.03)mm,极显著大于南方淀山湖群体[(1.11±0.05)mm](P<0.01)。北方兴凯湖群体的受精率为96.8%,略高于南方淀山湖群体的94.5%,但差异不显著(P>0.05)。翘嘴鲌北方兴凯湖群体仔鱼的出膜后体长为(5.77±0.29)mm,极显著大于南方淀山湖群体[(3.40±0.14)mm](P<0.01)。
2.2 翘嘴鲌南方淀山湖群体、北方兴凯湖群体的胚胎发育特征和时序比较
2.2.1 受精卵
翘嘴鲌南方淀山湖群体的受精卵黏性很强,卵膜较厚,不易剥离,呈灰黄色,吸水后膨胀不明显。北方兴凯湖群体的受精卵无黏性,为浮性卵,卵膜较薄,呈青灰色,吸水后体积膨胀明显。两个群体的胚胎发育特征和时序见表1、图2。
2.2.2 胚盘期
卵内出现帽状隆起,形成胚盘。南方淀山湖、北方兴凯湖群体在此期的发育时序和形态均无较大差异,但南方淀山湖群体进入胚盘期的时间相对较短。
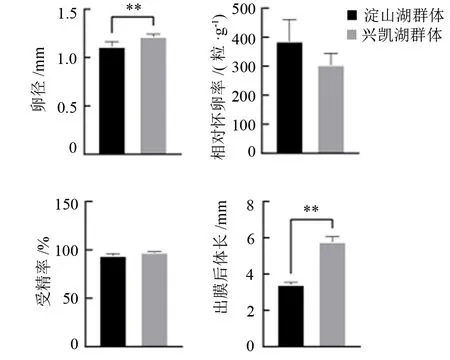
注:**表示组间差异极显著(P<0.01)。
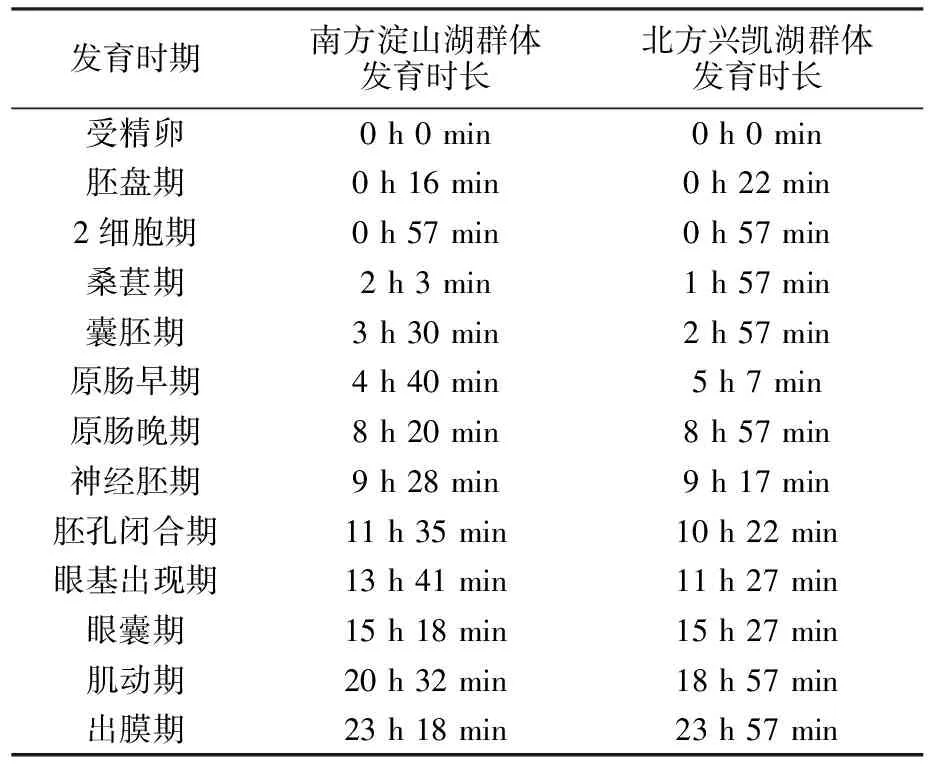
表1 水温(25±1)℃条件下翘嘴鲌南方淀山湖、北方兴凯湖群体的胚胎发育时序
2.2.3 细胞期
胚盘逐渐发育出分裂沟,胚胎不断地以倍数分裂,此为多细胞期。此后,受精卵继续经过数次卵裂形成大量紧密的细胞团,呈桑葚状,进入桑葚期。南方淀山湖、北方兴凯湖群体进入桑葚期的历时和形态特征无明显差异。
2.2.4 囊胚期
南方淀山湖、北方兴凯湖群体在此期的发育胚胎外部特征相似,在受精4 h后形成囊胚腔,囊胚层向下逐渐延伸,分裂球之间已无明显界限。囊胚层继续向下延展,1/3的胚胎被其包裹。但南方淀山湖群体较北方兴凯湖群体进入囊胚期的历时更短。

注:1. 受精卵;2. 胚盘期;3. 2细胞期;4. 4细胞期;5. 8细胞期;6. 16细胞期;7. 32细胞期;8. 桑葚期;9. 囊胚早期;10. 囊胚中期;11. 囊胚晚期;12. 原肠早期;13. 原肠中期;14. 原肠晚期;15. 神经胚期;16. 胚孔闭合期;17. 眼基出现期;18. 眼囊期;19. 尾芽期;20. 晶体出现期;21. 肌动期;22. 出膜前期;23. 出膜期;24. 1日龄。
2.2.5 原肠期
出现新月状的胚环,囊胚层继续向下延展,直至包裹住2/3的胚胎,进入原肠中期。囊胚层继续向下延展至胚胎的3/4,囊胚层开始增厚。形态上南方淀山湖、北方兴凯湖群体的发育胚胎外部特征无异,而南方淀山湖群体在原肠期发育开始变缓,发育时长较北方兴凯湖群体胚胎有所增加。
2.2.6 神经胚期
南方淀山湖、北方兴凯湖群体的发育胚胎外部特征无明显差异,胚层下包至4/5处,逐渐形成卵黄栓。胚环前段不断变大形成脑泡,此为神经胚期。南方淀山湖群体胚胎从原肠期至神经胚期发育均较为缓慢。
2.2.7 胚孔闭合期
随着胚层进一步下包,卵黄变成不规则圆形,外露渐小,最后卵黄栓消失,在卵黄栓末端形成圆形有凹陷的胚孔。胚孔逐渐变小直至封闭。此时期南方淀山湖、北方兴凯湖群体的胚胎在形态特征上相似,发育时序也大致相同,但南方淀山湖群体胚胎发育开始加快。
2.2.8 眼囊期
眼囊期又称视囊出现期,胚环前段变大,视泡突出呈椭圆形;尾部逐渐变细长,肌节逐渐发育出现。此时期两个群体胚胎形态特征一致,但北方兴凯湖群体的历时较长。
2.2.9 肌动期
胚体开始出现一定频率的收缩,且尾部在不断摆动。南方淀山湖、北方兴凯湖群体胚胎外部特征相近,南方淀山湖群体的胚胎发育略慢于北方兴凯湖群体。
2.2.10 出膜期
胚胎外膜破裂,仔鱼开始出膜。仔鱼呈透明状,鱼体前端具有一个梭状的卵黄膜以供营养需求。此时仔鱼发育出部分尾鳍,能间歇性地进行上下垂直游动。形态上,南方淀山湖、北方兴凯湖群体发育特征相似,北方兴凯湖群体的发育时序略短于南方淀山湖群体,但差异不明显。北方兴凯湖群体仔鱼的出膜后体长极显著大于南方淀山湖群体(P<0.01)。
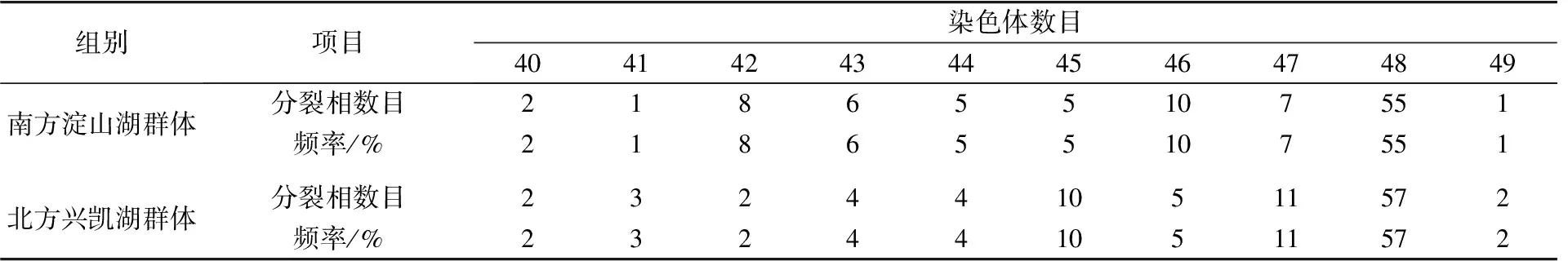
表2 翘嘴鲌南方淀山湖、北方兴凯湖群体的染色体数目分布
2.3 染色体数目与核型
从翘嘴鲌南方淀山湖、北方兴凯湖群体各挑选100个良好的染色体分裂相进行显微镜观察计数,统计染色体数目。结果显示,南方淀山湖群体中染色体为2n=48的占55%,北方兴凯湖群体中染色体为2n=48的占57%(见图3、表2)。
通过染色体臂比计算,发现m组臂比值为1.0~1.7,为中着丝粒染色体,南方淀山湖群体共有12对,北方兴凯湖群体共有11对;sm组为亚中着丝粒染色体,臂比值为1.7~3.0,南方淀山湖群体共有10对,北方兴凯湖群体共有13对;st组臂比值范围在3.0~7.0,为亚端着丝粒染色体,南方淀山湖群体共有2对。翘嘴鲌南方淀山湖群体的核型公式为2n=24m+20sm+4st,NF=88;翘嘴鲌北方兴凯湖群体的核型公式为2n=22m+26sm,NF=96(见图3、表3)。
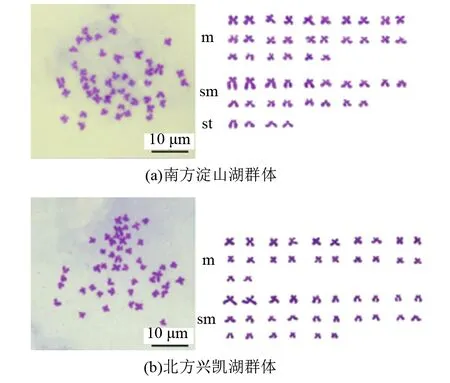
注:m为中着丝粒染色体,sm为亚中着丝粒染色体,st为亚端着丝粒染色体。
3 讨论
由于生存的地理环境差异,物种通常会形成具有环境适应性特征的地理群体[16-18]。据研究报道,南、北方的翘嘴鲌群体具有生长和繁殖习性的差异,这可能与其环境适应性密切相关[19]。本研究将翘嘴鲌北方兴凯湖群体引入南方水体进行培育,并比较了两者在相同养殖环境下繁育性能的差异,结果表明,翘嘴鲌南方淀山湖、北方兴凯湖群体的受精卵存在较明显的形态和结构差异。刘丹阳等[7]认为,在人工养殖环境下,水体的理化因子、饵料来源和营养配比的不同会对翘嘴鲌亲鱼产生影响,进而导致其受精卵的差异。然而,本研究中的北方兴凯湖群体是在上海松江地区通过人工繁育获得的,其亲鱼培育、胚胎发育的环境与南方淀山湖群体基本一致。之前的研究也表明,来自北方兴凯湖的翘嘴鲌,其受精卵为非黏性卵,与本研究的北方兴凯湖翘嘴鲌受精卵特征一致[20]。这说明北方兴凯湖翘嘴鲌与南方淀山湖翘嘴鲌受精卵形态和结构的差异并不是人工养殖环境影响所致,可能是长期的地理隔离形成的适应性演化[21]。本研究还发现,翘嘴鲌北方兴凯湖群体的卵径和出膜后的仔鱼体长都显著大于南方淀山湖群体。有研究认为,卵径是反映生物生活史进化过程的一个重要的生物学特征,其大小与环境相关,是鱼类长期适应环境的结果[22]。据此推测,翘嘴鲌北方兴凯湖群体的繁殖特性是其长期适应兴凯湖的地理环境而形成的适应性特征,是一种稳定遗传的特征。本研究中,翘嘴鲌北方兴凯湖群体在上海松江区水产良种场进行繁育时仍具有较高的相对繁殖力和受精率,说明其可以在南方水域进行正常繁育。
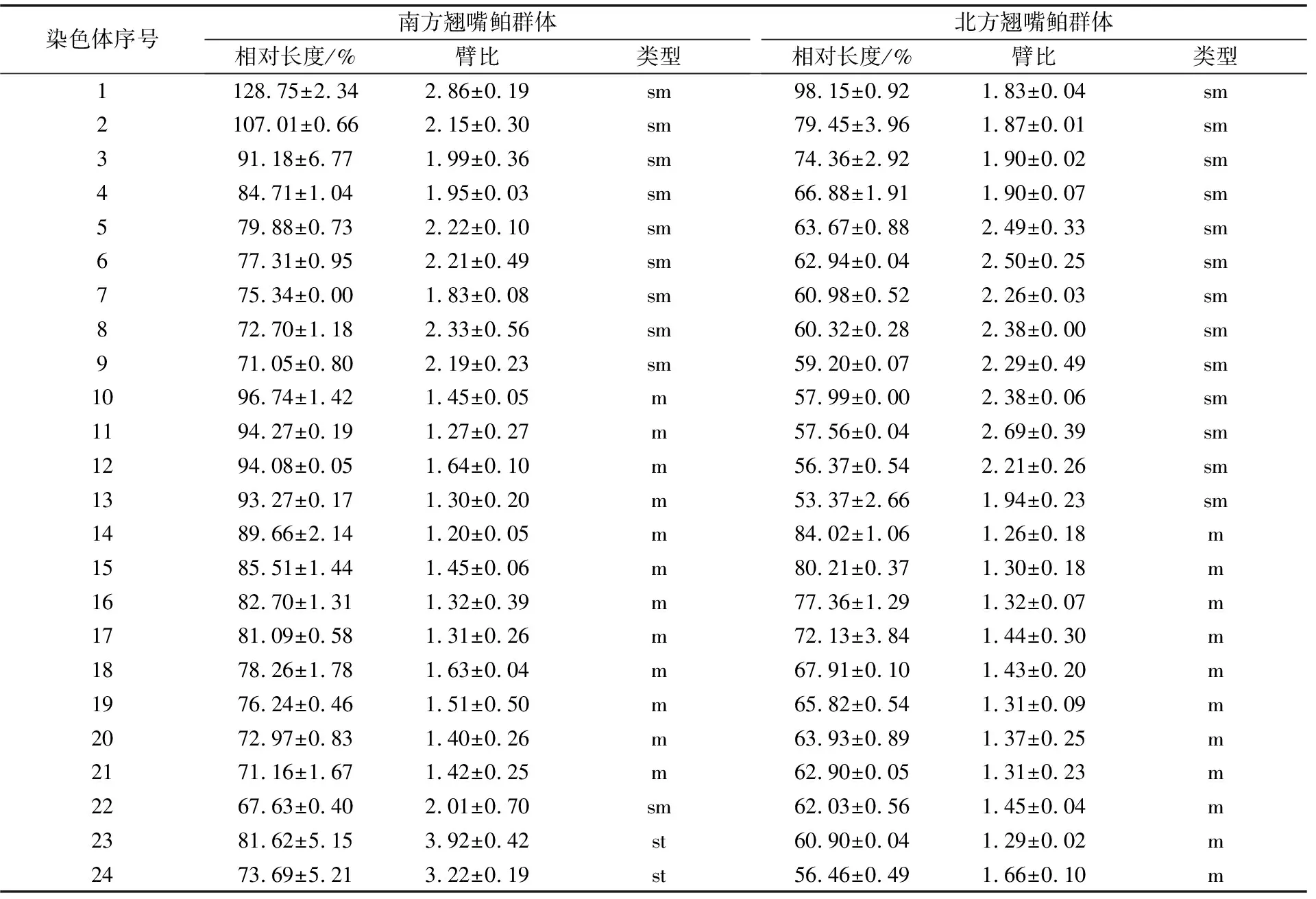
表3 翘嘴鲌南方淀山湖、北方兴凯湖群体核型分析参数(n=4)
染色体是遗传物质的载体,每个物种都具有独特的染色体组成。鱼类的染色体核型类型与其进化存在一定的关系[23]。通过对翘嘴鲌南方淀山湖、北方兴凯湖群体的染色体核型进行分析可知,南方淀山湖、北方兴凯湖群体的染色体数目均为2n=48,与国内绝大多数已有的关于翘嘴鲌染色体的报道一致,但与赵春霞等[12]报道的兴凯湖翘嘴鲌染色体数目(2n=44)有所差异。同种鱼染色体数目不一致的现象在大黄鱼(Larimichthyscrocea)[24]、马口鱼(Opsariichthysbidens)[25]、花尾胡椒鲷(Plectorhinchuscinctus)[26]、鲫(Carassiusauratus)[27]中均有存在。但本研究的结果支持北方兴凯湖翘嘴鲌的染色体数为2n=48。虽然翘嘴鲌南方淀山湖、北方兴凯湖群体的染色体数目均为48条,但其核型存在一定的差异,而此差异同样可能是长期的地理隔离形成的遗传变异造成的。导致染色体出现差异的原因可能有以下几点:(1)不同地理种群在长期的自然选择条件下产生了适应性演化;(2)种内核型多态化现象可能是群体的遗传多样性所致[28];(3)试验操作方法,如PHA和秋水仙素的用量和处理时长均会对染色体的形态造成一定的影响。尽管翘嘴鲌南方淀山湖、北方兴凯湖群体的染色体核型存在一定的差异,但整体而言,翘嘴鲌南方淀山湖、北方兴凯湖群体应属于同一物种的不同地理种群。
综上所述,虽然翘嘴鲌南方淀山湖群体、北方兴凯湖群体由于长期的地理隔离,种群间可能出现了一定的适应性演化,造成其繁殖性能,如卵的特性、相对繁殖力、卵径、出膜仔鱼的体长等有一定的差异,但北方兴凯湖翘嘴鲌能够在南方如上海地区正常繁育,其相对繁殖力和受精率等重要繁育指标仍具有较好的表现,并不影响其在南方水体的繁育实践。关于翘嘴鲌南方淀山湖群体、北方兴凯湖群体的分子遗传差异仍有待进一步的深入研究。
猜你喜欢
现代商业银行·财富生活(2023年12期)2024-01-20 12:16:33
生物技术进展(2022年1期)2022-02-11 03:45:46
地质与资源(2021年1期)2021-05-22 01:24:24
科学之谜(2018年9期)2018-12-17 09:05:08
中国诗歌(2016年6期)2016-11-25 23:54:04
生物学教学(2016年12期)2016-04-10 23:04:28
华人时刊(2016年3期)2016-04-05 05:56:28
雪花(2015年6期)2015-12-10 23:48:35
遗传(2015年4期)2015-02-04 06:29:22
城市建设理论研究(2014年11期)2014-04-21 00:36:00
