
新疆尉犁地区两种沙蜥的巢址特征比较
2023-05-16 03:06:34钟文赵伟祁玥陈伟基李铀
野生动物学报 2023年2期
钟文,赵伟,祁玥,陈伟基 ,李铀,3
(1.西北民族大学生命科学与工程学院,兰州,730030;2.兰州大学生命科学学院,甘肃省环境生物监测与修复重点实验室,兰州,730030;3.西北民族大学生物医学研究中心,甘肃省动物细胞技术创新中心,兰州,730030)
为适应不同环境,不同种群具有不同的生长率、死亡率、初次性成熟年龄和繁殖力,从而形成各自的生活史对策。自然选择有利于哪一类型生活史对策的形成,取决于各种生物所生活的栖息地特征[1-2]。巢穴是栖息地特征中重要的决定因素之一。巢对动物育幼、躲避敌害、休息和保持体温等具有重要的生态意义[3−5],巢址选择是在长期进化过程中形成的一种繁殖与防御对策,是与环境协同进化的结果[6],合适的巢址是动物繁衍生息的重要环节之一[7−9]。Calderón-Espinosa等[10]使用生物统计方法研究表明巢内土壤湿度、巢倾角和温度等生境因子会影响安第斯高原蜥蜴(Anadia bogotensis)选择合适的巢址。Tiatragul等[11]比较了美洲安乐蜥(Anolisspp.)2种不同栖息地之间的巢穴特征并描述差异。Muller等[12]研究发现雌性怀孕的疣尾蜥虎(Hemidactylus frena⁃tus)独居和群居时会选择不同的筑巢地点。目前国内学者对鸟类[6−7,9,13−17]和哺乳类[4−5,18−20]众多物种都进行了详尽的巢址研究,而爬行类中则仅有绿海龟(Chelonia mydas)、吐鲁番沙虎(Teratoscincus ro⁃borowskii)、丽斑麻蜥(Eremias argus)及青海沙蜥(Phrynocephalus vlangalii)几个物种有限的巢址研究[8,21−24]。
叶城沙蜥(Ph.axillaris)和南疆沙蜥(Ph.for⁃sythii)均隶属于鬣蜥科(Agamidae)沙蜥属,是我国特有的2种小型爬行动物。南疆沙蜥为卵胎生,主要分布于新疆维吾尔自治区天山山脉以南、海拔 1 400~3 100 m 的广大地区[25]。叶城沙蜥为卵生,主要分布于新疆天山山脉以南地区,由西部的喀什沿塔里木盆地的北缘经南湖戈壁,东抵敦煌、安西和酒泉等河西走廊西段地区;往南经叶城、和田、若羌与北部分布区连接,并伸入更南的阿尔金山昆仑山地区。垂直分布高度可自接近海平面至1 500 m,在阿尔金山南麓甚至高达3 000 m 左右[25]。目前针对南疆沙蜥主要集中于基因核型[26]、细胞学研究[27]、系统地理结构[28−29]、线粒体基因组特征[30−32]、微卫星位点开发[33−34]和遗传多样性及遗传结构的相关研究[35−36],而生活史方面的研究十分罕见。叶城沙蜥除了分类和分布区域调查外[37],仅见线粒体DNA 基因序列及遗传结构的相关研究[29,34,38−39]。关于2种沙蜥巢址的研究尚未见详细报道。
因此,为了解2种沙蜥的选巢特点,本研究首次从巢穴温度、湿度、深度、巢口倾角和巢口方向5 个指标揭示二者巢址选择的区别,以期为后续巢址研究奠定基础。
1 研究区概况
尉犁县位于新疆中部巴音郭楞蒙古自治州腹地(40°10′—41°39′ N,84°2′—89°58′ E)。该县年平均气温10.6 ℃,年降水量44.6 mm,年蒸发量2 856.8 mm[40]。气候属暖温带大陆性荒漠气候,昼夜温差较大,夏季炎热,冬季干冷,极高气温为42.2 ℃,极低气温可达-31.0 ℃;最大风力12 级,多出现在4 月,年平均干热风12 d,多出现在6 月中旬,是全国干热风严重地区之一[41]。尉犁县境内植被丰富,天然植物主要有胡杨(Populus euphratica)、灰胡杨(P.pruinosa)、柽柳(Tamarix chinensis)、胀果甘草(Glycyrrhiza inflata)和罗布麻(Apocynum venetum)等[42]。
2 研究方法
2.1 调查方法
于2019 年6 月26 日—7 月5 日在尉犁进行为期10 d的野外调查,调查期间最高气温38 ℃,最低气温19 ℃,10 d 平均气温28 ℃。通过徒步追踪南疆沙蜥和叶城沙蜥,观察蜥蜴巢穴位置并在其附近的灌木丛或其他植物上用彩带标记,同时利用手持GPS(GARMIN Etrex 229x)为每个巢穴定位。以含5~10个活跃巢穴为标准,设置200 m×200 m 的样方。测量和记录样方内巢穴的温度、湿度、深度、巢口倾角及巢口方向5 个指标。其中,温度使用数字式测温仪(UNI-T UT325)测量,测量时在确保不破坏巢穴的情况下,将测温仪伸入巢穴底部,以巢穴底部温度记为该巢穴温度;湿度通过探针式土壤检测仪(麦德牛MDN-1874)测量,与温度测量类似,将检测仪探针伸入巢穴底部记录湿度指标(以湿润WET、极湿润WET+、中性NOR、干旱DRY 及极干旱DRY+表示);利用卷尺测量巢穴深度(cm);倾角仪(途牛IP65)测量巢口倾角(水平为0°,向上倾斜为正值,向下倾斜为负值);巢口方向用指南针测定(以东、南、西、北、东南、东北、西南及西北表示)。样方数据采集于每日(晴天)09:30—11:30,所有测量工作尽量在2 h内完成。
2.2 数据处理
对2种沙蜥的3 个连续性数据(巢穴温度、巢穴深度及巢口倾角)进行单个样本的Shapiro-Wilk正态性检验,若某单因子的2 组数据均符合正态分布则使用独立样本t检验,若有一组数据呈离散型则使用Mann-WhitneyU检验。对2 个非连续性型数据(巢口方向和湿度),运用卡方检验分析蜥蜴对巢口方向和巢穴内湿度的选择差异性。同时,为比较同种蜥蜴的巢穴特征在采样巢穴之间是否具有差异性,对 2种蜥蜴的5 个指标均分别进行卡方检验。此外,对5 个指标进行主成分分析[43]。各数值型变量在描述时采用mean±SE 表示;所有检验的显著水平设定为0.05,即当p<0.05 时为差异显著。所有统计分析利用软件SPSS 21.0进行。
3 结果
3.1 2种沙蜥巢及巢穴特征
采用观察法共采集南疆沙蜥巢穴22 处,叶城沙蜥巢穴19 处。南疆沙蜥巢穴深度为18~61 cm,平均为(33.77±2.81)cm;巢穴温度最低为25.0 ℃,最高为40.4 ℃,平均值为(30.24±0.79)℃;巢口均为 向上倾斜;倾角最低21.8°,最高42.0°,平均值为(27.67±1.02)°。叶城沙蜥巢穴深度为18~40 cm,平均值为(26.42±1.46)cm;巢穴温度最低为26.2 ℃,最高为43.2 ℃,平均值为(33.55±1.20)℃;巢口均为向上倾斜;倾角最低为11.3°,最高为31.0°,平均值为(23.06±1.13)°(表1)。独立样本t检验结果显示,2种沙蜥巢穴温度差异显著(p=0.028)。巢穴深度和巢口倾角的组间Mann-WhitneyU检验表明,2种沙蜥巢口倾角组间差异显著(Z=−2.458,p=0.014),而巢穴深度差异不显著(Z=−1.523,p=0.126)。同种蜥蜴的巢穴特征比较显示,南疆沙蜥与叶城沙蜥对巢穴深度、温度及倾角3 个特征均未呈现出显著的组内差异(表2)。
采集的22处南疆沙蜥巢穴中,有20处巢穴湿度为极干旱(DRY+),1 处为湿润(WET),1 处为干旱(DRY);19处叶城沙蜥巢穴中有18处为极干旱,1处为干旱。调查表明南疆沙蜥有7 处洞穴巢口方向为东,6 处为西南,3 处为西北,洞口朝向东北和东南的分别有2 处,朝向南和北的分别有1 处;叶城沙蜥的洞穴洞口有6处朝向西南,4处朝向东北,朝向西、西北、北以及东南的分别为2 处,另有1 处洞口朝向为东(表1)。卡方检验分析结果表明,南疆沙蜥和叶城沙蜥组内的巢穴湿度结果均呈现显著差异(χ2=32.818,p<0.001;χ2=15.211,p<0.001,表2);但组间差异不显著(χ2=1.076,p=1.000)。南疆沙蜥的巢口方向在东和西南方向的频次较高,但组内未达到显著水平(χ2=11.091,p=0.091);叶城沙蜥的巢口方向偏好西南,但组内未达到显著水平(χ2=6.421,p=0.378,表2),且组间无显著差异(χ2=8.274,p=0.289)。

表1 南疆沙蜥与叶城沙蜥5个巢穴指标数据Tab.1 Details of the five nest indices for Phrynocephalus axillaris and Phrynocephalus forsythii
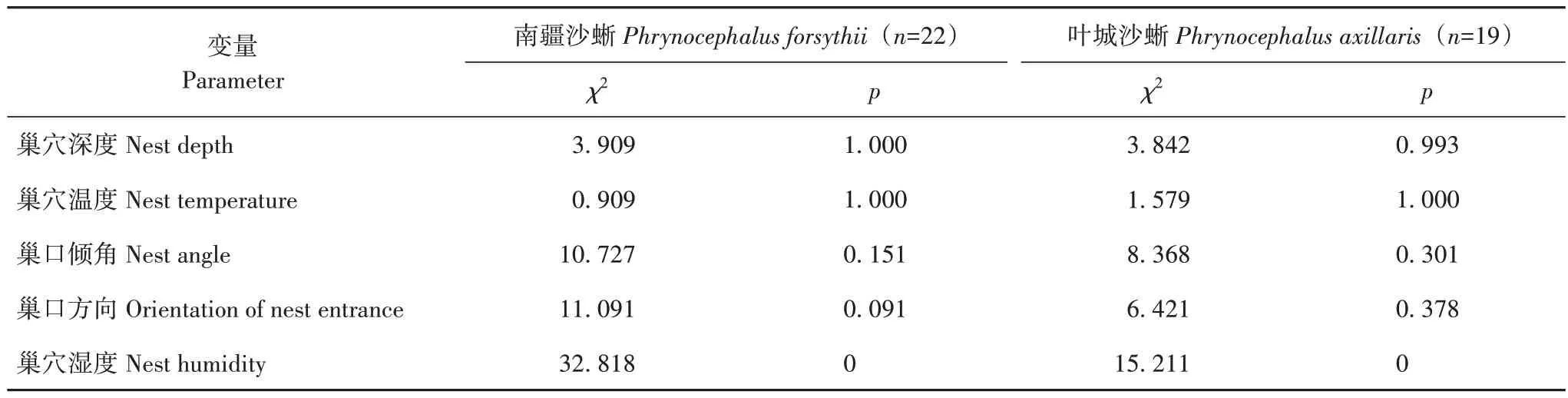
表2 同种蜥蜴采样巢穴间巢穴特征的比较Tab.2 Comparison in nest parameters of the sampled nests within species group
3.2 2种沙蜥巢址选择主成分分析
通过对22 个南疆沙蜥和19 个叶城沙蜥巢穴指标(巢穴温度、湿度、深度、巢口倾角及巢口方向)进行主成分分析,得到5 个因子的特征值和相关矩阵(表3、表4)。结果表明,南疆沙蜥巢址主成分 3(0.988)和叶城沙蜥巢址主成分3(0.942)接近1.000,所以将2种沙蜥的5 个生境因子聚成3 类,且前3 个主成分的累计贡献率(71.988%,76.108%)大于60%,表示这3 个主成分可反映原巢址选择参数具有的信息(表3)。
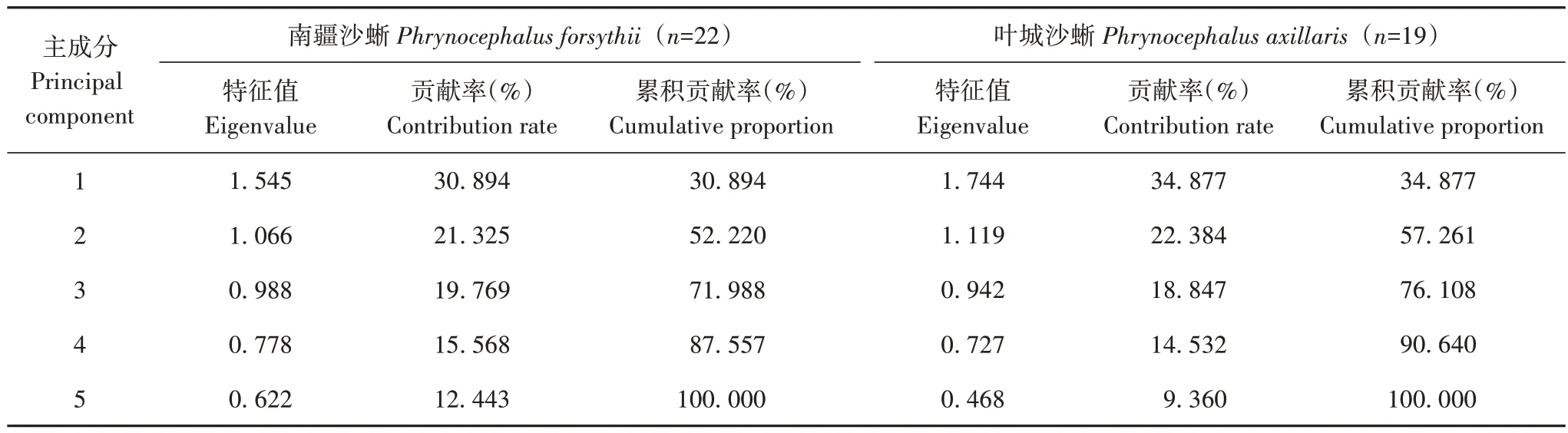
表3 2种沙蜥各巢址选择的主成分特征值Tab.3 The principal component analysis of nest-site selection of two species of lizards

表4 2种沙蜥巢址选择的各旋转后因子成分矩阵Tab.4 Matrix of rotated factors from principal component analysis of nest-site selection in the two lizard species
从数据分析结果(表4)可知,南疆沙蜥和叶城 沙蜥巢址选择生境因子的第一主成分中,巢穴深 度(0.782;-0.619)、温度(-0.635;0.832)及湿度(0.670;0.674)的特征值绝对值均偏高。2种沙蜥的第二主成分都是巢口倾角,其特征值绝对值分别高达0.929和0.898。巢口方向则在2种沙蜥的第三主成分中,其特征值分别为0.965 和0.970。4 因子主成分分析结果与5 因子主成分分析结果类似,当被分析因子中含有巢口方向时,该因子往往在最后一个主成分中。
4 讨论
本研究比较了南疆沙蜥和叶城沙蜥巢址选择的5种因子,结果表明2种沙蜥选择的巢址特征不尽相同,二者对巢穴生境均有明显的选择性。南疆沙蜥选择的巢穴温度显著低于叶城沙蜥的巢穴温度(p=0.028);巢倾角则是南疆沙蜥显著高于叶城沙蜥(p=0.014)。对于巢穴深度,数据显示南疆沙蜥高于叶城沙蜥,却未达到显著水平(p=0.126)。二者的巢口方向均无偏好,具有随机性。对于巢穴湿度,2种沙蜥都会选择合适湿度(干燥)的地点筑巢。
蜥蜴的巢穴深度与卵孵化及后代存活率相关,亲代可通过改变巢穴深度或调节胎生形式的母体体温等,对孵化期间的温度进行直接和间接的精确控制,使巢穴内处于适合卵孵化或后代存活的温度[44−45]。本研究结果显示,南疆沙蜥的巢穴深度为(33.77±2.81)cm,叶城沙蜥的巢穴深度为(26.42±1.46)cm,二者并未达到显著差异水平,说明巢穴深度可能不是2种蜥蜴巢穴选择的主要特征。另一方面,胚胎存活率是影响卵生动物巢址选择的重要因素之一[46−48]。卵胎生的南疆沙蜥胚胎能够得到母体的适当保护,后代成活率由于具有较好的亲代抚育而得到保障,因而巢穴更能反应胚胎孵化温度。卵生沙蜥相较于巢穴深度等物理环境,巢穴温度对卵的孵化率、胚胎存活率及后代表型和适合度的影响更大[49],且雌性蜥蜴在孵化期可能具有不同的巢址选择需求[50]。因此,虽然南疆沙蜥与叶城沙蜥对巢址深度选择不具有差异性,但不同的生殖方式可能导致二者在巢穴温度等方面的选择具有差异,本研究结果恰符合这一猜想。
爬行动物在适宜的体温下呈现正常的行为并高水平地表达生理潜力,极端体温则影响其运动、摄食、同化、能量代谢、生长繁殖、胚胎发育和免疫功能,甚至可导致死亡,而环境温度是改变爬行动物体温的重要原因之一[51]。因此巢穴温度不仅能够影响亲代蜥蜴的行为,还影响后代孵化率甚至有可能对维持整个种群存活都至关重要。南疆沙蜥的巢穴温度为(30.24±0.79)℃,显著低于叶城沙蜥的巢穴温度(33.55±1.20)℃,主成分分析结果也显示巢穴温度均在2种沙蜥的第1主成分中,这些结果表明叶城沙蜥比南疆沙蜥更喜高温。研究指出叶城沙蜥比南疆沙蜥的分布海拔更低、更耐干旱[52],本研究结论与这一发现相符。除生殖方式差异和生态环境的因素外,2种蜥蜴选择的巢内温度还可能与它们的体形大小有关。在相似种类的动物中,体形大的动物散热面积小,选择的环境温度就较低[53],但研究区域内的南疆沙蜥与叶城沙蜥体形差异仍有待进一步调查。
就巢口倾角而言,南疆沙蜥为(27.67±1.02)°,显著高于叶城沙蜥的(23.06±1.13)°,这可能与研究地的土壤质地或有利于2种沙蜥选择合适的巢穴深度、温度有关,其中具体关系还有待继续研究。
卡方检验结果发现,南疆沙蜥和叶城沙蜥巢穴的开口方向具有随机性,种间无显著差异。主成分分析显示2种沙蜥的巢口方向均处于第3主成分中,表明巢口方向不是影响蜥蜴筑巢的主要因素。
对于巢穴湿度,南疆沙蜥和叶城沙蜥种群内的选择结果均表现出显著性差异,但二者间未达到显著差异水平,2种沙蜥均喜好在干燥的土壤湿度下筑巢。该结果说明南疆沙蜥和叶城沙蜥在巢址选择时对巢穴内土壤湿度可能存在一定偏好。由于本研究缺乏随机巢址的湿度数据,采样点的湿度与一般土壤湿度的差异性尚不明确,因此2种沙蜥对土壤湿度的偏好及选择性有待进一步确认。
在许多卵生的爬行动物中,雌性对筑巢地点的选择会影响产卵的完整性、卵的存活率及后代表型[54−56]。根据母性偏好子代表现假说,雌性动物会选择适合子代存活的地方筑巢[44]。卵生动物亲代通过选择特定的筑巢地点是使后代孵化率最大化的重要因素之一[8]。南疆沙蜥和叶城沙蜥在塔里木盆地分布有重叠,叶城沙蜥分布区更宽,包括敦煌盆地、吐鲁番—哈密盆地,同时二者具有不同的生殖方式和生境需求。本研究表明南疆沙蜥和叶城沙蜥巢址选择均具非随机性,巢穴温度和巢口倾角具有显著种间差异。这种特定的巢址特征可能对二者卵的孵化及后代存活率具有重要意义。本研究首次就新疆尉犁县南疆沙蜥与叶城沙蜥的巢址特征进行了比较,丰富了该地区2 个物种的生态学资料,同时为后续更为详尽的巢址研究奠定基础。
猜你喜欢
歌海(2021年6期)2021-02-01 11:27:18
艺术家(2020年6期)2020-08-10 08:35:28
百家讲坛(2019年20期)2019-04-30 06:48:24
阅读与作文(小学高年级版)(2018年9期)2018-10-25 11:07:36
传奇故事(破茧成蝶)(2017年6期)2017-11-25 03:52:16
传奇故事(上旬)(2017年6期)2017-06-17 21:50:57
阅读(科学探秘)(2017年5期)2017-05-30 10:48:04
中国卫生(2016年7期)2016-11-13 01:06:40
科技知识动漫(2016年3期)2016-03-22 20:44:02
家教世界·创新阅读(2014年11期)2014-12-15 04:15:04
