
大白菜Ogura 胞质雄性不育恢复材料的创制与检测
2023-05-13 09:25:20魏小春原玉香赵艳艳杨双娟苏贺楠王志勇王晓青牛刘静张晓伟
中国瓜菜 2023年4期
魏小春,原玉香,赵艳艳,杨双娟,苏贺楠,王志勇,王晓青,冉 冉,李 林,牛刘静,张晓伟
(河南省农业科学院园艺研究所 郑州 450002)
国家历来重视农业生产,自2004 年以来,中央“一号文件”把种业发展提升到国家战略高度。近年来,我国种业发展势头良好,取得了一些突破,水稻、玉米、小麦等三大主粮高效育种技术体系逐渐完善,主要农作物自主选育品种占比在95%以上,基本实现主要粮食作物良种全覆盖。但很多种子的进口量远大于出口量[1],抗根肿病耐抽薹十字花科蔬菜品种就是其中一个例子,而且大都是雄性不育系制种,利用雄性不育系材料进行种质创新,最有效的办法就是筛选对应合适的恢复系。然而,在大白菜类蔬菜中尚未发现Ogura 雄性不育恢复基因材料。
Ogura 雄性不育系是日本科学家Ogura 首次在萝卜中发现的天然雄性不育类型,雄性败育彻底,为迄今发现的芸薹属作物不育源中不育性最理想的类型[2],越来越多的Ogura 雄性不育系被用于生产白菜类蔬菜杂交种,杂交种纯度可达到100%。但由于Ogura 是一种典型的母系遗传性状,降低了种质资源的多样性,如日本大白菜AKIMEKI 是一个抗根肿病的品种,含有3 个抗性基因,但由于其为Ogura 胞质雄性不育系杂交种,不能直接用于种质资源创新。降低Ogura 胞质带来的潜在风险,加强种质资源的创新利用,可以通过实现胞质多样化来开发新的胞质资源,如CMS-Pol、CMS-Nap、CMS-Hau、CMS-orf 220[3],也可以创制Ogura 胞质不育恢复系,恢复不育材料的育性。国内外现已有甘蓝类蔬菜Ogura 恢复系创制的报道[4-5]。主要是通过芥蓝(CC 基因组)与携带恢复基因(Rfo)的油菜(AACC 基因组)远缘杂交后,利用胚挽救技术获得种间杂种F1,利用恢复基因标记检测,以不育系为母本、恢复基因阳性单株为父本,进行连续回交后得到BC3代,获得染色体数为18、遗传背景接近于芥蓝Ogura CMS 的恢复材料[5]。
远缘杂交在创制新的作物类型、利用外来属种的特殊有利性状丰富作物变异类型、创制新的雄性不育源、探索和研究生物进化等方面具有重要意义。在十字花科作物中,为了克服远缘杂交中幼胚死亡或败育,研究人员利用胚胎挽救技术获得了结球甘蓝×大白菜、萝卜×大白菜、大白菜×萝卜等远缘杂交种[6-7]。为了促进两属作物间的优良基因交换,Bannerot 等[8]利用Ogura 萝卜不育胞质材料转育成甘蓝、甘蓝型油菜不育系。方智远等[9]利用胚挽救技术将萝卜雄性不育系转移至结球甘蓝。程雨贵等[10]对萝卜和甘蓝正反杂交后的杂种胚进行胚挽救,结果表明,以萝卜作为母本较容易成功,而以甘蓝为母本则很难成功。利用远缘杂交技术进行恢复基因转移,首先要知道恢复基因所在的染色体位置。萝卜中Ogura 胞质不育恢复基因所在的位置为R9 染色体[11]。将恢复基因导入到甘蓝型油菜后,通过原位杂交等技术将Rfo基因定位到甘蓝型油菜C9 染色体上[12]。恢复基因的成功定位及克隆[11,13]也为定向转移恢复基因及分子标记开发提供了背景及序列基础。
迄今尚未发现白菜类作物中存在Ogura 雄性不育系的恢复系,因此迫切需要创制白菜类作物Ogura 细胞质雄性不育的恢复材料。由于油菜与萝卜中恢复基因分别在C9 及R9 染色体上,相对于甘蓝类蔬菜(CC 基因组)恢复系材料的创制,白菜类蔬菜(AA 基因组)会更加困难。通过远缘杂交将甘蓝型油菜育性恢复基因Rfo导入到大白菜中,然后将种间杂种进行连续回交,获得回交BC1、BC2和BC3代育性恢复材料。然而,这些回交材料的遗传背景与大白菜仍存在一定差异,还需进一步回交。在保留恢复基因Rfo的同时,需尽可能消除甘蓝型油菜的遗传背景,这对最终实现Ogura CMS 白菜类蔬菜的育性恢复具有重要意义。
1 材料和方法
1.1 雄性不育系恢复系材料选择
Ogura 雄性不育系大白菜Tyms(Brassica rapassp.pekinensis,AA 基因组)由河南省农业科学院园艺研究所叶类蔬菜课题组提供;含恢复基因的甘蓝型油菜P1755(Brassica napusL.,AACC 基因组),由河南省农业科学院经济作物研究所张书芬研究员馈赠。
Ogura CMS 大白菜杂交商品种AKIMEKI(Brassica rapassp.pekinensis,AA 基因组),购自日本农林社株式会社(Norin)。
以大白菜Tyms 为母本、甘蓝型油菜P1755 为父本,制备种间杂交组合。蕾期进行套袋、人工授粉。杂交前2~3 d,摘除父本花序上已经开放的花朵,套袋隔离。套袋父本开花当天取花药,与母本植株适宜大小的花蕾进行剥蕾授粉。授粉后,套袋进行隔离,以防外来花粉污染。
杂交F1:Tyms×P1755。
第1 代回交:母本为Ogura 雄性不育系大白菜Tyms,父本为种间杂交F1代育性恢复Rfo单株(AAC+基因组)。
第2 代回交:母本为Ogura 雄性不育系大白菜Tyms,父本为BC1代回交后通过标记筛选获得的具有良好育性的恢复单株(AA+基因组)。
第3 代回交:母本为Ogura 雄性不育系大白菜Tyms,父本为BC2代回交后通过标记筛选获得的具有良好育性恢复、遗传背景更接近大白菜母本的优良单株(AA 基因组)。
上述材料均由河南省农业科学院园艺研究所叶类蔬菜课题组提供,均定植于河南现代农业研究开发基地,常规栽培管理,2018 年春季开始,2021年获得BC3、BC4株系。
1.2 染色体观察
试验前期在试验田分别选取Ogura CMS 不育系(Tyms)大白菜母本、恢复系(P1755)油菜父本及F1、BC1、BC2、BC3代中育性不同的植株,分别标号,并取其花蕾,分别用卡诺氏液固定保存,定期更换卡诺氏液。
之后在纯水中选择固定好的适当大小的花蕾,2 倍显微镜下解剖,取出一粒完整花药放入1 mol·L-1盐酸中,并在60 ℃烤片机上预热2 min。酸解完成后,取出花药,机械敲碎,用改良的苯酚品红染液染色,并在40 倍镜下筛选处于减数分裂时期的花药,分别标记保存。将已筛选出的花蕾进行酶解、低渗与荧光染色制片等一系列操作,并对育性不同的各世代植株进行染色体细胞学分析,荧光显微镜(型号:LEICA CTR4000)观察(花粉粒染色体各个时期配对情况),观察并记录染色体数,比较染色体形态与分裂是否异常,计算恢复基因的传递效率[14]。
1.3 植物细胞学倍性鉴定及细胞分裂过程观察
流式细胞仪用于鉴定远缘杂交杂种后代的倍性,取杂种后代植株叶片,利用流式细胞仪(Cy-Flow Cube 8)(德国PARTEC 公司)检测植株倍性。在本试验中,以大白菜(Tyms)、甘蓝(CC)、甘蓝型油菜(P1755)为模板,将电压调节至523 V,根据图中峰值位置判断植株倍性[15]。利用大白菜Ogura 雄性不育系Tyms 为母本、含有恢复基因的油菜P1755 为父本进行远缘杂交,获得种间杂交种,观察各个时期染色体减数分裂的变化,即细线期、粗线期、终变期、减I中期、减I后期、减Ⅱ中期、减Ⅱ后期[16]。
1.4 引物开发及恢复基因的PCR检测
以萝卜Ogura 胞质不育恢复基因Rfo序列信息(NCBI 登录号:AJ550021.1)为目标序列,在大白菜与油菜基因组数据库(http://www.brassicadb.cn/#/Download/)进行Blast 比对。萝卜Ogura 细胞质雄性不育恢复基因Rfo位于R09 染色体上,甘蓝型油菜A09 染色体、C09 染色体以及大白菜A09 染色体上存在Rfo同源序列。根据Rfo基因与亲本的序列比对结果,设计开发出2 对引物,即RsRfo-F/RsRfo-R 及Rfo-105F/Rfo-545R。
恢复基因PCR 所用引物为Rfo特异引物(表1)。采用的PCR 扩增体系为20 μL,体系中的具体成分含量为:PCR 酶TaqMix 10 μL,F-primer 1 μL,R-primer 1 μL,DNA 2 μL,ddH2O 6 μL。PCR 扩增反应的参数设置为:94 ℃预变性5 min,94 ℃变性50 s,55 ℃退火50 s,72 ℃延伸50 s,共35 个循环,最后72 ℃延伸5 min,16 ℃保存PCR 产物5 min。以此进一步确定Rfo特异分子标记筛选结果与花粉育性表现一致,从而选择出一株遗传背景与回交母本大白菜最相似,且含有Rfo恢复基因的育性恢复单株,用于下一步的回交转育。
1.5 回交后代恢复基因传递效率及检测
回交后代恢复基因Rfo的传递效率(transmission rate,TR)的计算,TR/%=Rfo阳性单株数目/群体总单株数×100。
恢复系新种质的检测:利用所创制的BC3阳性单株对Ogura CMS 大白菜商品杂交种AKIMEKI进行恢复试验,材料种植于河南现代农业研究开发基地,恢复基因Rfo及不育基因Orf138检测引物见表1。

表1 本项目中所涉及到引物序列
2 结果与分析
2.1 对杂交后代F1(AAC)进行形态学及流式细胞仪检测
Ogura 雄性不育系大白菜Tyms 抱茎生长,叶片较薄、叶色较绿。甘蓝型油菜P1755 叶色较深,主花序明显,抱茎生长叶片较厚且有蜡质层。杂种一代的生长势较旺,花枝发达,叶色浓绿,蜡粉较多,杂种一代在形态上有很明显的区别,更接近于普通的甘蓝型油菜(图1)。

图1 亲本以及子代花、叶、植株的表型形态
利用流式细胞仪检测技术,对高速流动的单细胞悬浮液进行了多项参数测量并对DNA 含量进行分析,经信号处理后绘制出DNA 含量分布曲线,能够直观地观察到植株的倍性。观察发现,大白菜Tyms 在减数分裂间期的DNA 荧光强度峰值位于横坐标35 位置,P1755 位于横坐标75位置,F1代植株减数分裂间期DNA 的荧光强度峰值的横坐标位置与双亲峰值对应的横坐标位置的平均值55 接近,此检测结果表明,AAC 杂交成功(图2)。
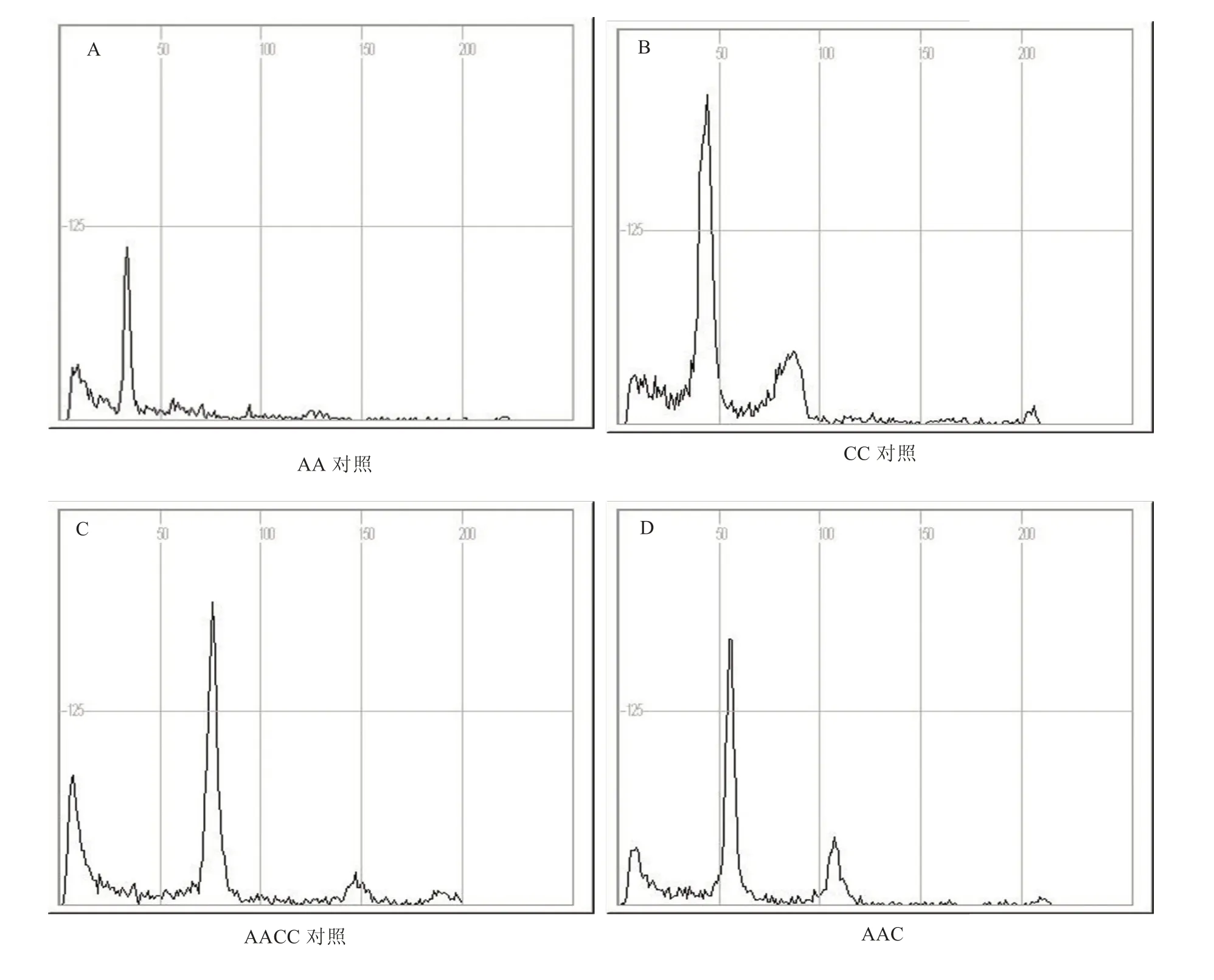
图2 大白菜Tyms 与油菜P1755 杂交后代的流式细胞仪检测
2.2 亲本及杂交种F1细胞分裂过程观察
由图3-a4 可知,Tyms 染色体数目为20 条,由图3-b4 可以看出,P1755 染色体数目为38 条,由图3-c4 可以看出,减数分裂一代F1染色体数目为29 条;由图3-a1~a7 可知Tyms 染色体为二倍体并且减数分裂正常;由图3-b1~b7 可知,P1755 染色体为四倍体并且减数分裂正常;由图3-c1~c7 可知,F1为三倍体并且减数分裂正常。F1为真杂种,所以经过染色体鉴定可以进一步确定杂交成功。
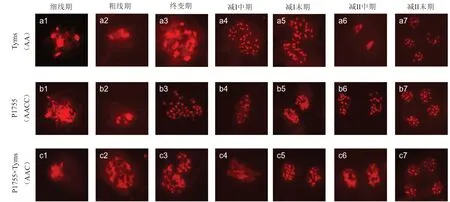
图3 大白菜与油菜杂交后代减数分裂染色观察
选取BC1、BC2、BC3回交后代处于终变期、减数第一次分裂后期和减数第二次分裂时期的花粉母细胞进行染色体细胞学观察(图4)。结果表明,大多数花粉母细胞在减数分裂后期不均等分离,也有落后染色体(图4-a)、染色体排列混乱(图4-b)、单价体(图4-c)、三价体(图4-d)等异常行为频繁出现,从而导致不育株常出现非整倍染色体的现象。说明回交后代育性不同很大程度上由减数分裂时期异常的染色体行为所致,A 基因组和C 基因组间的种间杂种在减数分裂过程中,常存在A-C 基因组间的非同源染色体配对和异源联会的现象。
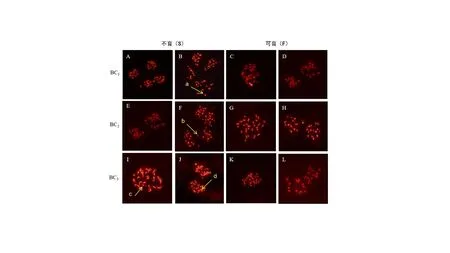
图4 回交世代单株减数分裂时期染色体行为观察
2.3 回交后代染色体数统计
笔者试验的母本为二倍体Ogura-CMS 不育系Tyms 大白菜(AA 基因组),父本为含恢复基因Rfo的四倍体甘蓝型油菜恢复系P1755(AACC基因组),它们所对应的染色体数分别为20 条和38 条。在前期获得种间杂交F1代育性恢复Rfo单株(AAC+基因组)的前提下,结合Rfo特异分子标记辅助筛选,继续进行回交转育,结合田间花粉发育情况,通过对BC1、BC2、BC3回交后代的染色体观察,发现随着回交代数的增加,回交后代染色体数接近母本(表2)。同时,笔者已经在BC3代中发现有染色体为20 条的、育性恢复的、表型接近母本大白菜的Rfo阳性恢复材料F10(表2),下一步应将其作为重点单株进行BC4代的回交转育。

表2 回交世代后代可育及不育单株的染色体数目统计
2.4 回交后代中恢复基因的分子标记快速检测
以萝卜Ogura 细胞质雄性不育恢复基因的Rfo序列为目标序列,通过与大白菜和油菜基因组数据库的比对,发现在甘蓝型油菜A09 染色体、C09 染色体以及大白菜A09 染色体上存在Rfo同源序列,萝卜Rfo基因与其同源率分别达到89.61%、89.66%及89.61%。根据Rfo基因与亲本的序列比对结果(图5),设计开发Rfo基因检测引物并用于检测(图6)。

图5 不同来源Rfo 基因多序列比对

图6 恢复基因Rfo 的PCR 分子标记检测
利用Rfo特异标记对获得的37 株BC3回交后代进行筛选,结果表明,37 株大白菜-甘蓝型油菜BC3后代中有17 株能扩增出清晰明亮的特异性目的条带,这17 株BC3回交后代中含有Rfo基因。该Rfo特异性分子标记筛选结果与花粉育性相一致,可以作为Ogura CMS 大白菜恢复植株Rfo基因的分子特异性标记。结合形态学、细胞学染色体观察及恢复基因Rfo全长特异分子标记技术,在BC3回交群体中发现1 株染色体数为20、遗传背景类似大白菜的单株。
2.5 回交后代中恢复基因的传递效率及检测
结合Rfo特异标记筛选结果对所有回交Rfo阳性单株育性进行调查,发现扩增出特异目的条带的17 株Rfo阳性单株均出现花粉,与Rfo标记筛选结果相一致。但是,对所有Rfo阳性单株花粉饱满程度进行调查的结果显示,有些单株花粉饱满,有些单株花药细小,仅有少量花粉。未扩增出明显条带的20 株,开花后仍表现为不育,可能由异源染色体障碍造成非整倍染色体现象所致。这17 株BC3代Rfo阳性株的Rfo基因的传递效率为46%,相对于BC2代Rfo阳性株40%的Rfo基因传递效率有所提高。该结果进一步表明,Rfo基因在BC1、BC2、BC3回交后代中可以稳定传递,传递效率正常且逐代增加,能够保证回交后代的育性稳定,花粉质量也有所提高。
利用所创制的大白菜Ogura CMS 不育系恢复系对Ogura CMS 大白菜商品杂交种AKIMEKI 进行育性恢复,结果表明,在43 个杂交后代中,发现11 个植株有花粉,通过恢复基因分子标记Rfo 检测发现该11 个单株含有Rfo基因;同时,由于Ogura CMS 为胞质线粒体缺陷与核基因互作的雄性不育,因此,在43 个杂交后代中,均会存在线粒体缺陷orf138,发现43 个单株中均存在胞质线粒体,与分子标记检测结果一致(图7),证明所创制的恢复系恢复基因有效。

图7 对AKIMEKI 进行育性恢复的43 个杂交后代花药器官观察、恢复基因及胞质基因Orf138 检测
3 讨论与结论
3.1 远缘杂交中种间三倍体的成功创制及利用
笔者将大白菜Tyms 作为母本和甘蓝型油菜品种P1755 作为父本进行远缘杂交,使大白菜具有Rfo恢复基因,用分子标记技术对其进行前景及背景的筛选,从而得到具有再生能力的杂种后代。研究中还对种间杂种的形态学、倍性、染色体做了观察分析,最终确定了种质资源AAC 的成功创制,为进一步优化种质资源提供了基础。
Ogura 胞质雄性不育为十字花科作物杂种优势利用的一个重要途径,而且它具有不育性稳定的特点[17]。但是,Ogura 胞质雄性不育是一种典型的母性遗传特征,其F1都是雄性不可育,无法自交分离,这对种质资源的开发和利用具有一定的局限性。研究和创制Ogura CMS 恢复系是一种行之有效的途径。因此,开发利用Ogura CMS 恢复材料,不仅有助于种质资源的多元性而且有利于种质资源的重复利用,对于提高我国十字花科蔬菜育种国际地位有着至关重要的作用。迄今为止,在白菜类蔬菜中尚未发现Ogura 胞质雄性不育系的恢复系,急需引入Ogura CMS 的恢复系,创制出白菜类蔬菜的Ogura CMS 不育系恢复材料。
甘蓝型油菜是由甘蓝和白菜通过自然种间杂交后双二倍化后形成的,远缘杂交已成为扩大其遗传背景、改良种质资源的重要手段。然而,由于杂交后代存在生殖障碍等问题,需要进行多代回交,甚至于多轮胚挽救工作[18]。徐书法等[19]在研究大白菜、芥菜和甘蓝型油菜的远缘杂交时发现,甘蓝型油菜与白菜间的杂交亲和性较强,且存在单侧杂交不亲和性。笔者的研究将Ogura CMS 大白菜Tyms与含有恢复基因的甘蓝型油菜P1755 杂交,一定程度上克服了远缘杂交障碍,相比甘蓝类蔬菜,白菜类蔬菜与甘蓝型油菜亲和性较强,较易获得种间杂交种。本试验结果表明,F1后代AAC 育性较高,且在减数分裂时期,A 染色体间配对稳定,整倍体配子出现频率较高。在F1种间杂交种,可将萝卜源的Rfo基因稳定固定并遗传,通过回交转育,可成功将Rfo基因导入Ogura CMS 大白菜的基因组中,是Ogura CMS 大白菜遗传背景拓展的重要手段之一。
3.2 远缘杂交后代染色体行为配对影响花粉育性
由于远缘杂种的亲本来源于不同的种或属,其后代相应就含有2 套或2 套以上的染色体组,育性通常较低或高度不育,染色体行为异常常发生在配子形成、胚胎发育和植物生长过程中。崔辉梅等[20]、乔海云等[21]在芸薹属远缘杂交种间杂种的花粉母细胞减数分裂时期染色体观察中均发现,杂交后代染色体间配对、重组、分离通常较为混乱,常产生不减数配子,使得配子遗传多样化,分离世代长,理想基因型不易出现。笔者通过对回交后代减数分裂染色体细胞学观察发现,在减数分裂后期多为不均等分离,也有染色体排列混乱、三价体、单价体、落后染色体等异常行为频繁出现,从而导致不育株常出现非整倍染色体的现象,说明回交后代育性不同很大程度上是由减数分裂时期异常的染色体行为所致。
3.3 恢复基因分子标记提高回交育种效率
相对于传统农艺性状选择方法,分子标记辅助选择具有显著缩短育种进程、加快新品种选育的优越性[22]。笔者课题研究人员在前期研究中,通过甘蓝和甘蓝型油菜远缘杂交和胚挽救获得F1代杂交种,通过回交获得BC1代植株。然而,BC1代的花粉活力、育性稳定性较差,人工授粉难以结实,无法直接利用[18]。有研究发现,在大白菜和羽衣甘蓝杂交后的回交过程中,BC4世代亲和指数显著增大[23]。
笔者利用Ogura 胞质雄性不育系大白菜为母本,用获得的BC1代中携带有Rfo阳性的植株进行回交。通过Rfo特异分子标记进行辅助选择,结合育性检测、形态标记、细胞学观察对遗传背景进行分析,分别筛选获得了BC2、BC3代Rfo单株,用于下一代回交转育。虽然笔者已经在BC3代中初步获得了育性稳定、染色体数为20、含有Rfo恢复基因、形态学性状总体偏向大白菜的阳性单株,进一步对Ogura CMS 商品种进行恢复测试,结果表明,所创制恢复系恢复基因有效;但仍需进行大量回交和筛选,从而进一步消除恢复材料甘蓝类油菜基因的背景。随着进一步BC4代的回交转育,Rfo基因有望稳定固定在Ogura CMS 大白菜基因组,为大白菜Ogura 雄性不育恢复系材料的创制提供了材料基础。
猜你喜欢
科学(2023年2期)2023-05-30 08:57:46
环境科学与管理(2023年3期)2023-04-29 03:02:27
麦类作物学报(2022年5期)2022-05-20 09:43:24
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
西藏农业科技(2018年4期)2018-04-25 06:39:28
现代园艺(2017年21期)2018-01-03 06:41:41
上海农业学报(2017年3期)2017-04-10 12:39:24
中国种业(2016年11期)2016-12-01 05:43:51
西南农业学报(2016年6期)2016-04-16 05:12:46
