
HCO3-Ca 型岩溶湖附着藻类生长对沉水植物退化的驱动研究
2023-05-09 00:55柴晓贞刘书敏
湖北农业科学 2023年4期
柴晓贞,刘书敏
(1.滨州市海洋发展研究院,山东 滨州 256699;2.同济大学,上海 200092)
随着流域自然环境因素的变化及人类经济社会活动干扰强度的不断加大,部分湖泊水生态系统结构及功能正呈现退化趋势[1,2]。退化湖泊在一定程度上丧失了自身调节功能,生态系统缓冲能力逐渐下降[3,4],具体表现为水质不断恶化,水生植物退化现象日趋严重,沉水植物分布范围缩小、分布深度下降,生物量及物种多样性下降[5],浮游藻类大量增殖逐渐成为生态灾害等[6,7]。恢复湖泊生态功能的完整性及系统健康并保持其自身良性循环成为湖泊保护工作的关键[8,9]。目前,较为常用的技术手段为通过组建退化生态系统的原有生态功能及结构[10],恢复系统功能及生物学潜能,确保恢复后生态系统具有长期稳定性[11]。沉水植物的恢复工作已经成为水生态系统恢复的重点工作内容之一。
岩溶水作为人类生产生活的重要水源,受气候变化和人类活动的影响较大,其在恒温且无光的管道或裂隙内传输[12],且岩溶地下水排泄形成的湖区内部生物群落结构不断变化[13,14]。岩溶水环境质量主要受含水介质及补给水源的影响,其通过工业污染、农业污染及生活污染影响水体[15],岩溶生态系统易受到外界环境因子的破坏[16,17],这实质上反映了岩溶生态系统环境承载的阈值弹性小、生态环境脆弱的特点[18]。因此,对脆弱岩溶生态系统中敏感环境下的退化触发机制进行研究具有重要意义。
沉水植物是水体中重要的初级生产者,是水生生态系统中的重要组成者之一[19]。当前对沉水植物退化驱动因子的研究较多集中于水体透明度、溶解氧、营养盐及底质方面[20,21],而对附着藻类驱动的研究较为鲜见。根据稳态转换理论,一旦大量外源污染引入,在微小的外来因子扰动下,生态系统则会由原本清水稳态转变为浊水稳态,修复困难程度加大[22,23]。藻类与沉水植物生态位高度重叠,竞争激烈[24],营养元素及光照等是两者竞争的重要环境因素[25],研究表明沉水植物叶片上附着大量增殖的藻类,不仅导致了沉水植物叶片细胞外扩散层厚度增加并降低无机碳的供应能力,且附着藻类对光照及无机碳的竞争影响了沉水植物的正常生长[26,27]。针对湖泊沉水植物退化的现状,通过对HCO3-Ca 型岩溶湖沉水植物退化驱动因子的研究,有利于为岩溶水退化生态系统修复提供治理思路。
本研究以典型HCO3-Ca 型退化岩溶湖灵水湖为研究对象,通过设置外加钙离子(Ca2+)和碳酸氢根(HCO3-)不同浓度梯度,探究不同氮(N)、磷(P)营养盐浓度水平下附着藻类叶绿素a(Chl-a)的浓度变化情况。结合灵水湖水域环境现状,对灵水湖沉水植物的退化驱动因子进行探究,以期为保障灵水湖作为水源地的水质安全和岩溶湖沉水植被的生态恢复提供治理思路。
1 材料与方法
1.1 研究地点及样本采集
灵水湖为典型的熔岩地下河出水口的大型泉群,清泉从四周石缝中涌出,位于广西南宁市武鸣区(图1),目前为武鸣区城区约12 万人口的地下水饮用水源地。灵水湖水面总面积为2.93 万m2,水深2.0~3.5 m,流量2~3 m3/s,全年的水温变化幅度为1 ℃。灵水湖发育类型为集中径流带式,在水文地质普查工作中被定为地下河出口,有9 个出水口,分别由不同方向发育的网络状裂隙或者小管道径流汇集而成。灵水湖水源主要为降雨补给及地表水的渗漏。近年来岩溶泉、地下河出口处沉水植物逐渐退化的趋势愈加明显。灵水湖沉水植物主要以苦草(Vallisneria natans)为优势种,满铺于水底,呈现“水下森林”景观,且水质一直维持在地下水Ⅰ类标准,于2010 年植物生物量直线下降,沉水植物呈逐渐消亡趋势,至2013 年湖区内沉水植物逐渐消失。
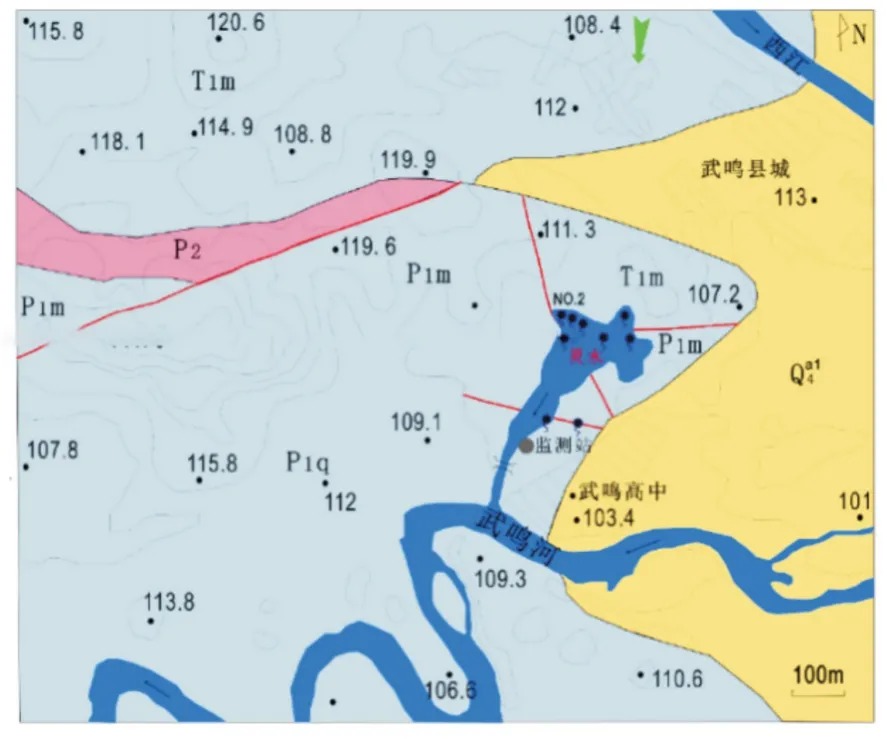
图1 灵水湖区位
1.2 设计
通过设计Ca2+和HCO3-不同浓度梯度探究不同N、P 营养水平下水生态系统初级生产者生物量的响应情况。采用容积为20 L(45 cm×30 cm×15 cm)的塑料箱进行生态箱设计,Ca2+、HCO3-浓度水平模拟灵水湖现状进行设计,通过向生态箱中投加长度为20 cm 的人工水草模拟小型生态系统沉水植物,从而得以检测附着藻类叶绿素a的含量。
如表1 所示,设置6 种生态箱,每种生态箱的总氮(TN)和总磷(TP)浓度不同,A 至F 总氮浓度分别设置为0.1、0.2、1.0、2.0、5.0、0 mg/L;总磷浓度分别设置为0.01、0.02、0.10、0.20、0.50、0 mg/L。每种生态箱设置6 个离子浓度水平,A1 至F1 生态箱Ca2+浓度设定为80 mg/L,A2 至F2 生态箱HCO3-浓度设定为4 mmol/L,A3至F3生态箱Ca2+浓度设定为60 mg/L,HCO3-浓度设定为3 mmol/L,A4 至F4 生态箱Ca2+浓度设定为80 mg/L,HCO3-浓度设定为4 mmol/L,A5至F5 生态箱Ca2+浓度设定为100 mg/L,HCO3-浓度设定为5 mmol/L,A6 至F6 生态箱(对照)均为自来水。
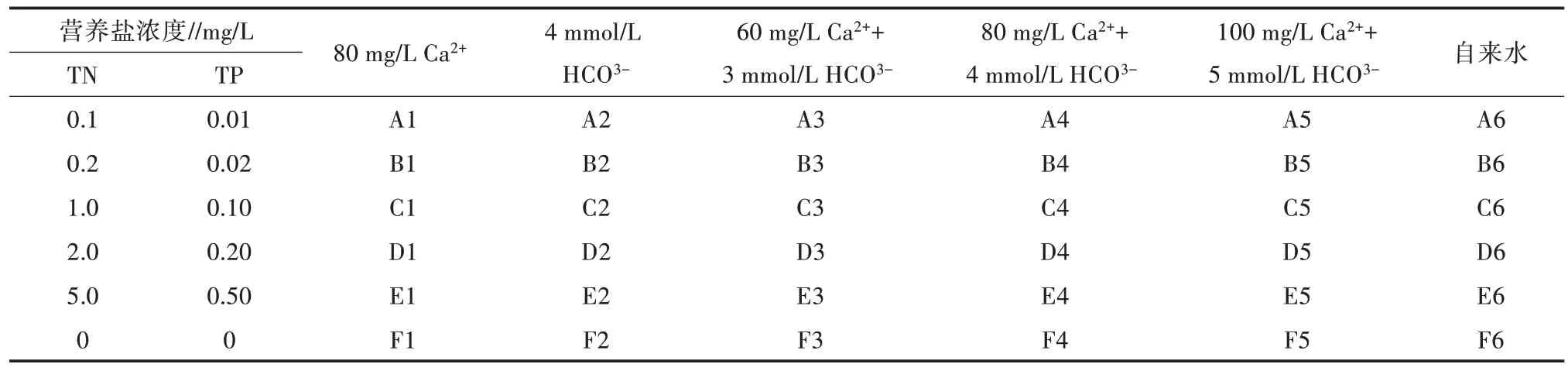
表1 生态箱营养盐浓度梯度和Ca2+、HCO3-浓度设定
培养液所用的储备液分别由CaCl2、NaHCO3、KNO3、KH2PO4配制而成。试验在室内进行,平均温度为18~25 ℃。检测生态箱中叶绿素a 的含量,每组重复3 次。试验为期90 d,从30 d 开始第一次检测,随后每隔7 d 检测1 次。
1.3 水样收集与检测
叶绿素是植物光合作用中重要的光合色素,通过测定水体中的附着藻类叶绿素a 质量浓度可代表生态箱中附着藻类的生物量,每次测量时均使用软毛刷将藻类洗刷下来,然后测定其质量浓度。水中TP 测定采用钼锑抗分光光度法(GB11893—89),水体TN 测定采用碱性过硫酸钾消解紫外分光光度法(GB 11894—89),叶绿素a 质量浓度测定采用《水和废水检测分析方法》中分光光度法;Ca2+、HCO3-浓度采用德国Merck 公司生产的便携式试剂盒测量,精度分别为2.0 mg/L 和0.1 mmol/L。
2 结果与分析
2.1 灵水湖HCO3-、Ca2+浓度的变化
对灵水湖的监测数据(图2)分析可知,Ca2+和HCO3-的年平均浓度水平呈缓慢上升的趋势,其中,2011 年Ca2+平均浓度约为61.7 mg/L,2015 年为76.5 mg/L,2016 年则逐渐上升至78.0 mg/L;2011 年HCO3-平均浓度为140.0 mg/L,2016 年上升为239.1 mg/L。分析其原因可能与灵水湖岩溶水动力系统的稀释效应、CO2效应及水动力效应(扩散边界层效应)有关[28]。
2.2 添加Ca2+对Chl-a 质量浓度的影响
通过分析生态箱附着藻类Chl-a 质量浓度进而探究添加Ca2+对不同TN、TP 营养盐浓度下Chl-a 质量浓度的影响,其中,试验组生态箱Ca2+浓度为80 mg/L,对照组生态箱未额外添加Ca2+,结果(图3)表明,随着TN、TP 营养盐浓度的提升,Chl-a 质量浓度随之上升,且添加Ca2+处理的Chl-a 质量浓度明显高于对照。在TN、TP 浓度分别为0.2、0.02 mg/L 的水平下,添加Ca2+的生态箱中Chl-a 质量浓度在7 次测量中均高于未添加Ca2+的对照生态箱;在TN、TP 浓度分别为1.0、0.10 mg/L 的水平下,前5 次测量的Chl-a 质量浓度均高于对照,后2 次测量的Chl-a 质量浓度均稍低于对照;在TN、TP 浓度分别为5.0、0.50 mg/L 的水平下,对照与处理的Chl-a 质量浓度总体水平相当。

图3 添加Ca2+对不同TN、TP 营养盐浓度下Chl-a 质量浓度的影响
钙元素是植物生长的必需营养元素,能够有效影响植物体内碳水化合物的形成与转化,从而影响植物的生长进程,已有研究表明钙元素可以增强凝集素的活性,这与微囊藻群落的形成息息相关[29]。在浅水湖泊中轮藻可以通过钙化作用缓解氨氮对轮藻生长的毒害作用,且轮藻叶表面可形成钙磷复合物,最终对水体的磷循环产生了重要影响[30]。
如图4 所示,图4A(第45 天)、图4B(第90 天)为添加80 mg/L Ca2+的试验水体与未添加80 mg/L Ca2+的空白对照水体中Chl-a 质量浓度的变化情况。试验水体中Chl-a 质量浓度与对照之间差异显著,整体来看,在相同TN、TP 营养盐浓度条件下,添加Ca2+在一定程度上可促进水体生态系统附着Chl-a 质量浓度的提高。

图4 添加Ca2+对Chl-a 质量浓度的影响
2.3 添加HCO3-对Chl-a 质量浓度的影响
如图5 所示,添加4 mmol/L HCO3-的生态箱,在TN、TP 浓度处于较低水平时,各试验处理附着藻类中Chl-a 质量浓度整体上明显高于未添加HCO3-的对照生态箱,在TN、TP 浓度分别为0.2、0.02 mg/L 和1.0、0.10 mg/L的生态箱中,含有HCO3-的水体中Chl-a 质量浓度上升迅速,但是二者在第5 次的测量中Chl-a 质量浓度维持较为平稳。在TN、TP 浓度较高的梯度中,即TN、TP 浓度分别为5.0、0.50 mg/L时,水体中Chl-a 质量浓度并未明显高于对照生态箱。
中国西南地区岩溶系统具有岩石可溶性特点,主要由CaMg(CO3)2组成的碳酸盐岩是全球最大的碳库[31]。植物能从HCO3-中获取碳源,开辟新的碳循环途径及光合作用途径[32]。已有研究表明,植物中碳酸酐酶能将细胞内的HCO3-转化为CO2和H2O,开启了利用可溶性无机碳的新途径[33],改变了碳源来源于水气交换所捕获CO2的传统认知。在高碳酸氢盐的岩溶水体中,附着藻类对营养盐的添加更敏感,相对低含量的营养盐浓度会造成附着藻类的大量增殖[34]。
图6A(45 d)、图6B(90 d)分别为添加4 mmol/L HCO3-的生态箱(试验生态箱)与未添加HCO3-的对照生态箱水体中Chl-a 质量浓度的变化情况。总体而言,在2 个时间段的检测中,试验生态箱与对照生态箱水体中的Chl-a 质量浓度存在显著性差异。在45 d 时的检测中,添加4 mmol/L HCO3-的生态箱水体中(除TN、TP 浓度分别为0.1、0.01 mg/L 的生态箱外)附着Chl-a 质量浓度显著高于对照生态箱,表明添加HCO3-能够促进附着藻类Chl-a 质量浓度的增加。在90 d 时的检测中,除TN、TP 浓度分别为0.2、0.02 mg/L 以及5.0、0.50 mg/L 时对照生态箱水体中Chl-a 质量浓度高于试验生态箱外,其余试验生态箱水体中Chl-a 质量浓度均高于对照生态箱。

图6 添加HCO3-对Chl-a 质量浓度的影响
2.4 添加Ca2+和HCO3-对不同氮磷浓度水平下Chl-a 质量浓度的影响
上述研究表明,生态箱中TN、TP 营养盐浓度相同时,不同生态箱中分别添加Ca2+或HCO3-可促进附着藻类Chl-a质量浓度的上升。设置Ca2+和HCO3-浓度分别为100 mg/L和5 mmol/L、80 mg/L 和4 mmol/L、60 mg/L 和3 mmol/L,探究同时添加Ca2+或HCO3-对不同氮磷浓度水平下Chl-a 质量浓度的影响。结果(图7)表明,随着添加Ca2+和HCO3-浓度的上升,相同营养盐浓度水平下的Chl-a 质量浓度逐渐上升,同时随着营养盐浓度的提高,Chl-a 质量浓度也呈上升趋势。

图7 Ca2+和HCO3-不同浓度梯度水平下Chl-a 质量浓度的变化情况
本试验设计中还存在不足之处,生态箱中设计的TN、TP 营养盐浓度虽包含了国内多数地表水由低至高的浓度梯度,但设计中TN 与TP 的浓度比均为10,呈现出一定的单一性。目前针对水体富营养化氮磷控制策略方面,已有不少对藻类生长限制因子的研究,主流结论为磷控制[35]、氮控制[36]、氮磷双控制[37]学说。由于水体生态系统存在时间和空间异质性的特点,不同氮磷比营养浓度条件下的岩溶水对附着藻类的影响还需进一步研究。
图8A(45 d)、图8B(90 d)为不同Ca2+和HCO3-浓度梯度下试验生态箱与对照生态箱水体中Chl-a 质量浓度的变化情况。整体而言,随着Ca2+和HCO3-浓度的升高,Chl-a 质量浓度上升,不同Ca2+和HCO3-浓度梯度水平下Chl-a 质量浓度存在着显著性差异,当生态箱Ca2+和HCO3-浓度分别为100 mg/L、5 mmol/L时,Chl-a 质量浓度最高。在45 d 模拟试验中,在TN、TP 浓度分别为0.1、0.01 mg/L 和0.2、0.02 mg/L时,Chl-a 质量浓度差异不明显,其余生态箱之间均差异显著;在90 d 模拟试验中,TN、TP 浓度分别为0、0 mg/L 和0.1、0.01 mg/L时,Chl-a 质量浓度差异不明显,其余生态箱之间均差异显著。

图8 不同Ca2+和HCO3-浓度梯度水平下模拟不同天数时Chl-a 质量浓度的变化
3 小结与展望
本研究结果表明,在为期90 d 的模拟岩溶水生态系统中Ca2+和HCO3-分别对不同营养盐浓度梯度下附着藻类叶绿素a 质量浓度产生显著影响。总体而言,在添加Ca2+的生态箱水体中,Ca2+的添加促进了相同N、P 营养盐浓度水平下藻类叶绿素a 质量浓度的上升;在添加HCO3-的生态箱水体中,HCO3-的添加促进了相同N、P 营养盐浓度水平下藻类叶绿素a 质量浓度的上升;在Ca2+和HCO3-同时添加的生态箱水体中也显著促进了附着藻类的生长。
尽管目前灵水湖水质能较好地满足水源地水质的要求,但在特殊的岩溶水环境下,相同的N、P 浓度条件下高氮高钙的岩溶水环境更加有利于附着藻类的生长,附着藻类生物量激增在一定程度上通过生态位的竞争可抑制沉水植物的生长,根据生态系统的灾难性突变理论,一旦营养盐水平在短时间内超过了生态系统稳态转换的阈值,那么灵水湖原有的草型稳态很有可能在短时间内系统垮塌,逐渐转向以藻类为主的浊水稳态,推测附着藻类对沉水植物的竞争是灵水湖岩溶生态系统灾难性转变的重要因素。
在水体系统的复杂营养链级和外界干扰下,若使沉水植被的修复达到稳定和平衡的状态,仍存在一定的困难,实际水体修复工程的实施目前尚未形成较为完善的方法及体系,沉水植物修复仍需要大量野外观测及原位修复工程的实施,进而总结经验并推广。
猜你喜欢
草业学报(2022年5期)2022-07-08
中国医学物理学杂志(2021年10期)2021-11-04
草业学报(2021年3期)2021-03-22
哈尔滨轴承(2020年2期)2020-11-06
花卉(2020年12期)2020-01-09
湖南大学学报·自然科学版(2019年6期)2019-07-26
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
草业学报(2018年10期)2018-10-19
电镀与环保(2017年5期)2017-12-19
大科技(2016年1期)2016-07-17
