
白桦BpAMT基因家族鉴定及表达模式分析
2023-04-29 00:44:03杨海昕刘晓莹詹亚光范桂枝
林业科学研究 2023年2期
关键词:白桦
杨海昕 刘晓莹 詹亚光 范桂枝
关键词:白桦;AMT;基因家族;基因表达模式
氮素是植物生长发育过程中所必需的大量营养元素之一,是核酸、蛋白质和叶绿素等大分子物质的重要组成成分。作为植物吸收无机氮素的低耗能方式,铵态氮的吸收利用效率直接影响植物生长发育,吸收过量的铵对植物具有毒性,其吸收和代谢受到严格的调控。
铵态氮的吸收主要由铵根转运蛋白(Ammoniumtransporter,AMT)调控,它将铵根离子从细胞外转运到细胞内进而调节植株地上部分与地下部分的氮素平衡。随着测序技术和生物信息学的发展,研究者发现,基因AMT一般包括AMT1和AMT2两个亚家族,亚家族成员之间具有相似的高级结构,但其基因结构和成员数量不同。在基因结构上,AMT1家族成员一般不含内含子,多为高亲和转运体;AMT2家族成员含多个内含子且位置和大小基本保守,其亲和性没有明确规律;在基因数量上,AMT家族成员一般在6~16个之间。在基因表达上,AMT家族成员在根、茎和叶等的基因表达具有组织部位特异性,同时也受病毒和害虫等生物以及氮素浓度、光照、植物激素和干旱等非生物因素的调控,如水稻(Oryza sativaL_)OsAMT1.3在根系维管束和侧根原基中表达,百脉根(Lotusjaponicas Regel.)LjAMT2.1主要在根瘤细胞的原生质膜上表达,柑橘(Citrus reticulata Blanco.)CitAMT1和棉花(Gossypium hirsutum L_)GhAMT1.3在低氮的条件下表达量大幅度提高,拟南芥(Arabidopsis thaliana L、)AtAMT1.1及转录水平随日变化周期中光照强度的降低而下降。
在自然状态下,土壤中的氮素受到限制或有效性降低都会限制植株的生长、影响植株对碳同化物质的分配格局以及抵抗胁迫的能力。白桦(Betula platyphyHa Suk.)是喜光耐寒的先锋树种,它对土壤适应性强且生长速度较快。研究发现,在一定浓度范围内随着供氮水平的增加,白桦幼苗根系、茎和口十中全氮浓度提高,其中,无机氮比有机氮更易于其生物量的积累。AMT作为氮代谢的关键基因之一,其家族成员在白桦中的研究还未见报道,尤其是其成员数量及功能还不清楚,有关AMT基因在非生物胁迫条件下表达的研究文献有限。因此,本研究以10年生白桦树、2年生白桦盆栽苗及白桦悬浮细胞为试验材料,基于最新公布的白桦基因组序列,筛选鉴定BpAMT家族成员,初步解析BpAMT家族成员在不同组织、不同氮素条件、日变化周期以及不同激素和非生物胁迫处理后的表达模式,该研究将为深入探究BpAMT家族成员的功能提供参考依据。
1材料与方法
1.1材料
白桦组培苗与悬浮细胞为实验室保存材料,组培苗培养基为WPM培养基附加蔗糖20g.L-1与琼脂6.5g.L-1,每25~30d继代1次;悬浮细胞培养基为B5培养基附加蔗糖20g.L-1、琼脂8g.L-1、水解酪蛋白1g.L-1、TDZ0.6mg.L-1、6-BA 0.3mg.L-1与2,4-D0.5mg.L-1.每7d继代1次;10年生白桦树材料采集于东北林业大学主校区丹青楼前白桦林;2年生白桦盆栽苗培养于东北林业大学逸夫教学楼旁。
1.2方法
1.2.1生物信息学分析
利用拟南芥AtAMT1.1(CAA53473). AtAMT1.2 (AAD54639). AtAMT1.3(AAD54638). AtAMT1.4 (CAB81458). AtAMT1.5(NP_189072)和AtAMT2.1(NP_181363)6个AMT序列,在白桦基因组数据中进行比对,数据库网址为https://www.ncbi.nlm.nih.gov/genbank,登录号为PRJNA285437,剔除重复序列后获得BpAMT候选基因,进行保守结构域鉴定,确定BpAMT家族成员并进行编号。通过在线分析软件EXPASY进行理化性质分析;通过在线软件WoLFPSORT进行亚细胞定位预测;使用MEGAX构建系统进化树,构树方法选择NJ法;使用GSDS2.0绘制基因结构,使用在线MEME对白桦目标氨基酸序列进行比对,并对保守区进行分析;通过在线软件MG2C绘制基因在染色体上位置;使用TBtools进行共线性分析;运用SPDEv2.0确定启动子序列,plantcare进行顺式作用元件功能预测,导人TBtools进行可视化。
1.2.2表达模式分析
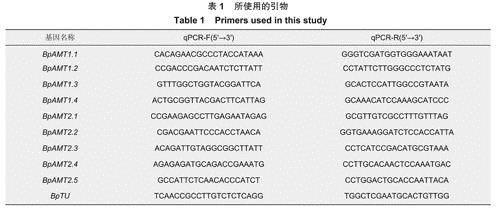
1.2.2.1组织部位表达模式分析于2021年7月15日采集2年生白桦盆栽苗的根、茎和叶组织,每次取材进行3次生物学重复,测量不同组织下BpAMT基因表达情况,采用改良的CTAB法提取总RNA,使用Takara公司ScriptTM RT试剂盒合成cDNA。使用Premier5.0设计定量引物(表1),引物由生工生物工程上海股份有限公司合成,以白桦微管蛋白基因BpTu为内参基因,进行qRT-PCR分析基因表达模式,目的基因的相对表达量用:表示。运用IBM SPSS Statistics 23软件对BpAMT基因表达进行统计及显著性差异分析。
1.2.2.2不同氮素处理下表达模式分析
以含不同氮源的B5培养液处理白桦悬浮细胞,设置对照CK组(正常B5培养液),CKO组(CK组去除NH4N03试剂)、低浓度单一硝态氮组(CKO组添加0.1 mmoI.L-1KN03试剂)、高浓度单一硝态氮组(CKO组添加3mmoI.L-1KN03试剂)、低浓度单一铵态氮组(CKO组添加0.1mmoI.L-1NH4CI试剂)与高浓度单一铵态氮组(CKO组添加3mmoI.L-1NH4CI试剂)为实验组,分别培养白桦悬浮细胞,每个处理进行3次生物学重复,在1d后取样测量不同处理下BpAMT基因表达量的变化情况,并进行统计及显著性差异分析。
1.2.2.3日变化表达模式分析于2020年8月3-4日1个日周期内,每隔3h取样1次,共9个时间点采集主校区丹青楼前10年生白桦树叶片,每次取材进行3次生物学重复,对不同时间下BpAMT基因表达量进行统计分析。
1.2.2.4不同激素处理下表达模式分析选择长势一致的白桦盆栽苗,分别用100umoI.L-1茉莉酸甲酯(MejA)、100mg.L-1赤霉素(GA3)和10umoI.L-1脱落酸(ABA)处理24h后取样,每次取材进行3次生物学重复,测量不同激素处理下BpAMT基因表达量的变化情况。
1.2.2.5不同非生物胁迫下表达模式分析选择长势一致的白桦盆栽苗,分别用50mmoI.L-1氯化镉(CdC12)、10%(W/V)PEG6000和4℃环境模拟重金属、干旱和低温胁迫,处理24h后取样,每次取材进行3次生物学重复,测量不同非生物胁迫下BpAMT基因表达量的变化情况。
2结果与分析
2.1BpAMT家族成员鉴定及基本特征分析
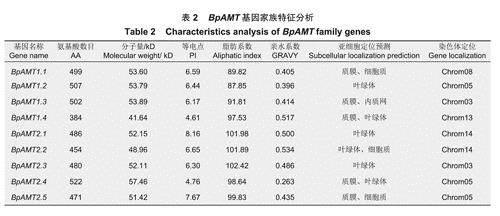
从白桦基因组中筛选鉴定了9个BpAMT基因,分别编号为BpAMT1.1~1.4、BpAMT2.1~2.5(表2)。对BpAMT蛋白理化性质分析表明:氨基酸残基数范围为384~522,分子量范围为41.64~57.46kD,蛋白质等电点范围为4.61~8.16。BpAMT蛋白亲水系数均为正值,均为疏水性蛋白。此外,亚细胞定位结果显示:BpAMT主要定位于膜结构上,多分布在叶绿体、质膜与内质网上,这可能与BpAMT蛋白的跨膜转运功能有关。BpAMT基因成功定位于第3、5、8、13和14条染色体上,其中,第5号染色体上分布基因最多,有3个基因,且均分布在染色体下端。第8号和13号染色体上基因分布最少,均有1个基因。BpAMT2.1和BpAMT2.2在第14号染色体上的位置接近,并且二者的序列长度和基因结构相似,可能是BpAMT中的串联复制基因。
2.2BpAMT系统发育分析
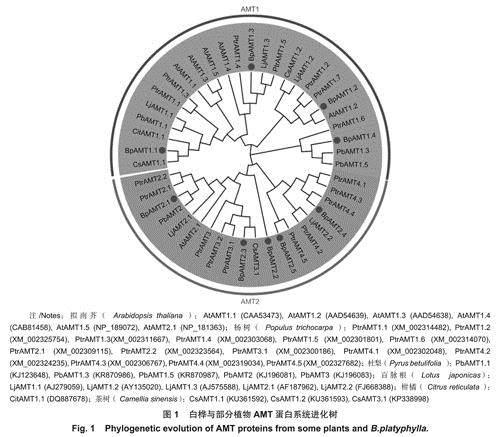
如图1所示,将白桦BpAMT与拟南芥、柑橘、百脉根、杨树(Populus trichocarpa Torr.&Gray)、茶树(Camellia sinensis L)和杜梨(Pyrusbetulifolia Bunge.)AMT氨基酸序列构建进化树。结果显示:BpAMT家族分为BpAMT1与BpAMT2两个进化支,在9个BpAMT基因中,其中4个属于BpAMT1亚家族,5个属于BpAMT2亚家族。其中BpAMT1.4与BpAMT1亚家族中其余3名成员亲缘关系较远,与杜梨PbAMT1.3、PbAMT1.5及杨树PtrAMT1.6聚类在一起。白桦BpAMT与杨树PtrAMT聚类最多,亲缘关系较近。
2.3基因结构与保守基序分析
为了了解BpAMT基因的序列特征,对其基因结构及保守基序进行分析。图2A表明:BpAMT1.1~1.3均含有—个1个外显子,无内含子;BpAMT1.4含有4个外显子与3个内含子;BpAMT2基因序列包含3~6个外显子与3~5个内含子,且具有未翻译区结构。
BpAMTs保守基序见图2B,BpAMT两个亚家族中均存在motif 4、5、6、7、9这5种保守基序。BpAMT2亚家族成员氨基酸序列中含有特定识别基序motif3,除BpAMT1.4外,BpAMT1亚家族成员氨基酸序列中含有特定识别基序motif10,motif 7在两个亚家族中都存在,但是位置有很大差异,motif 7在BpAMT1的5端,而在BpAMT2的中段。BpAMT的保守性较高,同一个亚家族的BpAMT蛋白motif分布较相似,这些保守motif的存在保证了家族成员在转运铵盐中发挥稳定的作用。
2.4顺式作用元件分析
提取BpAMT基因编码区上游2000bp的核苷酸序列,根据BpAMT基因家族的性质,去除一般性转录调控元件和功能未知元件,图3表明:发挥功能的元件主要分为3类:逆境胁迫响应元件、光响应元件及植物激素响应元件,从中选取了10种重要的顺式元件进行可视化。
除BpAMT1.3外,其余BpAMT启动子区具有1~2个逆境响应元件,包括TC-rich repeats元件、响应干旱胁迫的MYB元件以及响应低温胁迫的LTR元件,说明这些成员与逆境应答密切相关。BpAMT启动子区的光反应元件主要为G-box元件,除BpAMT1.2与BpAMT2.5不包含光反应元件外,其余BpAMT启动子区具有1~3个光响应元件,BpAMT2.3启动子区具有参与昼夜节律调控的元件。BpAMT启动子区激素响应元件种类较多,主要有参与脱落酸反应的ARBE元件、响应生长素的TGA-box元件、参与水杨酸反应相关的TGA-motf元件以及响应赤霉素的TATC-box元件,表明BpAMT可能受到各种不同激素的调控。以上结果表明,BpAMT可能与逆境应答、激素信号转导和生长发育过程密切相关。
2.5BpAMT的组织部位表达模式分析
如图4所示,BpAMT家族基因在组培苗和盆栽幼苗根、茎和叶中的表达趋势相同,均呈现叶>根>茎趋势,其中,在盆栽苗和组培苗茎和叶部位BpAMT1.1的表达量最高,分别为153.05和377.41、267.88和253.06;在盆栽苗和组培苗根、茎、叶中BpAMT1.3的表达量均最低,分别为9.35、2.14、1和1、12.18、2.19。
2.6不同氮素条件下BpAMT的表达模式分析
如图5所示,BpAMT家族基因在硝态氮和铵态氮处理后的表达趋势不同。在去除培养基中无机氮源(CKO)的条件下,除BpAMT2.3之外的BpAMT表达量均呈上调趋势;在KN03与NH4CI处理下,BpAMT1.1和2.1表达量均呈上调趋势,其余BpAMT在KN03处理下均呈下调趋势,其余BpAMT1与BpAMT2在NH4CI处理下呈现相反的变化趋势,其中,BpAMT2.1表达量在CKO、3mmoI.L-1 KN03.0.1mmoI.L-1NH4CI与3mmoI.L-1NH4CI处理下均呈上调趋势,表达量分别为对照组的19、58、50和18倍。BpAMT1.3和1.4的表达量在不同氮素条件下的表达都比较低,这可能与其较大的基因家族有关系,表明家族基因之间存在功能冗余。
2.7BpAMT的日变化表达模式分析
如图6所示,BpAMT1.1、1.3、2.1、2.2和2.3表达量在3:00-15:00时段呈上升趋势,在18:00-0:00时段呈下降趋势,BpAMT1.4和2.4的日变化规律与之相反。不同基因表达量达峰值的时间不同,BpAMT1.1和1.3表达量在18: 00达到峰值,BpAMT2.1、2.2和2.3表达量在15: 00达到峰值,BpAMT1.4表达量在00:00达到峰值,BpAMT2.4表达量在3:00达到峰值。
2.8BpAMT在激素处理下的表达模式分析
如图7所示,BpAMTs在MejA、GA3和ABA处理下呈现不同的表达特征。经MejA处理后,除BpAMT1.3和BpAMT2.1外,其余7个BpAMT的表达量上调,其中,BpAMT1.4表达量在根部上调最明显,为对照组的4.18倍,BpAMT2.7表达量在根部下调最明显,为对照组的0.14倍。经ABA处理后,BpAMT1.1、2.1、2.2和2.5的表达量上调,其余5个BpAMT的表达量下调,其中,BpAMT2.7表达量在Ⅱ十片中上调最明显,为对照组的6.01倍,BpAMT1.3表达量在茎部下调最明显,为对照组的0.18倍。经GA3处理后,BpAMT1.1、2.2和2.4的表达量上调,BpAMT2.5的表达量下调,其余5个BpAMT的表达量没有显著变化,其中,BpAMT2.2表达量在根部上调最明显,为对照组的6.17倍,BpAMT2.5表达量在叶片下调最明显,为对照组的0.41倍。在MejA、GA3和ABA处理下,BpAMT1.1和2.2在叶、茎和根中均呈现上调表达趋势。以上结果表明,BpAMT受多种激素的诱导表达,但对不同激素的响应程度与响应部位存在差异。
2.9BpAMT在非生物胁迫下的表达模式分析
如图8所示,BpAMT在低温、干旱和氯化镉处理下呈现不同的表达特征。经低温处理后,BpAMT1.2、7.4、2.2、2.4和2.5的表达量上调,BpAMT1.1和2.3的表达量下调,其中,BpAMT2.4在茎部上调最明显,为对照组的12.25倍,BpAMT1.1在根中下调最明显,为对照组0.13倍。经干旱处理后,BpAMT1.1、1.4、2.1和2.3的表达量上调,BpAMT1.2、1.3、2.2和2.5的表达量下调,其中,BpAMT1.4在茎部上调最明显,为对照组的9.97倍,BpAMT2.5在茎部下调最明显,为对照组的0.13倍。经氯化镉处理后,BpAMT2.1和2.4的表达量上调,BpAMT2.2和2.3的表达量没有显著变化,其余BpAMT的表达量下调,其中,BpAMT2.4的表达量在茎中上调最明显,为对照组的9.74倍,BpAMT1.3在茎中下调最明显,为对照组的0.09倍。在低温、干旱和氯化镉处理下,BpAMT2.1在叶、茎和根3个组织部位中均呈现上调表达趋势。以上结果表明,BpAMTs在不同非生物胁迫中的表达情况存在差异,推测在逆境中发挥不同的作用。
3讨论
铵是植物根系吸收的经济有效的一种氮素形式,AMT介导的高亲和力铵跨质膜运输是植物根系吸收铵的主要途径。近年来,有关植物吸收和转运铵态氮的分子机制有了较大突破,特别是随着高通量测序技术的发展,拟南芥、谷子(Setariaitalica Beauv.)、玉米(Zea maysL_)及甘蔗(Saccharum officinarum L-)等植物的AMT遗传信息逐步被发掘。
白桦基因组数据的公布为系统解析BpAMT家族成员提供了可能。因此,本论文基于白桦基因组和本研究室获得的转录组数据,共筛选了9个具有完整Ammonium-transp superfamily结构域的BpAMTs,与已报道的植物AMT家族分为AMT1和AMT2两个亚家族一致,9个BpAMTs也分为两类,其中,4个属于BpAMT1亚家族,5个属于BpAMT2亚家族。
BpAMT2亚家族成员的氨基酸序列中均具有特定识别基序motif 3,保守基序的数量和位置相同,其基因结构中外显子数目在3~6个之间,这与葡萄(Vitis vinifera L.)、水稻与百脉根等植物AMT2亚家族成员的报道一致; BpAMT1亚家族成员BpAMT1.1~1.3的氨基酸序列中保守基序的数量和位置相同,均含有特定识别基序motif10,基因结构中不含内含子,这也与大麦(Hordeum vulgare Ll)和杨树等植物中大多数AMT1基因不含内含子的报道一致。BpAMT1.4基因结构中包含3个内含子,其氨基酸序列中缺失motif8和BpAMT1亚家族特定识别序列motif10,但其它保守基序的数目和位置都与BpAMT1.1~1.3相同;进化关系分析发现,BpAMT1.4与BpAMT1.1~1.3定位在同一分支上,故将其归类于BpAMT1亚家族。BpAMT1.4氨基酸序列中缺失motif10可能与其结构演化过程中内含子的获得有关,同时推测其可能参与特殊的铵转运机制。
基因家族的进化过程中会出现基因复制、扩展、丢失和漂移等事件,BpAMTs也出现了基因片段复制事件,且不均匀分布于5条染色体上。由此推测,为了适应不同生境下的氮素种类及供给量,白桦在进化过程中可能通过扩展BpAMT基因家族来适应环境。启动子顺式式作用元件分析显示:BpAMTs含有多个胁迫响应元件、激素响应以及光反应元件等,表明BpAMTs基因潜在的重要功能。同一亚家族内成员含有的元件类型和数量也各不相同,表明BpAMTs的表达调控方式具有一定的特异性。
植物AMT基因的表达具有组织特异性,如拟南芥在根中表达最高,在茎中表达最高,AtAMT1.4在花粉中专一性表达。同样,在2种白桦材料中对BpAMTs的组织部位表达模式进行了验证,不同组织部位的BpAMTs表达趋势均为叶>根>茎。由此推测,BpAMTs基因表达主要受叶片中系统氮态信号以及根系中局部铵态信号的控制。在响应光照的日变化中,菜心BcAMTs基因表达量在6:00-12: 00为上升趋势,15:00-00:00呈下降趋势,在中午12:00时表达量达到峰值。同样,BpAMT1.1、1.3与BpAMT2.1~2.3的在同一日周期内的表达量也有相似的趋势,而BpAMT1.4、BpAMT2.4日变化趋势与之相反,表明在光照逐渐减少的情况下,白桦通过提高BpAMT1.4、BpAMT2.4基因的表达,维持植物夜间的铵根离子吸收。由此可见,BpAMTs表达受日变化光周期的影响,而铵根离子吸收速率是否具有日变化趋势还需进一步验证。
一般情况下,缺氮或低氮条件对铵根离子的吸收有促进作用,不同的氮源对铵根离子的吸收也有影响,硝酸根作为氮代谢过程中氮元素吸收同化的源头,可以在酶促反应下合成铵根离子,作为铵根离子转运基因,BpAMTs表达的规律性有待明确。BpAMTs在不同氮素处理下呈现不同的表达特征。在缺氮条件下,除BpAMT2.3外,其余BpAMTs表达量均上升,这与柑橘、地钱(Marchantiapolymorpha L-)及小麦[29]在缺氮后AMT基因表达趋势相似;在高浓度单一硝态氮为氮源的条件下,BpAMT2.1、1.1与2.5表达量显著提高,推测这可能与高浓度单一硝态氮对悬浮细胞产生了胁迫有关系,使之趋向于吸收耗能更低的铵根离子。
研究表明,氮素形态影响植物体内部分激素的合成和运输,同时通过外施激素也可以增加植物的氮素利用率。外源激素是影响AMT基因表达的重要因素,MejA和ABA处理杜梨后,发现PbAMT1.5的表达显著上调; GA3和NAA处理毛竹后,多个PeAMTs显著上调,PeAMT4显著下调。BpAMT对激素的响应存在差异,BpAMT1.1、2.1、2.2、2.4和2.5对MejA、GA3和ABA处理均具有敏感性,其中,BpAMT1.1和2.2在叶.茎和根中均呈现上调表达趋势,且在根部响应最强烈,推测白桦根部铵根离子的吸收过程与激素互相作用,共同调节白桦的根系发育。
非生物胁迫是影响植物铵根吸收转运的重要因素。干旱胁迫下楸子(Malus prunifolia Borkh.)MpAMT1.2和4.2的表达显著上调;茶树在盐胁迫下CsAMT1.3b和3.1表达显著下调,CsAMT1.1a、2.1a和3.3表达显著上调。BpAMT对非生物胁迫的响应存在差异,BpAMT1.1、1.2、2.1和2.5对低温、干旱和镉胁迫处理均具有敏感性,其中,BpAMT2.1在叶、茎和根3个组织部位中均呈现上调表达趋势,且在茎部响应最强烈,可能由于胁迫处理改变了不同组织部位对铵态氮的利用度,打破了铵根离子在根与茎之间运输过程的动态平衡,导致BpAMT在茎部的表达改变。镉胁迫下的下调基因数量最多,可能因为NH4+与Gd2+同为阳离子,白桦通过抑制铵根离子的吸收降低植物对Cd2+的吸收,对植物本身起保护作用。由此认为,BpAMT基因可能通过在逆境环境中调节铵态氮的吸收与转运,从而保证氮代谢的稳定性。以上结果为白桦氮素吸收代谢与激素以及非生物逆境之间的互作研究提供了参考思路。
4结论
从白桦基因组数据库中鉴定了9个BpAMT家族成员,定位到白桦5条染色体上,根据进化关系和结构特征分为2个亚家族。BpAMT基因表达具有组织部位特异性,受日变化、不同氮素、激素和非生物胁迫等影响,其中,BpAMT1.1与BpAMT2.1响应较强。以上结果为BpAMT家族成员的功能研究提供了有价值的信息和可靠的候选基因。
猜你喜欢
《学习方法报》小学语文四年级(2023年26期)2023-04-29 00:44:03
作文周刊·小学四年级版(2022年8期)2022-03-11 07:42:32
知音(月末版)(2020年5期)2020-07-04 12:33:23
作文周刊·小学四年级版(2020年8期)2020-05-08 12:57:05
文苑(2019年20期)2019-11-20 02:12:33
文苑·经典美文(2019年10期)2019-10-16 02:47:16
环球人物(2019年3期)2019-02-27 08:47:02
天然产物研究与开发(2016年6期)2016-06-05 10:29:26
小天使·二年级语数英综合(2016年8期)2016-05-14 13:03:38
华人时刊(2016年16期)2016-04-05 05:57:20
