
驯化对谷子-根际微生物互作机制影响的进展与展望*
2023-04-22 10:44崔文秀宰小玉赵美丞郑春燕宋春旭
中国生态农业学报(中英文) 2023年4期
崔文秀,宰小玉,赵美丞,郑春燕,宋春旭,4**,朱 峰**
(1.河北省土壤生态学重点实验室/中国科学院农业水资源重点实验室/中国科学院遗传与发育生物学研究所农业资源研究中心石家庄 050022; 2.中国科学院大学 北京 100049; 3.植物-土壤相互作用教育部重点实验室/中国农业大学资源与环境学院/中国农业大学国家农业绿色发展研究院 北京 100193; 4.河北曲周农业绿色发展国家野外科学观测研究站 曲周 057250)
在人类文明发展进程中,作物驯化发挥着不可忽视的推动作用,其不仅保证食物的可持续供应,而且促进了定居农业社会的形成[1-2]。作物驯化是指人类以满足自身需要为目的,为野生物种提供适宜的生长发育条件,并对野生物种的某些性状进行选择、修饰和利用[3-5],从而获取满足人类所需用途的农艺性状的过程。作物驯化起源于12 000年前,考古证据表明,作物具有不同的驯化中心,例如小麦(Triticum aestivum)起源于土耳其,玉米(Zea mays)和大豆(Glycine max)起源于墨西哥,以及水稻(Oryza sativa)和谷子(Setaria italica)起源于中国等[5-7]。植物驯化过程往往伴随着少数基因的突变,其中,与产量和适口性相关的基因逐渐被保留下来,在一定程度上降低了作物的遗传多样性,使得性状表型趋于一致,这种现象被称为“驯化综合征”[1,4-5,7-8]。例如,在漫长的驯化过程中,驯化增加了作物籽粒/果实产量,改善了植株形态和营养物质组成,改变了根系形态或次生代谢产物组成[9-11],进而引起作物根际微生物群落结构和功能的变化。目前,对于作物驯化的研究多关注植物遗传基因与植物表型和生理特性之间的关联机制,微生物在作物驯化过程中的作用及其与植物互作机制的探索仍处于起步阶段。
植物微生物组包括与植物广泛互作的微生物群落,是植物的第二基因组,其中,根际微生物是植物与土壤交互作用的媒介[12-13]。根际有益微生物有助于植物获取养分,促进植物生长发育,增强植物抗逆性,减少植物土传病害的发生。然而,根际有害微生物与植物竞争营养物质、产生毒素、导致植物感病,有碍植物健康生长[13-14]。不同种类作物中,驯化对其根际微生物组的影响存在差异[15-17],呈现微生物多样性减少[18]、共生相互作用减弱[19]、有益微生物菌群丰度降低等特征[17,20]。驯化导致的根际微生物群落变化是由植物根形态和分泌物变化所引起的。例如,野生菜豆(Phaseolus vulgaris)根际拟杆菌(Bacteroidetes)丰度与比根长(specific root length,SRL)和根密度显著相关[16-17]。植物通过根系分泌物和沉积物选择性地招募特定的和潜在的有益微生物,并且根系分泌物能够影响根际生物共生关系。例如,类黄酮能够提高植物对病原菌、寄生虫的抗性,对根瘤菌和菌根的共生起着促进作用[13-14]; 植物激素水杨酸影响拟南芥(Arabidopsis thaliana)根际微生物的组成[21]。相反,在对抗性品种的选育过程中,研究人员无意识地选择了更多可培养的有益根际微生物,例如,水稻抗条斑病品种‘CG2’根际可培养的优势菌群为芽孢杆菌(Bacillus),具有抑制条斑病的作用[22]。驯化过程中有益根际微生物的缺失与植物表型紧密关联,然而,驱动微生物群落组装和功能的潜在机制以及微生物组与植物特征相关性仍有待挖掘[13]。目前,对于驯化的研究大多关注作物农艺性状、作物基因演变与性状关联,对于驯化过程中作物-微生物互作的研究尚处于起步阶段,有待深入研究[21,23]。有关驯化-作物-微生物的研究大多集中在豆科(Leguminosae),对禾本科(Gramineae)作物探索相对较少,仅在水稻、大麦(Hordeum vulgare)等作物中有少数研究报道,而对于起源于中国的谷子研究更为缺乏[15-16,23]。
人类驯化的粮食作物主要有谷类、薯类、豆类。其中,谷类的种植最为广泛且绝大部分属于禾本科[24-25]。近年来,由于谷子具有基因组小、易于遗传转化及突出的抗逆性等特点,受到国际学术界的广泛关注,对其进行植物与微生物互作机制的探索,可以为其他禾本科植物的研究提供理论指导和依据[26-30]。考古学和遗传学证据表明,谷子起源于距今约8700年前的中国北方干旱半干旱地区,由青狗尾草(Setaria viridis)驯化而来[31-33]。谷子属于抗旱作物,具有耐瘠薄、水分利用效率高、适应性广、稳产性强、化肥农药需求量少等特点,是典型的环境友好型作物。在干旱形势日益严重的情形下,谷子具有重新成为主栽作物和主要粮食作物的潜力,挖掘谷子种植的潜在价值有利于应对日益严重的粮食危机[34]。谷子种植不仅在旱作农业可持续发展中具有潜力,也在调整饮食结构中具有重要意义。谷子不含麸质,其中籽粒79%的部分可被消化,含有丰富的淀粉、蛋白质、维生素、矿物元素、纤维等营养物质,品质优于小麦、玉米、水稻等主栽禾谷类作物,并且是杂粮作物中含酚酸类最高的作物[35-37]。此外,谷子还是粮饲兼用的作物,作为牲畜饲料在内蒙古等地广泛种植[38]。本文首先对驯化过程在禾本科作物根际微生物多样性及群落构建中的作用进行了回顾,随后重点针对起源于中国的重要旱作C4 模式作物谷子,系统总结了谷子的驯化过程及其对谷子生长、生理性状和根际微生物结构和功能的影响,旨在进一步推动驯化对谷子-根际微生物互作机制的研究,为恢复农田生态系统中的作物-微生物有益关系提供理论指导。
1 驯化对禾谷类作物根际微生物的影响
禾本科是一个庞大且分布广泛的单子叶开花植物家族,包括非常重要的作物类群。其中,至少有48个物种经过驯化,30 个以上的物种是重要粮食来源,如小麦、大麦、燕麦(Avena sativa)、水稻、玉米、高粱(Sorghum bicolor)、谷子等[39]。在驯化过程中,禾本科的基因型、性状演变等方面具有一定的共性,例如遗传多样性丧失、分蘖减少、落粒性和休眠性丧失等[39]。
驯化对禾本科作物根际微生物的影响存在差异。驯化显著影响水稻根际微生物群落结构与组成,野生稻(Oryzaspp.)根际细菌优势类群为厌氧绳菌纲(Anaerolineae)和硝化螺旋菌纲(Nitrospira),而栽培稻根际则富集了腐败螺菌纲(Saprospirae)微生物[15]; 野生稻根际真菌中的子囊菌门(Ascomycota)和担子菌门(Basidiomycota)的相对丰度高,而栽培稻根际优势类群为接合菌门(Zygomycota)真菌[40]。研究表明栽培稻根际细菌、真菌α 多样性高于野生稻[15,40-41],也有研究表明野生稻与栽培稻根际细菌α 多样性无显著差异,栽培稻根际真菌α 多样性高于野生稻,且驯化对水稻真菌群落结构影响更显著[42]。通过对比不同地点栽培稻和野生稻根际微生物群落多样性和稳定性差异发现,栽培稻的根际微生物群落对环境变化更敏感[40,43],表明野生稻稳定的根际微生物组成可以协助其抵抗胁迫。Shi 等[44]将稻瘟病菌(Magnaporthe oryzae)接种到栽培稻和野生稻的根际,发现病原菌导致栽培稻根际细菌和真菌多样性发生显著变化,栽培稻根际微生物群落对病原菌的响应更敏感、更易受干扰; 而在野生稻的根际病原菌丰度较低,有益细菌和真菌类群的相对丰度较高,抗逆性强。研究人员将稻瘟病菌接种到水稻根际后,通过扩增子和转录组分析发现,球囊菌门(Glomeromycota)能够促进野生稻中与苯丙烷和木质素合成以及脂肪酸降解有关的基因转录,进而提高了野生稻对稻瘟病菌的抗性[44]。
驯化对小麦根际微生物群落多样性的影响尚未得到一致性规律。例如,研究发现,野生小麦和栽培小麦根际微生物多样性无显著差异[45-47],而Hassani 等[20]研究发现与栽培小麦品种相比,野生小麦根际细菌群落的多样性更丰富,这与不同研究中选用的野生小麦和栽培小麦品种有关,亲缘关系的远近在一定程度上影响微生物群落,亲缘关系越近的品种根际微生物群落越相似[48-49]。不同驯化阶段的小麦根际富集的微生物类群存在差异,野生型根际细菌优势类群为厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes),栽培型根际细菌中变形菌门(Proteobacteria)和放线菌门(Actinobacteria)占比较多[20,48];野生二倍体小麦根际潜在的病原菌粪壳菌纲(Sordariomycetes)相对丰度低于驯化品种,其根际的共生体球囊菌纲(Glomeromycetes)相对丰度高于驯化品种[45]。同时,与育成品种相比,小麦地方品种根际具有更多特有微生物,如粪锈伞科(Bolbitiaceae)、大孢子科(Gigasporaceae)、丝盖伞科(Inocybaceae)和被孢霉科(Mortierellaoeae)等[46]。结果表明驯化使得部分有益微生物缺失,育种改良导致根际招募特定微生物的能力降低。此外,驯化还影响根际微生物群落装配模式和根际微生物之间的关系。驯化小麦根际微生物群落的形成依赖于随机组装过程,野生小麦根际微生物群落形成过程中生态位分化占主导地位[20]; 而且小麦种子内含有垂直传播的微生物,与栽培品种相比,野生品种种子内更多微生物类群与后续发育植株根际微生物组成相似[48]。与现代品种相比,地方品种的微生物网络更复杂,微生物间正负关联占比均衡,根际微生物群落更加平衡稳定[46]。
Bulgarelli 等[50]研究发现,在大麦根际土壤中,微生物群落β 多样性与宿主基因组之间存在显著的相关性,宿主基因型在一定程度上决定根际微生物群落特征,野生型根际拟杆菌门丰富,栽培型根际细菌优势类群为放线菌门和变形菌门。驯化大麦及其野生近缘种所面临的生态地理限制对根际微生物群的招募和维持具有一定影响。大麦“生态型”是根际微生物群落的决定因素,野生大麦调节根际微生物群落适应环境的能力更强[51]。通过宏基因组分析大麦根际微生物的潜在功能发现,与病原菌致病相关的基因(Ⅲ型分泌系统T3SS、毒力调节、入侵和细胞内抗性)以及与微生物分泌系统(Ⅵ型分泌系统;T6SS)、噬菌体-微生物相互作用(转座元件、噬菌体整合)和养分活化相关的基因在栽培型根际富集[50]。说明大麦的驯化影响其根际微生物的群落结构和功能。
玉米是重要的粮饲作物,也是高度驯化的C4 作物。在驯化过程中,玉米基因型对根际原核生物和真菌类群群落组成具有显著影响,研究发现玉米野生品种、自交系、杂交系根际3.7%的原核生物类群和4.9%的真菌类群与品种显著相关[52-54]; 同时驯化和生长发育时期对玉米根际细菌类群的相对丰度具有交互效应,自交系玉米根际厚壁菌门相对丰度高于地方品种和野生品种; 地方品种玉米苗期和开花期根际优势类群为链霉菌属(Streptomyces),而野生玉米品种在成熟期根际土壤检测到较高比例的链霉菌属[55]。虽然驯化影响玉米根际微生物组成,但研究发现其存在一个核心微生物组,主要包括: 农杆菌属(Agrobacterium)、德沃斯氏菌属(Devosia Nakagawa)、假单胞菌属(Pseudomonas)、梭状芽孢杆菌属(Clostridium)、拟杆菌属(Bacteroides)、肠杆菌属(Enterobacter),并且能够垂直传播(从一个世代传递到下一个世代)[56-57]。驯化对玉米根际微生物多样性的影响亦呈现出不同规律。Brisson 等[58]对玉米野生品种、自交系、杂交系的研究发现各个品种根际真菌α 多样性无显著差异; Matus-Acuña 等[59]发现自交系根际真菌群落多样性高于地方品种; Huang 等[55]对玉米根际细菌的研究也发现类似规律,自交系比地方品种根际细菌群落多样性高; 而Szoboszlay 等[11]发现野生玉米根际细菌多样性高于杂交系玉米,品种的差异对于玉米根际微生物多样性的影响更重要。此外,驯化和改良育种影响了根际微生物群落组装和功能,自交系根际微生物共现网络与野生品种相似,并且野生品种根际微生物网络结构较复杂[54]。玉米野生品种和地方品种的气生根粘液中存在固氮微生物,而现代杂交种中则不存在; Van Deynze 等[60]和Favela 等[61]研究了20 个玉米自交系(1949-1986年)发现,随着时间变化玉米对根际微生物供氮的依赖性降低,重氮营养体的丰度减少,表明驯化影响根际微生物的功能。
综上所述,禾本科主要C3、C4 作物根际微生物群落结构与功能均受宿主基因型影响。作物品种对根际微生物多样性影响较大,野生品种根际微生物多样性丰富。在胁迫环境下,栽培品种根际微生物群落通常易受干扰; 野生品种根际微生物群落更稳定,在一定程度上有助于提高其逆境适应能力。驯化使得植物自身生长策略发生改变,影响根际微生物功能,现代品种对根际供氮微生物的依赖性降低,野生品种根际富集更多的与营养获取有关的菌群。这些发现对研究驯化过程中谷子-微生物互作机制具有重要的指导意义; 通过研究驯化形成的不同植物基因型与微生物组之间的关系,探究植物与微生物潜在的互作机制,有助于深入理解根系相关微生物群落构建的生态过程,通过调节与植物基因型相关的微生物组成,可定向改善植物对环境的适应性和生产力,为育种策略提供信息,有助于作物生产与农业生态系统功能更好地结合。
2 驯化对谷子生长及生理特性的影响
谷子由青狗尾草驯化而来,据报道,大约5000~6000年前已在中国北方地区广泛种植,此后传播到世界其他地区[29,62]。据考古发现,史前时期的谷子遗迹在中国发现约60 处,黄河流域的谷子遗存分布最为密集,谷子和水稻的驯化种植共同构成了我国灿烂的农业文明[27-28,33]。
经过长期的人工选择和自然选择,现代谷子与青狗尾草相比,形态和生理方面发生了巨大的变化(图1): 谷子分枝分蘖性减少、植株更为健壮、落粒性和休眠性丧失、种子较大、产量更高、食用及饲用品质更佳[33,63-64]。青狗尾草基因组分型研究开展较早,其基因组在20世纪40年代就被命名为A[63]。由于谷子和青狗尾草同是二倍体且不存在生殖隔离,其基因组也被命名为A,但谷子损失了超过50%的遗传多样性,与水稻等作物的驯化过程极为相似[63]。谷子与其他禾本科之间存在密切的进化关系,其基因组中存在其他禾本科植物的保守基因,99%的保守基因与其他禾本科具有同源性[65]。将谷子的基因组图谱与短柄草(Brachypodium sylvaticum)、水稻、高粱和玉米进行对比研究发现,谷子约在27 000 万年之前从高粱和玉米中分化出来[64]。因此,谷子的相关研究对其他禾本科作物的探索具有一定的参考价值。随着遗传学和分子生物学的快速发展,谷子及其野生种的全基因组测序已经完成[63]。在此基础上,谷子驯化的遗传基础被逐步揭示,研究表明,驯化性状是由一个或少数几个主要基因所控制的。例如,Doust 等[66]发现谷子TB1 同源基因对谷子分枝分蘖性形成影响较小,仅与分枝伸长有关; 谷子有16 个遗传区域控制着光合作用响应和开花时间[67]。近期基于青狗尾草群体深度重测序的研究发现,谷子落粒性的丧失与青狗尾草中一个MYB 转录因子编码区转座子的插入有关[38]。在谷子功能基因组研究领域,谷子遗传转化及重要功能基因的克隆有了较大进步。Jia 等[63]对916 个谷子品种进行了测序,鉴定了58 万个单核苷酸多态性(single nucleotide polymorphism,SNP),并使用80 万个常见SNP 构建了谷子基因组的单倍型图谱,通过全基因组关联研究确定了与农艺性状相关的51 个基因座。Zhao 等[68]克隆了控制谷子叶片披垂的关键基因并开展了深入的分子遗传学研究。这些遗传学上的进步都将进一步促进谷子-根际微生物互作的研究。

图1 驯化过程中谷子遗传和农艺性状变化Fig.1 Changes of the genetic and agronomic traits of foxtail millet (Setaria italica) during domestication
谷子在我国栽培面积较广,据联合国粮农组织(FAO)统计,我国谷子种植面积为世界种植面积的80%,目前全国谷子年度播种面积在130 万hm2左右,总产量520 万t 左右,是潜在的解决粮食危机的重要杂粮作物。我国不同驯化阶段的谷子种质资源丰富,国家基因库已保存了近3 万份谷子种质资源,占世界保有资源的67%,主要来自北纬32°~48°和东经108°~110°,搜集了约500 份青狗尾草种质资源,地方种种质资源亦十分丰富。因此,选择不同驯化阶段的谷子进行研究,对驯化过程中谷子性状、基因、微生物组及互作等进行挖掘,揭示谷子根际微生物响应驯化的机制,将为其他禾谷类作物根际微生物的研究提供科学依据[26,69]。
3 驯化对谷子根际微生物的影响
驯化对谷子根际微生物群落多样性及组成的影响存在差异。Chaluvadi 和Bennetzen[70]对4 种狗尾草和6 种栽培谷子根际相关的细菌和古菌群落组成及多样性进行了对比研究,发现狗尾草和栽培谷子品种间根际细菌多样性无显著差异,而栽培谷子根际古菌物种丰富度和均匀度高于狗尾草根际古菌群落; 且狗尾草和栽培谷子根际细菌和古菌群落组成存在较大差异,与栽培谷子相比,狗尾草富集更多与固氮作用相关的细菌类群。对狗尾草和金色狗尾草(Setaria pumila)的根和穗中微生物群落组成研究发现,与穗相比,根部微生物类群丰富,其中,根瘤菌属(Rhizobium)为优势类群,两个狗尾草根相关细菌多样性存在差异,狗尾草根部细菌多样性高; 根和穗相关微生物区系受植物部位和采样地点的影响显著,根部微生物群落受宿主基因型的影响较大[71]。在不同品种谷子各生长发育阶段,由于谷子基因型不同,造成根际微生物环境存在差异,真菌种群随着谷子的生长发育由复杂趋于简单,在谷子生长发育之始真菌多样性高于细菌; 而细菌的多样性和群落组成则越来越复杂,数量也越来越丰富[72]。在前人的研究中也发现,作物驯化对真菌群落的塑造效应大于细菌群落,且真菌群落比细菌群落与宿主植物的关系通常更为密切。如菌根真菌能够促进谷子对氮磷等养分的吸收,提高其生长及抗性能力。然而,作物驯化造成细菌和真菌群落差异的机制及驱动因素研究少见报道,仍有待深入探索。
驯化影响作物根际微生物功能[45-46,73],不同品种谷子根际微生物的功能存在差异。Jin 等[72]通过对2882 例谷子的根表、根际和空白土壤进行微生物测序研究,并与谷子的重要产量性状等进行关联分析,鉴定出了与谷子高产相关的潜在有益菌,并发现这些益生菌间存在显著的互作关系。此外,研究人员通过微生物功能预测发现,栽培品种的根表面富集了与养分吸收、环境响应相关的微生物群,而根际土壤则富集了与碳固定和氨基酸合成相关的微生物群[72]。该研究为进一步筛选并建立潜在益生促生菌资源库提供了指导。Niu 等[74]从辽宁建平的谷子根际土壤中分离得到了6 个不同属[假单胞菌、肠杆菌、泛菌(Pantoea)、节杆菌(Arthrobacter)、克雷伯氏菌(Klebsiella)和赭曲霉(Aspergillus)]的细菌菌株,在干旱胁迫下,这些细菌对谷子种子的萌发和幼苗生长具有促进作用。谷子根际微生物群落中特定的菌群在一定程度上增强了其自身抗性。Han 等[75]将克氏黑穗病菌(Ustilago crameri)接种到3 个黑穗病抗性品种后,发现发病率低的‘吉谷20’根际微生物多样性更高,与‘金谷21’和‘昌农35’相比,其放线菌门丰度较高,拟杆菌门和厚壁菌门丰度较低; 慢生根瘤菌(Bradyrhizobium)和链霉菌(Streptomyces)的丰度与黑穗病发病率呈负相关。然而,植物与微生物的相互作用并不是随机的,且受到植物基因型的影响。研究人员基于827 份不同品种谷子的基因组遗传变异、根表微生物以及谷子生长和产量等数据,应用多组学(全基因组、宏基因组和微生物组全基因组)联合分析,揭示了谷子基因型、根系微生物组与农艺性状之间的互作关系,提出了基因型依赖的微生物效应。为深入理解根系微生物群落构建的生态过程,通过调节与植物基因型相关的微生物组成,定向改善植物对环境的适应性和生产力提供了新途径[76]。然而,作物根际微生物群落组成除与宿主基因型相关外,还受微环境及气候变化等因素的影响[77-79]。驯化导致谷子-微生物互作、微生物间互作变化的机制仍存在许多关键问题有待厘清。如驯化对谷子根际微生物群落塑造的驱动因素有哪些? 是如何影响微生物群落的组装过程的? 在气候变化背景下,要恢复和强化谷子与有益微生物的相互作用,需加强驯化对谷子微生物群落塑造的过程和机理的研究。
4 结论与展望
人类的驯化和选育过程导致现代作物品种与其野生祖先品种在生理生长特性、遗传多样性等方面存在显著差异,同时,与野生品种相比,现代作物品种根际微生物多样性降低、有益微生物丧失、根际微生物互作网络稳定性降低(图2)。随着作物微生物组的深入研究,人们逐渐意识到驯化过程对微生物产生的影响不可忽视,作物和微生物之间存在共进化关系,探寻并利用驯化过程中丢失的有益微生物,或可弥补栽培种在抗逆方面的缺陷[2]。然而,目前对于驯化过程中作物-根际微生物相互关系的研究仍处于起步或描述性阶段,且多集中于豆类、水稻等几种作物中。伴随着遗传多样性的丧失,作物根际微生物多样性降低; 根际微生物群落对环境更敏感,有益微生物群落失去优势或丢失,但其内在机制仍不明确。对于作物驯化梯度对根际微生物效应的对比分析不全面,并且作物不同生长时期也影响根际微生物,但不同驯化阶段作物各生长时期根际微生物的对比几乎空白。另外,在环境胁迫条件下,不同驯化阶段作物根际微生物的相关研究亦是缺乏。因此,丰富研究物种类群,挖掘不同驯化阶段作物-根际微生物互作机制,并摸索应用作物-微生物互作机制及潜在微生物类群在现代育种中的应用等仍是亟需推进的研究方向[76]。
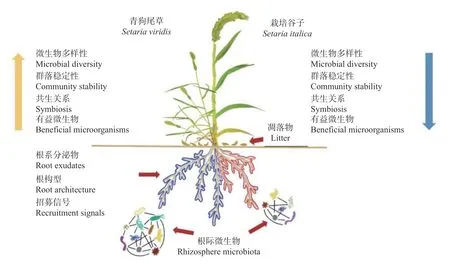
图2 谷子驯化对其根际微生物影响的模式图Fig.2 Schematic illustration of the impact of foxtail millet (Setaria italica) domestication on the rhizosphere microbes
近年来谷子及其野生品种作为C4 和抗旱耐逆研究的模式植物越来越受到关注。谷子不仅是抗旱作物,因其耐瘠薄、适应性广、稳产性强,化肥农药用量少等特性,也是典型的环境友好型作物[26,31-32]。野生品种抗逆性更强,结合现有多组学研究手段,有效利用分子生物学方法,探究谷子野生型和栽培型品种的表型与根际微生物互作机制,利用野生品种根际微生物生态功能,可以在低投入或基于生物的系统中提高谷子及其他禾本科生产力。谷子和青狗尾草的基因组小且具有株型矮小、自花授粉、生育周期短、繁殖系数高等优势,适合利用遗传转化和基因编辑等分子手段对其进行分析和实验栽培[26]。在遗传学上,谷子与高粱、玉米、水稻等的序列一致性超过70%[65]。谷子的这些特性使得其成为基因组学、遗传学、发育与进化研究的新兴C4 模式植物,应用谷子及其野生种进行驯化-作物-微生物互作机制的研究,对推进其他禾本科植物的相关研究具有参考意义[26,35-38]。
在探索不同驯化阶段谷子-微生物互作机制过程中,将微生物组与作物驯化遗传调控网络系统结合,也是研究作物驯化过程对微生物影响机制的一个关键突破口。由于根系分泌物以及养分的传输是作物和微生物相互选择的重要媒介,相对于基因组学和转录组学来说,代谢组学的分析和研究能更有效地解释作物驯化带来的根际微生物组的变化规律[76-77]。将生物和非生物胁迫引入到驯化-作物-微生物互作这个系统中也是研究的一个必要方向[80]。更好地了解不同驯化阶段作物根际微生物群落结构和功能,阐明微生物性状丧失或获得的遗传基础,将提高我们预测驯化过程中植物相关微生物群落的结构和对植物健康影响的能力,以保证农作物的健康和生产力[81-82]。
探究作物与微生物组之间的互作效应,有助于深入理解根系微生物群落构建的生态过程,通过调节与植物基因型相关的微生物组成,可定向改善植物的环境适应性和生产力[76,83]。目前,为恢复农田系统作物-微生物的有益关系,基于对多种作物的研究从育种角度提出了一个可行的操作步骤。首先从作物驯化起源地获取土壤,从种质资源库中选取品种,通过温室栽培试验比较现代种、当地种、野生种在原生土壤与农田土壤的微生物组成与功能,然后分离培养有益微生物,最后寻找微生物群落组成和功能与作物遗传之间的相关性,通过分子育种和标记筛选选择能建立作物-微生物有益关系的品种,重建作物与有益微生物之间的关系[2]。虽然这一流程的实际应用仍在不断尝试探索,但也为谷子育种提供了一个全新的方向。在未来的农业系统中,利用作物-有益微生物互作关系,通过精准的微生物组管理设计高产的栽培品种,将更加高效地促进农作物产量的提升,提高农业绿色可持续性发展。
猜你喜欢
当代陕西(2021年21期)2022-01-19
中国土壤与肥料(2021年5期)2021-12-02
作文周刊·小学一年级版(2020年40期)2020-10-19
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
小学生作文选刊·低年级版(2016年6期)2016-06-24
现代农业(2015年5期)2015-02-28
中国火炬(2012年9期)2012-07-25
植物营养与肥料学报(2010年6期)2010-10-26
