
草地贪夜蛾幼虫感受器扫描电镜观察
2023-04-06 06:43伏倩文吴海朝胡瑞瑞李星月祖国浩白鹏华
天津农业科学 2023年3期
伏倩文,吴海朝,胡瑞瑞,李星月,祖国浩,白鹏华
(1.天津农学院园艺园林学院,天津 300384;2.天津市农业科学院,天津 300384;3.四川省农科学院植物保护研究所,四川成都 610066)
感受器由昆虫体壁的皮细胞以及感觉细胞构成,是昆虫感知外界刺激的基本单位,可以对外界刺激和内部刺激产生反应,负责嗅觉的检测和触觉信号的感知[1]。气味分子通过扩散进入昆虫的感受器中,在嗅觉蛋白作用下运输至嗅觉受体,将外界化学信号转化为电信号,并最终传递到更高级的中枢神经系统,引起昆虫对外界刺激作出行为反应[2]。化学气味经感受器介导而指导昆虫产生不同的行为,如求偶、交配、取食、寄主选择和躲避天敌等重要生命活动[3-4]。感受器的类型、分布、数量等会因为昆虫种类、地理种群和性别的不同存在显著差异,进而影响昆虫的取食、交配和产卵等行为特异性。美国白蛾Hyphantria cunea 雄性触角上的毛形感受器数量明显多于雌性成虫的数量,且长度更长[5];稻纵卷叶螟Chaphalocrocis medina 雄成虫触角上锥形感器数量显著多于雌虫的数量[6];与雄虫触角相比,桃小食心虫Carposina niponensis Walsingham 雌虫触角上感受器的种类更加丰富,推测其可能与雌虫产卵密切相关[7];马涛等[8]推测香梨优斑螟Euzophera pyriella Yang 雄虫触角上特有的柱形感受器可能参与性信息素的识别。因此,研究昆虫感受器的结构、分布对害虫取食、交配和产卵等行为具有重要指导意义。
草地贪夜蛾Spodoptera frugiperda 隶属于鳞翅目Lepidoptera,夜蛾科Noctuidae,灰翅夜蛾属Spodoptera Herrich-Schaeffer,是世界农业重大迁飞性害虫,并被列入世界十大植物害虫“黑名单”[9]。草地贪夜蛾迁飞能力极强,自2019 年入侵我国云南省,现已扩散至全国27 个省(区、市),年均发生面积近120 万hm2,每年对我国玉米造成的潜在经济损失可达54 亿~470 亿美元[10-11]。该昆虫食性杂,尤其嗜好禾本科作物,分为玉米型和水稻型2 个品系。经鉴定入侵中国的地理种群为玉米型,玉米田草地贪夜蛾发生面积占该虫总发生面积的98%以上[11]。草地贪夜蛾的快速入侵和严重危害,对我国的农业生产造成了巨大的经济损失和严重的威胁。报道表明,草地贪夜蛾完成一个世代要经历卵、幼虫、蛹和成虫,完成整个世代平均为45 d[12]。其中幼虫期共分为6 个龄期,1~2 龄幼虫喜好聚集取食,但进入3 龄幼虫不再聚集取食且进入暴食阶段,因此在3 龄期前对其进行防治极为重要[12]。基于此,本研究选择聚集取食阶段的低龄幼虫期2 龄为研究对象,对草地贪夜蛾2 龄幼虫的头部、胸足,以及腹足进行电镜扫描,对感受器的类别、数量、分布等分别进行了详细的观察,为探索草地贪夜蛾2 龄幼虫化学感受器行使的聚集取食、寄主识别和躲避天敌等重要行为功能提供理论依据,并为进一步采用化学生态学方法防控草地贪夜蛾低龄幼虫提供理论支持。
1 材料与方法
1.1 供试虫源
草地贪夜蛾幼虫购自于河南省科云生物农药有限公司,用人工饲料在智能气候箱(RXZ-280B,宁波东南仪器有限公司)进行饲养并繁殖多代,饲养条件设置为:温度24~27℃,相对湿度60%~75%,光照周期L∶D 为16∶8。采用最新孵化的2 龄幼虫进行扫描电镜试验。
1.2 样品制备
取草地贪夜蛾2 龄幼虫用于样品制备,首先用2.5%戊二醛将样品固定24 h;其次用磷酸缓冲液洗涤3 次;最后用不同浓度(75%、85%、95%、100%)的乙醇进行逐级脱水,室温自然风干。将脱水的样本在临界干燥器上干燥后,置于电镜专用载物台上,经溅射仪喷金后进行拍照记录[5]。
1.3 数据统计与分析
在电镜下扫描拍照后,利用软件自带的测量与标尺功能进行数据的标示,数据以平均值±标准误(Mean±SE)表示。每组测量数据至少观察5 头2 龄幼虫体躯的各项数值,观察到的感器根据Schneider(1964)和Zacharuk(1980)的文献进行分类[13-14]。
2 结果与分析
2.1 头部
草地贪夜蛾2 龄幼虫的头部黑色,呈现椭圆形(图1-A)。头部表面具有刚毛,头式为下口式,口器为咀嚼式口器。头部可见额、额缝、头冠缝(图1-B),和1 对触角、刚毛、口器、吐丝器、单眼(图1-C)。单眼两侧各6 个(O1~O6),呈“C”字状排列(图1-D)。
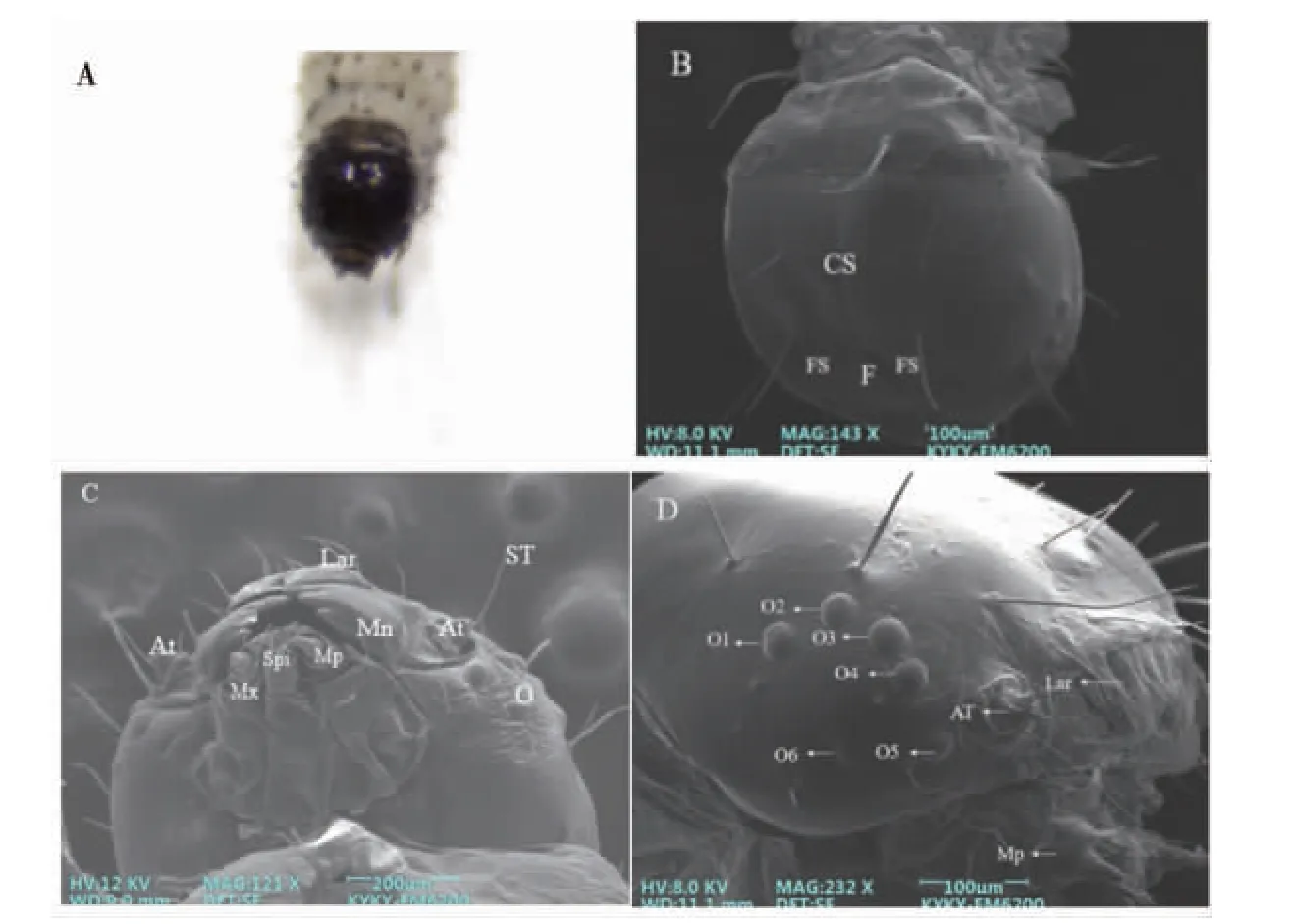
图1 草地贪夜蛾2 龄幼虫头部扫描电镜图
2.1.1 触角 草地贪夜蛾2 龄幼虫的触角形状粗短,位于头部的侧下方、单眼和上颚之间(图2-A)。触角长约(53.1±1.48)μm,由柄节、梗节和鞭节3 部分构成,柄节长约(7.4±0.2)μm、宽约(68.4±1.38)μm,位于触角窝内;梗节为圆柱形,嵌于柄节端部凹陷的窝内,长约(22.8±0.378)μm、宽约(38.4±0.34)μm;鞭节呈现短锥状,长宽分别约为(22.9±0.33)μm 和(16.6±0.19)μm。触角柄节处未观察到感受器(图2-A);梗节上着生栓锥形感受器、毛形感受器、刺形感受器各1 个(图2-B),长度分别约为(10.2±0.46)μm、(112±1.91)μm、(4.9±0.21)μm;鞭节上着生1 个栓锥形感受器,长宽分别约为(9.9±0.48)μm、(4.79±0.15)μm,2 个长约(2.61±0.16)μm 的Ⅰ型锥形感受器和2 个长约(12.2±0.2)μm 的Ⅱ型锥形感受器,其中Ⅰ型锥体短小,而Ⅱ型锥体较大(图2-B)。

图2 草地贪夜蛾2 龄幼虫触角及上唇扫描电镜图
2.1.2 上唇 上唇着生大量刺型感受器,对称分布约6~7 对刺型感受器(图2-C)。
2.1.3 上颚 上颚位于上唇的下方,与上唇紧挨。上颚前端有锋利的切齿,用来切断食物;后部具有一个粗糙面,用以磨碎食物,因此上颚的硬度较高并且十分发达(图3-A)。
2.1.4 下颚及下颚须 下颚拥有协助草地贪夜蛾幼虫刮、切、握、持食物等功能,对上颚起到辅助作用。下颚分为轴节和茎节,在两节交界处着生2 个长约(15.6±0.19)μm 的毛形感受器(图3-B);端节着生2 个长约(8.51±0.16)μm、宽约(5.8±0.15)μm 的栓锥形感受器和4 个刺形感受器(图3-B);下颚须共分为三节,其远端体节上分布8 个小锥形感受器(图3-C)。
2.1.5 下唇 下唇由前颏和后颏两部分组成,下唇须位于前颏前侧的两端,上面各有1 个刺形感受器和1 个栓锥形感受器,分别长约(14.6±0.52)μm、(11.7±0.36)μm(图3-D)。下唇上着生有1 个吐丝器,吐丝器的下方着生1 对长约(3.54±0.12)μm 的刺形感受器(图3-D)。
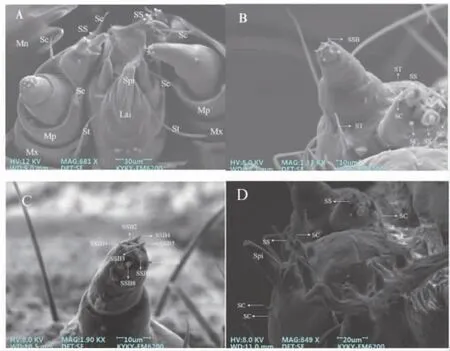
图3 草地贪夜蛾2 龄幼虫口器扫描电镜图
2.2 体躯
草地贪夜蛾2 龄幼虫体长约(7.14±0.13)mm,宽约(1.06±0.04)mm。正面观与背面观体节分节明显,可见12 个体节,其中胸部3 节,腹部9 节(图4-A,B)。幼虫背部着生大量刚毛,腹部感受器相比背部较多,胸节腹面具有大量突起。
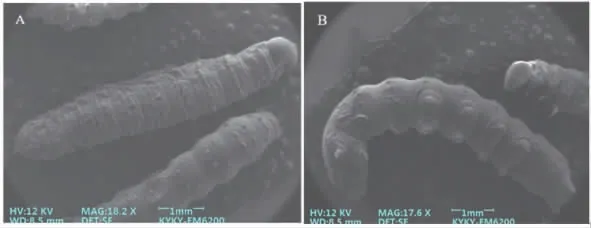
图4 草地贪夜蛾2 龄幼虫正面观与腹面观
2.2.1 胸足 草地贪夜蛾2 龄幼虫共有3 对胸足,每一胸节上各有1 对胸足(图5-A)。胸足顶端具有2~5 个爪,呈现弯曲状;胸足周围和胸节上具有数根刺形感受器与毛形感受器(图5-B)。第1 胸节两侧具有纵向排列的3 个气孔(图5-C、图5-D)。
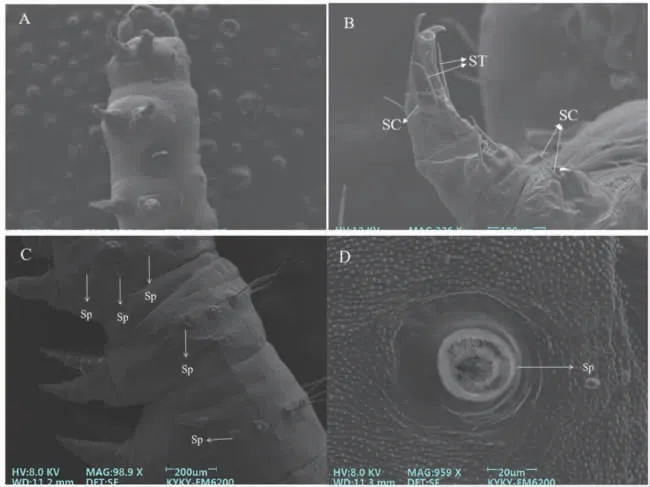
图5 草地贪夜蛾2 龄幼虫胸足及其气孔
2.2.2 腹足 草地贪夜蛾2 龄幼虫具有9 节腹节,其中第1 腹节到第6 腹节的两侧各有一对气孔。5对腹足分别着生在第3~6 腹节以及第9 腹节上,其中末节上的足又称为尾足、臀足。在5 对腹足末端存在数量不一的趾钩,数量为8~10 个(图5-C)。与胸足相同,腹足上分布大量的刺形感受器、毛形感受器和少数锥形感受器。
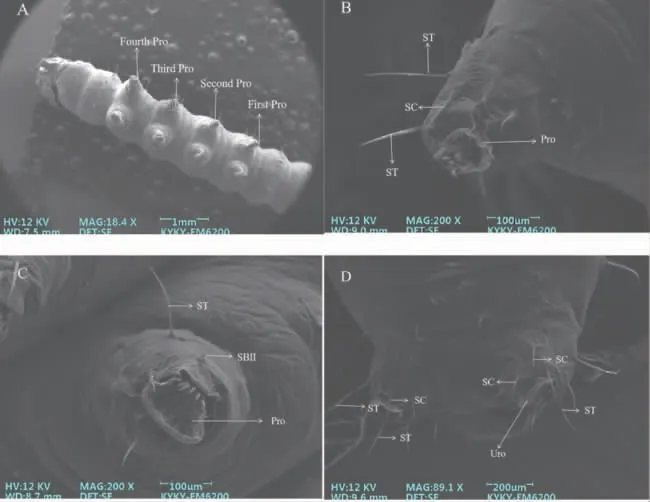
图6 草地贪夜蛾2 龄幼虫腹足及其上感受器
3 讨论和结论
草地贪夜蛾具有寄主范围广、繁殖能力强、迁飞扩散快、暴饮暴食等特点,给防治带来巨大困难与挑战。草地贪夜蛾幼虫的口器为咀嚼式口器,2 龄幼虫在小喇叭口期和大喇叭口期主要取食叶片,在抽雄吐丝期取食花丝,严重时会危害雌穗籽粒。因此,全面了解2 龄幼虫头部、胸足和腹足的感器类型和分布,有助于从形态结构方面进行鳞翅目幼虫的分类研究,同时揭示幼虫取食寄主植物的化学生态机制。
本研究采用扫描电镜观察了草地贪夜蛾2 龄幼虫周身感受器的种类、数量和分布。结果表明,幼虫触角上着生的感受器有锥形感受器、刺形感受器、毛形感受器,以及栓锥形感受器,与5 龄幼虫和鳞翅目昆虫大菜粉蝶Pieris brassicae 和云南锦斑蛾Achelura yunnanensis 幼虫触角感受器类型完全一致[15-17]。尽管草地贪夜蛾2 龄与5 龄幼虫触角上感受器类型一致,但数量存在一定的差异,因此,昆虫触角感器随着昆虫发育过程的改变可能与外界环境相适应有关[18]。研究表明,毛形感受器和锥形感受器均可以感受植物挥发物,进而定位识别寄主植物完成取食和产卵等重要行为活动[19-20]。如东亚飞蝗Locusta migratoria manilensis 负责识别寄主植物的气味受体基因LmigOR2 和LmigOR12 均在触角上锥形感受器神经元中高度表达[21],因此推测幼虫触角上锥形感器可能参与了寄主植物识别。此外,刺形感受器在桃小食心虫C.Walsingham 和朱红毛斑蛾Phauda flammansWalker 中推断具有感知触觉和味觉刺激的作用[7,22]。与幼虫触角毛形感受器不同的是,鳞翅目成虫触角上的毛形感受器不仅可以用来感知寄主植物,对于求偶交配、繁衍下一代也极其重要。例如,鳞翅目昆虫多音天蚕Antheraea polyphemus、甜菜夜蛾Spodoptera exigua和烟青虫Helicoverpa assulta等成虫触角上的毛形感受器中均有高度表达性信息素受体的嗅觉基因[23-25]。综上所述,草地贪夜蛾幼虫触角拥有感知味觉、嗅觉、与温湿度的功能,如感知植物气味进而识别寄主植物。
与触角感器类型一致,草地贪夜蛾2 龄幼虫口器也分布有锥形感受器、刺形感受器、毛形感受器和栓锥形感受器,与其他鳞翅目幼虫口器感受器类型相似[20,26]。草地贪夜蛾2 龄幼虫口器上感受器类型和数量均多于5 龄幼虫,有研究表明昆虫特定部位的触角感器类型伴随发育过程中会消失[18]。刺形感受器是草地贪夜蛾幼虫口器上数量最多的类型,根据该感受器功能推断其与昆虫取食时握持食物有关[27];其他感受器可能具有定位寄主植物作用[26,28-29]。
草地贪夜蛾2 龄幼虫胸足、腹足的形态存在明显差异,且感受器数量不一。例如,胸足端部具有2~5 个数量不一的弯曲型的爪,有报道表明其与鳞翅目幼虫钻蛀、取食、爬行有关[26,30];腹足末端存在数量不一的趾钩,而趾钩的长短、数目和排列方式是鳞翅目幼虫分类鉴定的主要特征之一[31]。此外,胸足和腹足上存在大量的毛形感受器和刺形感受器。刺形感受器除了机械感受外,在爬行过程中对体躯具有一定的支撑和保护作用[32]。
近年来,关于昆虫感受器感知外界环境的研究备受关注,通过扫描电子显微镜对鳞翅目、半翅目和膜翅目等昆虫的触角感器类型和分布开展大量研究。多数感受器研究集中在成虫触角,而作为取食、危害的关键虫态幼虫的报道相对较少。本研究全面观察了草地贪夜蛾2 龄幼虫周身的感受器类型、数量和分布,为鳞翅目幼虫分类和幼虫感知寄主植物等功能奠定了理论基础。
猜你喜欢
昆明医科大学学报(2021年1期)2021-02-07
云南农业大学学报(自然科学)(2021年1期)2021-01-22
华人时刊(2020年19期)2021-01-14
中国现代医药杂志(2020年10期)2020-12-14
文学少年(有声彩绘)(2017年1期)2017-02-14
中华老年口腔医学杂志(2016年5期)2016-03-01
中国继续医学教育(2015年5期)2016-01-07
天津大学学报(自然科学与工程技术版)(2015年10期)2015-12-29
中国火炬(2013年6期)2013-07-24
环境昆虫学报(2013年4期)2013-06-11
