
丛枝菌根真菌修复重金属污染土壤及增强植物耐性研究进展
2023-03-24 08:52:16苗志加孟祥源李书缘马超李晴安贺銮赵鑫赵志瑞
农业环境科学学报 2023年2期
苗志加,孟祥源,李书缘,马超,李晴,安贺銮,赵鑫,赵志瑞,2,3*
(1.河北地质大学水资源与环境学院,石家庄 050031;2.河北省高校生态环境地质应用技术研发中心,石家庄 050031;3.河北省水资源可持续利用与产业结构协同创新中心,石家庄 050031;4.河北地质大学自然资源资产资本研究中心,石家庄 050031)
土壤重金属污染是关乎人类健康和粮食安全的重大问题。采矿、冶炼、电子工业、化石燃料燃烧、农业生产等人类活动所产生的重金属是土壤重金属的主要来源[1-2]。重金属污染具有隐蔽性、难降解、易积累等特点,土壤中过量的重金属可以通过生物链在人体内富集,对人体产生“三致”毒害作用[3-4],因此土壤重金属污染修复是目前亟需开展的工作。目前,常用的土壤重金属修复方法有物理法、化学法、生物法[5]。植物修复作为生物法中的一种,其与物理化学方法相比,具有成本低、对土壤没有二次污染及损伤、简单易行等优点[6],同时也存在植株矮小、生长缓慢、修复效率低、修复时间长等缺点[7],但是微生物和植物的联合作用可以弥补单一修复技术的缺陷。
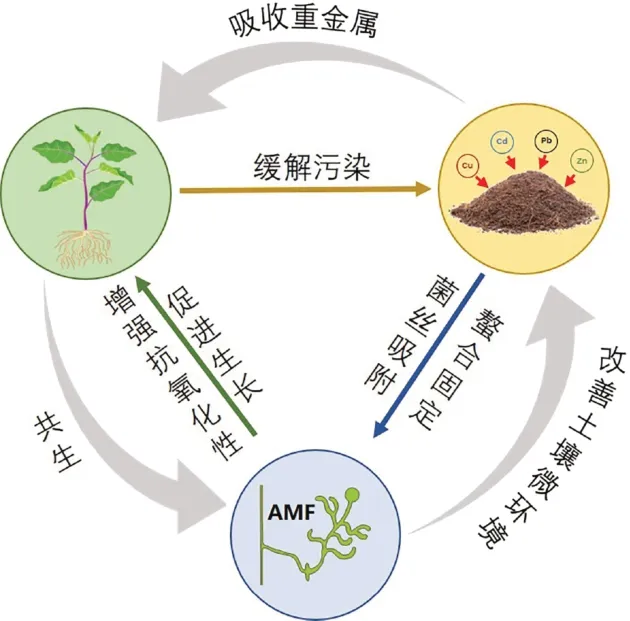
丛 枝 菌 根 真 菌(Arbuscular mycorrhizal fungi,AMF)是一种能与大多数陆地植物建立互惠共生关系的土壤有益菌[8],可以帮助宿主植物扩大根系,促进植物吸收营养元素及矿物质,刺激菌根分泌抗氧化物质抵御重金属污染胁迫,从而达到提高植物修复效果的目的[9]。目前,关于接种AMF 对重金属胁迫下植物生长发育、重金属吸收、分布规律及富集特征方面的影响研究较多,但是关于AMF增强植物修复重金属污染的机理研究并不深入。基于此,本文概述了AMF对土壤重金属修复的作用效果,并分别从植物、土壤及真菌自身3个角度阐述了AMF对重金属的解毒机制。
1 AMF概述
1885年,Frank定义了“菌根”这一名词[10],它是一种高等植物和真菌的特殊共生体,与植物共生的真菌为菌根真菌,其菌丝可延伸到根际周围和土壤中[11]。按照真菌在植物根系的寄生位置可以将菌根分为内生菌根(Endomycorrhiza)、外生菌根(Ectomycorrhiza)以及内外生菌根(Ectoendomycorrhiza)[12]。AMF 是一种内生菌根真菌,其与植物形成的菌根主要包括菌丝体、泡囊和孢子等结构,因在宿主植物根皮层细胞内的“丛枝”结构而得名[13]。目前研究和应用较多的AMF 有根孢囊霉属(Rhizophagus)中的异形根孢囊霉(Rhizophagus irregularis)、根内根孢囊霉(Rhizophagusintraradices),球囊霉属中的摩西球囊霉菌(Glomus mosseae),斗管囊霉属(Funneliformis)中的摩西斗管囊霉(Funneliformis mosseae)、地斗管囊霉(Funneliformis geosporum),多样孢囊霉属(Diversispora)中的地表多样孢囊霉(Diversispora epigaea)[14-18]等。AMF 广泛分布于各陆地生态系统中,其能与80%~90%的陆生植物共生形成菌根结构,这对于维持植物的多样性和生态系统的稳定性有着重要意义[19]。目前研究表明,AMF 在促进植物生长、增强植物在逆境胁迫下耐性、提高植物修复重金属及有机物污染能力方面都有十分重要的作用。
在植物-真菌-土壤体系中,AMF 菌丝的存在增大了植物根系与土壤的接触面积,从而可以帮助植物根系吸收水分以及营养物质,促进植物生长。很多研究表明,接种AMF 可以活化根系土壤中氮磷元素,从而提高玉米(Zea mays)、小麦(Triticum aestivum)、大豆(Glycine max)等主要粮食作物的产量[20-21],增加苜蓿(Medicago sativa)、白三叶草(Trifolium repens)、黑麦草(Lolium perenne)等草本植物植株高度、生物量以及根系长度[22-23]。AMF 促进植物生长的同时能帮助植物抵抗生物和非生物胁迫,提高植物对不良环境的接受程度。生物胁迫包括害虫、杂草及病原菌,其中真菌导致的病害占所有统计的85%以上[24]。目前已被报道的具有抗病虫害能力的AMF 有摩西管柄囊霉、地斗管囊霉、摩西球囊霉、地表球囊霉、缩球囊霉、幼套球囊霉、根内根孢囊霉等,其中90%以上集中在球囊霉科的球囊霉属和斗管囊霉属[24]。非生物胁迫主要有干旱、盐碱、重金属等。干旱环境下,AMF 可以显著提高植物的生长指标和光合参数[25],菌丝可直接吸收部分水分改善植物水分状况[26],还可诱导相关基因表达间接提高植物的抗旱性[27]。徐瑶等[28]发现在NaCl 胁迫下接种摩西球囊霉能减缓红花的盐胁迫,主要通过以下途径:提高红花渗透调节物质的含量、增强矿质元素的吸收、减少红花对Na+的吸收、减少盐对叶绿素合成的干扰及减少膜脂过氧化的程度等。面对重金属的毒害,Deram 等[29]研究了Cd、Pb 和Zn复合污染土壤中AMF的定殖情况,发现菌根侵染几乎完全不受土壤污染的影响,这说明AMF可以抵抗重金属的胁迫。将AMF 与重金属修复相关联最早是在2002年,Jamal等[30]首次发现在Zn 和Ni污染土壤中接种AMF可以提高大豆和小扁豆对重金属的吸收效率,从而引发了科研人员对于AMF 缓解土壤重金属污染方面的关注。
2 AMF对重金属污染土壤的修复
AMF 缓解土壤重金属污染的方式主要是通过影响植物对重金属的吸收,但AMF 种类繁多,其功能呈现多样化,因此对于不同宿主植物以及不同重金属的作用效果也不一致。表1 总结了近年来关于AMF 对重金属修复的相关研究及修复效果。一般来说,接种AMF 可促进重金属固定在植物根系部分,减少其向地上部分的迁移。在Cd 浓度为400 mg·L-1条件下分别接种5 种AMF,各处理均明显促进了芹菜对Cd 的吸收,根部Cd 含量、Cd 滞留率及富集系数显著提升(P<0.05),茎叶部Cd 含量、迁移率显著降低(P<0.05)[9]。Huang等[39]在4种Pb水平下(0、90、900、3 000 mg·kg-1)对刺槐接种摩西管柄囊霉进行对比实验,结果表明接种菌剂植株的根和茎的Pb积累和转运能力更强,但叶片中的Pb 积累和转运能力更低。然而并不是所有的AMF 都可以将重金属螯合固定在根部。如沾屑多样孢囊霉可以增加紫花苜蓿地上部分Cd、Zn 两种重金属的累积量,减少根部Zn 含量[40],这表明部分AMF 也可能促进重金属向地上部分的转移。此外也有研究表明,AMF 在植物根部定殖对植物吸收重金属几乎没有影响[41]。Citterio 等[42]在100 µg·g-1Cd 和Ni 及300µg·g-1Cr 污染的土壤中接种摩西球囊霉菌,发现与不接种菌剂的植物相比,两者植物器官中的Cd 和Cr 含量没有统计学差异,并认为植物体内吸收的重金属的分布与土壤重金属含量有关。接种AMF后不同植物对重金属胁迫产生的响应差异较大,甚至同种AMF 对于不同品种的同类植物的重金属累积效应也有差距。Chen等[43]观察到,外来AMF能促进雌性植物根系对Pb的吸收和积累,但对雄性植物根系没有促进作用。宿主植物类型、对菌根依赖强度以及基因型的差异会对AMF 的作用效果产生影响[44]。AMF对植物吸收和转运重金属的效应可以是促进、抑制或不产生影响,但目前AMF与哪些特定的植物联合表现出促进或是抑制作用,尚无一致的结论。

表1 AMF修复重金属污染的作用效果Table 1 Effects of AMF on remediation of heavy metal pollution
AMF 的作用效果还与重金属含量、重金属种类、菌根真菌种类、土壤理化特性、金属之间的竞争、植物-微生物关联类型、植物生长条件和根系密度[45]、植物对污染物的耐受性和重金属的生物有效性[46]等诸多因素有关。目前人们关注较多的因素为重金属含量与土壤理化特性的影响。随着土壤重金属污染程度的增加,可能存在一个临界含量,低于该含量时,AMF 促进植物对重金属的吸收,而高于该水平时,重金属向寄主植物地上部分的转移受到抑制[47]。如Zn元素在植物生长过程中发挥着重要作用,是植物体必需的营养元素,但当含量过高时,其也是一种潜在的有毒金属。Chen 等[48]在石灰性土壤中添加不同比例的Zn(ZnSO4溶液)来研究AMF 在红三叶草Zn 吸收中的作用,结果均显示对应临界施Zn 量为50 mg·kg-1。这一结果并不是对所有土壤及植物都适用,对于不同土壤类型和植物物种的临界值还需要更准确地确定。一般来说,沙土、轻壤土中孢子密度较大,黏土中较小[49];有机质、速效氮、速效钾含量较高的土壤AMF丰度及侵染率较大,而pH较大、速效磷含量较高的土壤AMF存活会受到抑制[50]。因此土壤的理化性质会影响菌根侵染程度,从而间接影响AMF对土壤重金属的修复效果。AMF耐重金属胁迫的规律及上述影响因子发挥的作用机理目前尚不明确,仍需进一步研究。
目前,AMF 增强植物修复土壤重金属污染能力这一技术尚未得到广泛应用,其限制因素除AMF 作用效果不一、对土壤及植物的限制条件较高外,还与AMF 很难进行离体纯培养有关。AMF 菌剂扩繁常用的方法有盆栽培养、无土培养、大田培养、离体纯培养和离体双重培养[51]。其中盆栽培养成本低廉、操作简单,是目前应用最为广泛的菌剂培养方法,但是其缺点是培养周期长、培养效率较低。无土培养对配置要求较高,需要专业设备和技术人员,成本较为昂贵,并不能作为主要的菌剂生产手段。大田培养成本最低,可快速获得大量菌剂,但得到的真菌孢子不易收集且不够纯净。AMF 的离体纯培养方法还处于探索阶段,目前没有取得突破性进展,因此限制了其在各领域的深入研究及其在生产中的大规模应用[52-53]。我国也已经分离获得了多个高效菌株,但由于市场需求有限、技术体系不成熟及缺乏投入,尚未实现实验菌剂的大规模工厂化生产和广泛的商品化应用[54]。
3 AMF增强植物重金属耐性机制
AMF 促进植物吸收重金属,提高其对重金属耐受性,减小重金属毒害作用的机制十分复杂。在共生系统中,土壤、植物以及真菌自身对于重金属胁迫都会发生系列变化以进行防御,AMF 可以作为植物和土壤之间的桥梁,在吸收、转运、固定重金属过程中发挥重要作用。一方面,AMF 可以通过改善植物的生长状况、调节转运蛋白抑制重金属吸收、增强植物抗氧化能力等途径提高植物对重金属胁迫的抗性;另一方面,AMF 可以通过菌丝吸附固持重金属、分泌有机酸螯合重金属、改变重金属有效态含量等途径减缓重金属对植物的毒害作用。以下分别从生长稀释及限制吸收作用、抗氧化机制、螯合重金属机制、吸附固持重金属机制4个方面来详细阐述。
3.1 生长稀释及限制吸收作用
许多研究报道,接种AMF 能够增加植物根系对微量元素及矿物质等营养元素的吸收,帮助植物吸收水分,使植物各生长指标得到改善[55-57],如图1 中的①、②过程。这种生长刺激与以下事实有关:AMF 将吸收网络延伸到根际的养分消耗区之外,从而可以接触到更大体积的土壤[58-59]。此外,真菌菌丝远比根细,因此能够穿透较小的气孔并吸收更多的营养[60]。在Cd胁迫下,接种AMF能显著提高Cd污染土壤中植物新梢的氮磷含量[61]。Sudová 等[62]从污染基质中分离出来的AMF 密集定殖片段中的P和Pb的浓度显著高于非定殖片段,这表明真菌结构除积累重金属外还可以累积矿质营养元素。P 是植物生长所必需的营养元素,它在土壤中是不可移动的,大部分P 与有机分子或矿物表面结合,植物根系自身对P 的利用率较低[63]。AMF可以辅助吸收P元素,通过AMF辅助途径可提供高达80%的无机磷[64-65]。真菌菌丝一方面增加宿主植物根表面积以吸收更多的P,另一方面也能水解有机磷。菌丝吸收无机磷后将其运送到根内丛枝结构中,从而与宿主交换营养物[66]。除此之外,AMF 可以通过调控磷酸盐转运蛋白基因表达来加速宿主植物对磷酸盐的吸收,Gao等[67]的研究表明,根内球囊霉菌与棉花共生,可使棉花根和叶中磷酸盐转运蛋白家族基因激活和上调,棉花生物量中的P 含量提高了43.27%。AMF 也可以帮助植物吸收N、Ca、S、K、Zn 等其他营养元素。通过对上述元素的吸收,植物中的重金属相对含量能够减少,这一过程可以解释为“生长稀释”效应[48,68-70]。

图1 AMF增强宿主植物重金属抗逆性机制示意图(根据陈保冬等[58]修改)Figure 1 Mechanism of AMF enhancing heavy metal resistance in host plants(revision after Chen et al[58])
AMF 刺激植物根系分泌的有机酸等物质影响土壤微生物群落的组成,间接改变宿主植物根系周围土壤的理化性质,改善土壤养分[71],并通过促进植物对营养元素的吸收稀释植物体内重金属含量,达到解毒的目的。Jansa等[72]曾表明AMF 与土壤微生物之间是互利共生的关系。AMF 菌丝分泌物优先被菌丝体表面或附近的特定微生物群落利用,而与AMF 相关的微生物通过微生物介导的氮矿化、氮固定和磷增溶作用,又为真菌提供专属的氮磷等其他不可利用的养分。袁丽环等[73]的研究表明翅果油树幼苗接种AMF后,根面上的细菌、放线菌、固氮菌的数量显著增加,植物可吸收利用的氮磷含量增高。
植物体内的转运蛋白在限制重金属吸收过程中发挥着重要作用,AMF 定殖可以影响部分转运蛋白的编码基因表达[74-75]。这些蛋白包括锌转运蛋白、金属硫蛋白、90kD 热休克蛋白以及谷胱甘肽转移酶等[76]。上述蛋白的基因表达受到抑制会降低植物对重金属的吸收程度,从而达到解毒的目的。Sun 等[31]在Se 胁迫的土壤中接种AMF,发现AMF 与玉米共生降低了Se 吸收相关离子转运蛋白基因(ZmPht2、ZmNIP2;1、ZmSultr1;3)的表达,Se 从土壤到玉米根系的跨膜转运过程受到抑制。除Se 外,也有研究发现,AMF 可以降低As、Cd、Cu、Zn 等相关离子转运蛋白基因的表达[77]。
3.2 抗氧化机制
当植物受到重金属胁迫时,其体内分泌的抗氧化酶等渗透调节物质可以清除金属诱导的活性氧化自由基(ROS),减轻重金属对植物的氧化损害,提高植物的耐性。在金属胁迫下,植物体内的抗氧化酶含量降低,AMF 可以调节抗氧化酶系统,减少植物细胞内ROS 的积累,减轻膜脂过氧化程度,确保物质运输和能量传递[78]。杨玉荣[79]通过对比盆栽实验,发现AMF接种明显增强了Pb胁迫条件下刺槐幼苗叶片中超氧化物歧化酶、谷胱甘肽过氧化物酶、抗坏血酸过氧化物酶活性,AMF 通过促进宿主植物的抗氧化酶活性提高植物的抗逆性。
3.3 螯合重金属机制
重金属污染程度较高的土壤中,AMF 自身细胞内外及宿主植物根系都会分泌相关物质对重金属进行抵御,以增加土壤中重金属的稳定性,减少对植物的毒害作用。AMF 分泌物(如细胞壁分泌的黏液、聚磷酸盐、有机酸根离子、无机酸根离子等)可以螯合过量的重金属,阻碍重金属的迁移[80]。重金属胁迫下,AMF 也能刺激植物根系有机酸的分泌。这些分泌物包括氨基酸、糖类、酚类、次生代谢产物等低分子量分泌物和多糖、蛋白质等高分子量分泌物[79]。根系有机酸具有一个或多个羟基、氨基或羧基等活性官能团,在土壤-根界面与重金属离子发生络合-解离作用,活化土壤中的营养元素,将离子态的金属转变为毒性较低或者无毒的螯合态,从而降低重金属离子对植物细胞的毒害作用[81]。一些超累积植物已被证实可以通过释放有机酸来缓解重金属胁迫,如超累积植物叶芽鼠耳芥根部分泌的苹果酸和柠檬酸可螯合Zn 离子[82]。此外,有机酸对重金属的另一作用为改变根际土壤pH,进而影响土壤中重金属的有效性。Claroideoglomus etunicatum真菌侵染玉米可显著促进根系分泌有机酸,从而降低pH 及As 的生物有效性,抑制玉米对As 的吸收,但该菌作用于蜈蚣草会得到相反的结果[83]。张慧娟[84]发现接种AMF 能降低刺槐根系分泌有机酸的总量,从而显著提高根际土壤pH,土壤中离子交换态Pb 含量随pH 升高而减少,进而减轻了Pb 胁迫对刺槐的毒害作用。上述分析表明,AMF对植物根系有机酸分泌的影响效果并不一致,因此在评价其影响时应该考虑AMF 种类、宿主植物类型、土壤自身理化性质等多方面因素。
AMF 还可以分泌一种顽疏水、耐热、顽抗的糖蛋白——球囊霉素相关蛋白(Glomalina-related soil protein,GRSP),它包含在菌丝和孢子壁中[85],在菌丝和孢子衰老后被释放到土壤和沉积物中。GRSP的有利位置可以帮助菌丝黏附在污染物表面,有助于将重金属固定在土壤-菌丝界面[86],图1 中④过程可以表示重金属被阻滞在菌根共生结构中。González-Chávez等[87]分别用柠檬酸、硼酸和盐酸多次解吸与GRSP 溶液结合的Cu,发现Cu 很难被全部解吸下来,解吸后Cu 含量占解吸前含量的7.2%~12.0%,这说明球囊霉素对土壤中重金属有强化学络合作用,可降低这些元素的生物可利用性和毒性风险。Wang 等[88]通过主成分分析表明,9 种重金属的综合指标与GRSP 含量呈正相关,GRSP 的大量分布可以提高重金属的固定潜力[89]。有机酸及GRSP可改变重金属的形态从而降低重金属在土壤中的迁移及其生物有效态含量,如图1中⑤过程所示。
3.4 吸附固持重金属机制
AMF 的根外菌丝能够吸附部分重金属(图1中的③过程)。利用菌丝与植物根系直径差异悬殊这一特点,可以采用玻璃珠分室培养系统收集菌丝,研究菌丝对重金属的吸附。陈保冬等[90]发现离体真菌菌丝能够吸附Mn、Zn 和Cd,吸附量分别为自身干物质量的1.6%、2.8%和13.3%。AMF 菌丝通过自身吸附作用将重金属阻拦在植物根系外,减少进入植物体内的重金属含量,即AMF 的“过滤机制”。同时庞大的菌丝体系可以增大植物根系与土壤接触位点,增加对重金属的吸附量[91]。
AMF 菌丝中存在的转运蛋白在缓解重金属毒害及重金属区域化过程中起到重要作用。从Glomus intraradices中分离出来的GintABC1基因可以诱导ATP 结合子转运蛋白将重金属(如Cu、Cd 等)络合物转运至液泡中[92];其根外菌丝中的Zn 转运蛋白与AMF 缓解Zn 毒害及Zn 的区域化有关[7]。当重金属进入根细胞后,生物体常会将重金属稳定在某一特定部位从而降低关键生理功能部位重金属含量,在这一过程中部分重金属限制基因开始发挥作用,AMF 的存在也可以调控植物相关基因的表达。如HMA3基因可以将Cd 螯合到根细胞的液泡中,从而降低Cd 从根到地上部分的易位率[93-94]。接种摩西管柄囊霉菌后HMA3基因表达上调,这与根中Cd 含量较高,地上部分Cd 含量较低的结果一致[95]。表2 总结了接种AMF对相关转运蛋白基因表达调控的结果,证明了AMF可以通过阻滞重金属进入细胞、降低植物对重金属的吸收转运、根部细胞螯合重金属、刺激宿主合成植物螯合素等途径减缓重金属毒性。但没有直接证据表明AMF 可以直接调控植物体内相关基因表达,这也可能是接种AMF 后植物体的生理响应对相关基因的表达产生了影响。部分研究认为AMF 菌丝分隔重金属,降低植物重金属含量,同时也会抑制这些转运基因的表达。Motaharpoor 等[104]的研究表明,接种AMF会使污染土壤中植物螯合素合酶合成基因(MSPCS1)、金属硫蛋白合成基因(MSMT2)、两种转运基因(MsIRT1和MsNRAMP1)的转录丰度降低,这可能是菌丝的金属隔离使根细胞中的Cd 浓度不足,无法诱导上述基因的表达。AMF 对重金属吸收相关离子转运蛋白基因表达的调节作用可能与不同植物在不同生长时期和不同环境条件下的特定重金属含量有关。到目前为止,关于AMF 对重金属胁迫下植物吸收相关离子的分子学机制研究尚在起步阶段,AMF 对重金属的分子响应常伴随植物体内重金属含量和矿物质营养状况的变化,因此难以判断AMF 是直接调控还是间接调控基因的表达[58]。明确AMF 对于重金属胁迫下基因表达的直接作用,系统阐述AMF 增强植物重金属抗性分子学机制是未来要开展的工作。

表2 AMF调控相关转运蛋白基因表达的结果Table 2 Results of AMF regulating gene expression of related transporters
4 总结与展望
(1)随着土壤及农产品重金属超标问题的出现,土壤污染治理技术也得到重视。丛枝菌根真菌(AMF)强化植物修复重金属污染是一种绿色健康、经济有效的修复手段。但目前该技术仍有很多局限,如修复时间较长,对于复合污染土壤修复效果不佳,对土壤肥力、水分的自然和人为条件要求较高,这些缺点导致大部分研究只是停留在实验室阶段,并不能投入到实际应用中。如何解决上述缺陷,是目前研究的重点和难点。
(2)AMF 增强植物在重金属胁迫下的耐受机理十分复杂且受到诸多因素的影响,本文只是简单地综述了其中几个重要因素,对于此方面的研究仍在继续。此外,分子生物学的发展有助于AMF修复土壤重金属机理的研究。虽然目前也有一些报道从分子生物学角度分析了AMF 的作用机制,但是AMF 是否直接作用于植物基因表达的调控,又是怎样诱导植物分泌某些聚合物的问题目前尚不清楚,有待继续研究。
(3)目前,AMF 的离体培养以及工业化菌剂的制备仍是亟待解决的技术难题,解决这一问题将会对联合修复技术在实际应用中的发展提供帮助。
