
基于分子与形态证据的傣药“傣百解”基原考证
2023-03-13 07:47李金月谭运洪李海涛岩罕单龚燕雄肖云学郁文彬
广西植物 2023年1期
李金月, 谭运洪, 李海涛, 岩罕单, 龚燕雄, 肖云学, 郁文彬*
( 1. 中国科学院西双版纳热带植物园 综合保护中心, 云南 勐腊 666303; 2. 中国科学院大学, 北京 100049; 3. 中国科学院东南亚生物多样性研究中心, 云南 勐腊 666303; 4. 中国医学科学院、北京协和医学院药用植物研究所云南分所, 云南 景洪 666100; 5. 西双版纳州傣医医院, 云南 景洪 666100; 6. 云南省热带作物科学研究所, 云南 景洪 666100; 7. 中国科学院西双版纳热带植物园 园林园艺部, 云南 勐腊 666303 )
“傣百解”为国药准字品种“雅解片”,傣族称为“雅解先打”,也俗称“大百解”,意译为“解百毒的药”(云南省食品药品监督管理局, 2005)。“解药”是傣族医药体系中最为重要的组成部分,也是区别于其他民族药的特色药物。“傣百解”是傣医常用的解药之一,在中国云南省傣族聚居的西双版纳州、德宏州、新平县、元江县等地,以及老挝、缅甸等国家广泛使用(中国科学院中国植物志编辑委员会,1977)。“傣百解”可用于清热解毒、止咳平喘,具有抗肿瘤、降血压、抗炎等功效,还常用于治疗药物或食物中毒等(国家中医药管理局, 2005; Wang et al., 2018)。根据已有的文献报道,从“傣百解”基原植物通光散(Marsdeniatenacissima)中已分离出190余种化学成分,主要包括甾体类化合物、三萜类化合物、有机酸化合物和生物碱化合物等(廖矛川等, 2016),其中固醇类化合物是抗肿瘤和耐化疗逆转作用的特有物质和主要活性成分(Wang et al., 2018)。
“傣百解”用药历史悠久,现已被西双版纳州傣医院用作多个院内制剂的组方药材,如百解胶囊、雅解嘎罕、版纳凉剂等。其中,百解胶囊在傣医临床应用最为广泛,具有良好的临床疗效和开发应用前景(杨莲等, 2021)。由于“傣百解”具有重要药用价值、市场前景良好、开发潜力大,因此需要加强对“傣百解”基原植物野生药材资源的保护,使该药用植物资源得到可持续利用。研究和开发利用傣药的首要环节就是对傣药药材品种进行整理与考证,这对澄清基原植物和确保傣药药材质量以及用药安全具有重要意义(段宝忠等, 2015)。
基原鉴定是药材鉴定的核心内容之一。自然条件的限制以及医著记载不详等原因,导致各地的傣医在一些药材基原的认识上存在分歧,其中对“傣百解”也存在基原不清和错误鉴定的问题。《云南省中药材标准》(2005年版)记载了“傣百解”基原是萝藦科(Asclepiadaceae)的苦绳(Dregeasinensis)的根(云南省食品药品监督管理局, 2005)。管艳红等(2012)采用石蜡切片对“傣百解”药材和苦绳的根进行横切面结构比较发现两者存在明显差异,认为“傣百解”和苦绳是两种不同的植物。同年,管志斌等(2012)利用ISSR分子标记分析“傣百解”和几种牛奶菜属(Marsdenia)药用植物的亲缘关系,发现“傣百解”与通光散的亲缘关系最近。随后,李海涛(2014)等通过综合文献查证、形态学解剖、生药学等证据对“傣百解”的基原进行了考证,发现“傣百解”基原植物为苦绳是早期的错误鉴定,认为“傣百解”的基原植物应该是萝藦科牛奶菜属的通光散。值得注意的是,在最新的被子植物分类系统中已将萝藦科归并到夹竹桃科(Apocynaceae)作为1个亚科,即萝藦亚科(Asclepiadoideae) (The Angiosperm Phylogeny Group, 2016)。
目前,由于“傣百解”药材的研究主要在形态学、生药学和化学成分等方面且研究比较零散(管艳红等,2012;李海涛等,2014;廖矛川等,2016),所以整合形态特征与分子证据以开展“傣百解”基原植物考证具有重要意义。由于传统的药材鉴定方法具有一定的局限性,尤其是近缘种、易混种在没有花和果等关键特征的情况下分类鉴定难度比较大,因此容易出现错误鉴定。随着DNA测序技术的快速发展,分子鉴定方法因具有方便、快捷和准确等特点而被广泛应用于物种和药材鉴定(Yu et al., 2021)。本研究以澄清傣药“傣百解”基原植物为目的,利用栽培的“傣百解”基原植物材料、通光散以及牛奶菜族的其他植物样品,采用形态结合分子的方法,通过整合形态性状比较和系统发育树分析,探讨以下问题:(1) “傣百解”基原植物是否为通光散;(2) “傣百解”的近缘种有哪些;(3) 如何鉴别区分“傣百解”及其近缘种。
1 材料与方法
1.1 材料
新收集了夹竹桃科牛奶菜属5种共17份样品,包括栽培于中国医学科学院药用植物所云南分所南药园“傣百解”样品4份,以及南山藤属(Dregea) 2种7份样品和匙羹藤属(Gymnema)1种1份样品。具体样品来源和凭证信息如表1所示。采集植物样品的新鲜叶片经变色硅胶进行快速干燥,并低温保存,以备实验所需。

表 1 研究材料及其凭证信息Table 1 Study materials and voucher information

续表 1
1.2 DNA提取、测序和数据处理
对用硅胶干燥后的样品,先利用改良的十六烷基三甲基溴化铵(CTAB)法提取总DNA (Doyle & Doyle, 1987),再使用琼脂糖凝胶电泳和Nanodrop分光光度计评估DNA质量。对检测合格的DNA样品,使用Covaris 超声波破碎仪随机打断,随后通过末端修复、加A尾、加测序接头、纯化、PCR扩增、片段筛选等步骤,完成片段长度为350 bp的测序文库制备。对构建好的测序文库使用Qubit 2.0对浓度进行检测,定量范围为2~1 000 ng,依据溶度进行多样品混合,并利用Illumina Novaseq 6000进行双向150 bp读长测序。对测序原始数据(raw data)直接使用GetOrganelle软件包 (Jin et al., 2020) 进行叶绿体基因组和核糖体DNA从头(denovo)组装。选取长春花(Catharanthusroseus) 的叶绿体基因组(KC561139)和核糖体基因(HQ130657)序列作为参考序列,借助Geneious (Kearse et al., 2012) 对新测物种进行初步注释,相似性参数设置为70%,并结合ORF (open reading frame)手工调整注释结果,其中蛋白编码基因的注释依据参考序列和开放阅读框确定起始密码子和终止子位置。
1.3 系统发育分析
在Geneious软件中提取新测29份样品的叶绿体基因片段psbD-trnT和trnL-trnF,以及核糖体基因序列ITS(包括ITS1、5.8S、ITS2三个区),并从GenBank上下载已有的牛奶菜属42种、南山藤属1种以及匙羹藤属2种的psbD-trnT、trnL-trnT以及ITS序列,以吊灯花属(Ceropegia)植物锥顶吊灯花(Ceropegianilotica)作为外类群(表2)。对3个基因片段的序列分别利用MAFFT 7.450软件进行多序列自动比对 (Katoh et al., 2013),输入到Geneious软件将模糊的区域或空位进行删除或调整。使用贝叶斯推断法(Bayesian inference, BI)、最大似然法(maximum likelihood, ML)和最大简约法(maximum parsimony, MP)对3个片段分别进行单独和联合重建。对3个基因片段矩阵分别利用jModeltest 2 2.1.6,选择贝叶斯信息量准则(Bayesian information criterion, BIC)(Darriba et al., 2012) 筛选出序列矩阵最佳核苷酸替代模型,并采用Mrbayes 3.2.7(Huelsenbeck & Ronquist, 2001)进行贝叶斯系统发生分析。采用最大似然法时选用RAxML-HPC2 on XSEDE 8.2.12工具(Alexandros, 2014) ,选择GTR+CAT模型,并设置1 000次靴带值估算分支的支持率(bootstrap support value, BS),使用CIPRES网站在线构建ML树。采用最大简约法时使用MEGA 10.2.6软件(Kumar et al., 2018),使用子树修剪和嫁接(Subtree-Pruning-Regrafiting, SPR)算法进行分析获得MP树(Nei & Kumar, 2000)。构建系统进化树时使用FigTree 1.4.1 (Price et al., 2009) 进行美化。

表 2 其他取样物种和序列GenBank登录号Table 2 List of taxa and GenBank accessions of other sequences used in this study
1.4 形态特征比较
将采集到的“傣百解”样品与通光散的模式标本进行形态观察,并进一步比较分析在系统树上与“傣百解”亲缘关系最近的物种,观察和比较“傣百解”基原植物与其姐妹种之间的形态特征差异。
2 结果与分析
2.1 基因序列特征
核糖体ITS矩阵的长度为768 bp,包含334个变异位点(variable sites),其中202个为位系统发育信息位点(parsimony-informative site)。叶绿体片段psbD-trnT矩阵的长度为1 676 bp, 包含247个变异位点,其中有51个为系统发育信息位点。叶绿体片段trnL-trnF矩阵的长度为1 006 bp,包含142个变异位点,其中有47个为系统发育信息位点。psbD-trnT+trnL-trnF+ ITS序列矩阵全长3 326 bp,包含597个变异位点,其中有305个为系统发育信息位点。
2.2 系统发育分析
同一基因片段矩阵采用贝叶斯法、最大似然法和最大简约法构建的系统发育树的拓扑结构基本一致(图1)。3个片段联合矩阵得到的系统发育树比单基因构建的系统发育树能更好地澄清物种之间的关系(图1:D)。在单基因系统发育树中,核糖体ITS矩阵(图1:C)比叶绿体psbD-trnT矩阵 (图1:A)和trnL-trnF矩阵(图1:B)能更好地将物种进行区分和鉴别。
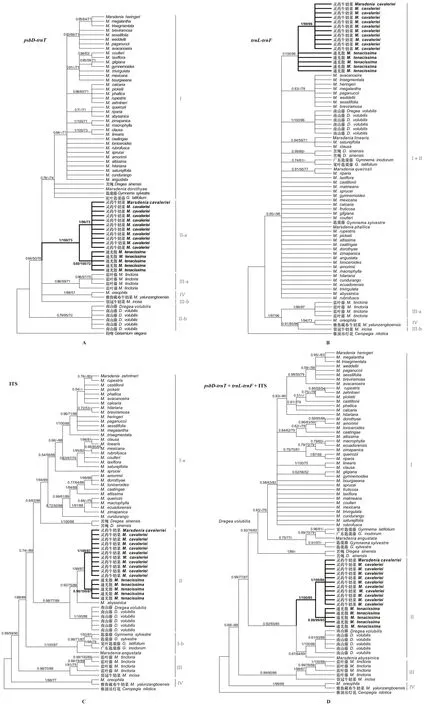
罗马数字表示主要分支,分支节点上的数值以BIpp/MLBS/MPBS的方式表示贝叶斯后验概率/最大似然树自展支持率/最大简约树自展支持率,后验概率不足0.50或支持率不足50的均不标记。Roman numbers indicate main clades, and data upon each node (BIpp/MLBS/MPBS ) indicate Bayesian inference posterior probability / maximum likelihood bootstrap values/ maximum parsimony bootstrap values, and PP below 0.50 and BS below 50 are unlabeled.图 1 基于叶绿体片段 (psbD-trnT、trnL-trnF) 和核基因ITS构建“傣百解”及其近缘种的系统发育树Fig. 1 Phylogenetic reconstruction of “Dai-Bai-Jie” and its related species based on two plastids (psbD-trnT and trnL-trnF ) and nuclear ribosome DNA ( ITS ) data
联合数据矩阵系统发育树结果表明,牛奶菜属不是一个单系类群,即南山藤属和匙羹藤属物种与牛奶菜属混合在一起。系统发育分析显示,牛奶菜族分为4个主要分支,其中雅鲁藏布牛奶菜(Marsdeniayarlungzangboensis)与(M.oreophila)构成了分支IV (BIPP/MLBS/MPBS=1.00/99/68),是最早分化出的分支,与其他3个分支构成姐妹关系 (BIPP/MPBS= 0.68/88);随后是蓝叶藤(M.tinctoria)和裂冠牛奶菜(M.inicisa)组成了分支III (BIPP/MLBS/MPBS= 0.99/80/86),并较好支持的是分支I + 分支II的姐妹群(BIPP/MLBS/MPBS= 0.99/77/87)。分支II包括了“傣百解”样品,以及通光散、灵药牛奶、南山藤(Dregeavolubilis)和Marsdeniaabyssinica(BIPP/MLBS/MPBS= 0.92/93/85),其中“傣百解”基原植物样品先与通光散样品聚在一起形成单系 (BIPP/MLBS/MPBS= 0.92/93/85),再与灵药牛奶菜(M.cavaleriei)互为姐妹类群,所形成的分支节点支持率较高 (BIPP/MLBS/MPBS= 1.00/100/85),这种关系也得到了单基因片段数据的支持;另外,南山藤与M.abyssinica形成了1个分支(BIPP/MLBS/MPBS= 0.67/64/69)。分支I包括了美洲产的牛奶菜属物种,以及匙羹藤(Gymnemasylvestre)、宽叶匙羹藤(G.latifolium)、广东匙羹藤(G.inodorum)和苦绳 ( BIPP/MLBS/MPBS= 0.92/76/82),并且非单系的匙羹藤属分支包括Marsdeniaabyssinica。
2.3 形态学分析
2.3.1 “傣百解”与通光散模式标本的比较 通过比较“傣百解”基原植物的凭证标本与通光散的模式标本(图2),发现两者的叶片和花形态特征基本一致,即茎干和叶片表面密被淡黄色柔毛,叶片在顶端出急骤趋于尖狭,叶基部深心形,近圆形弯缺;花序均为多歧聚散花序,花外部密被绒毛,花冠近钟状,花萼5瓣深裂,裂片肉质膨胀,柱头宽圆筒状凸起等。因此,进一步证明了“傣百解”的基原为通光散。

图 2 “傣百解”凭证标本(A)与通光散模式标本(B) 的比较Fig. 2 Comparisons of a voucher specimen of “Dai-Bai-Jie” (A) and the type specimen of Marsdenia tenacissima (B)
2.3.2 通光散与灵药牛奶菜的形态比较 “傣百解”基原植物与灵药牛奶菜在系统发育分析中是姐妹类群,但通过比较两者的形态性状,发现它们两者的花冠颜色、叶型、叶面、茎干等相关性状存在差异(图3, 图4)。详细的特征区要点如表3所示。

图 3 通光散(A, B) 与灵药牛奶菜(C, D) 植物图Fig. 3 Photos of Marsdenia tenacissima (A, B) and Marsdenia cavaleriei (C, D)

A, C, E, G, I. 通光散各部分的形态结构; B, D, F, H, J. 灵药牛奶菜各部分的形态结构。A, C, E, G, I. Morphological sturcture of each part from M. tenacissima; B, D, F, H, J. Morphological structure of each part from M. cavaleriei.图 4 通光散与灵药牛奶菜在叶子、花、茎、根及其横切面的形态比较Fig. 4 Morphological comparisions of Marsdensia tenacissima and M. cavaleriei about leaf, flower, stem, root and its cross section

表 3 通光散与灵药牛奶菜形态特征的比较Table 3 Comparison of morphological characters between Marsdenia tenacissima and M. cavaleriei
3 讨论与结论
“傣百解”被收录于2005年版的《云南省中药材标准》时,其基原植物记载是南山藤属植物苦绳。基于叶绿体基因psbD-trnT和核糖体基因ITS序列单独分析和3个片段的联合分析表明,“傣百解”基原植物样品和通光散样品形成1个单系分支,并且与南山藤和M.abyssinica一起属于分支III。因此,系统发育分析支持“傣百解”的基原植物是通光散,并与灵药牛奶菜成姐妹关系。在trnL-trnF序列的系统发育树中“傣百解”基原植物和通光散样品未聚成1支,而灵药牛奶菜所有样品形成1个单系,这可能是由于该基因片段的变异位点较少所造成,因为其他物种的关系也未能得到较好的解析。在系统发育树上,苦绳与美洲的牛奶菜属物种和匙羹藤属物种聚在一起属于分支I,与“傣百解”基原植物关系较远。结合《云南省中药材标准》后面附录的植物照片和药材照片,进一步确认“傣百解”的基原就是通光散,这与早期的研究和考证结果一致(李海涛等, 2014)。
值得关注的是,从2012年开始与“傣百解”基原植物相关的考证研究主要基于形态分析和显微鉴定(管艳红等, 2012)。由于传统的鉴定方法会存在局限性,容易受物种本身所处的环境和生长期的影响,因此需要依赖鉴定者的经验,进而判断鉴定结果的准确性。已有的研究使用叶绿体基因组序列psbD-trnT、trnL-trnF、核糖体ITS和ETS较好地解析了牛奶菜族的系统发育关系 (Espírito- Santo et al., 2019), 以及利用叶绿体基因rbcL和核糖体ITS2序列能很好地区分一些牛奶菜属物种与其近缘类群(张忠廉等, 2013),特别是核糖体ITS2序列,在DNA条形码 (DNA barcoding) 中已被推荐用于物种鉴定的通用条形码,并已用于药材市场中通光散及其掺假品的鉴别 (Yu et al., 2018)。当前,植物DNA序列和系统发育的分析方法应用于药材的分子鉴定已成为植物分类和物种鉴定研究的热点(陈士林等, 2007)。不过,对于亲缘关系接近的物种或姐妹物种,由于单个基因片段因变异度低而使得物种鉴定率低(倪梁红等, 2014),因此采用变异位点数较多的基因片段或多片段组合可有效提高物种鉴定率(张越等, 2021)。本研究中的叶绿体片段trnL-trnF由于序列变异位点较少,只占所分析序列长度的14.11%,因此系统发育分析显示许多物种之间的关系未能得到解决。相比而言,核糖体ITS具有较多的变异位点,在系统发育树中能将大多数物种区分开。因此,本研究推荐使用核糖体ITS,或者使用核糖体ITS与叶绿体片段进行联合分析,用于“傣百解”基原植物的分子快速鉴定(倪梁红等, 2014)。
本研究形态特征比较结果显示,“傣百解”基原植物的凭证标本与通光散的模式标本在叶子形态、花的结构以及植株的被毛等特征基本一致。形态上,“傣百解”基原植物和通光散的茎都密被淡黄色柔毛,叶两面被绒毛,叶顶端急尖,基部深心形,近似圆形弯缺;茎被毛多,为圆柱形,折断后有白色乳汁;花序多歧分支,伞形聚伞花序,花冠近钟状,裂片肉质膨胀,展开;柱头凸起,为宽圆筒状,被花药附属物覆盖 (William et al., 1819; Li et al., 1995)。本研究结果与李海涛等(2014)对“傣百解”基原植物考证结果一致,进一步确定了“傣百解”的基原植物是通光散。从比较通光散与近缘种灵药牛奶菜的形态结果可以看出,两者具有明显的形态差异:(1)灵药牛奶菜的叶子基部没有形成圆形弯曲,叶被毛稀疏,表面较为光滑;(2)茎为扁圆柱形,被毛沿着狭窄的两侧生长,花冠颜色与通光散不同,为橙红色;(3)花冠裂片较浅,张开的幅度较小,直立后稍外展;(4)根表面粗糙,不为粉质,不易剥落。因此,通光散和灵药牛奶菜可依据根、茎、叶、花的形态性状差异进行物种鉴别(图 4)。西双版纳傣药“傣百解”一直以来备受人们关注,其基原考证首次从分子系统学角度获得进一步的澄清,将植物的形态特征与其DNA序列相结合可以很好地对物种及其近缘种进行鉴别,并且鉴别效果比单一的传统鉴定法更佳(金海湘等, 2019; 苏畅等, 2019; 姚纲和薛彬娥, 2021)。
傣药,不仅是我国四大民族药之一,也是我国医药文化不可或缺的组成部分。民族医药形成于少数民族区域,长久以来,当地的民众都使用民族药来治疗疾病和保健身体。因其具有良好的效果而得以世代相传(张梦娜, 2018)。为了使民族药物资源和民族生物文化多样性得到保护和持续利用,我们需要对一些常用的傣药进行系统的整理和物种基原考证,从而能对傣药的基原植物做准确的鉴定,这是确保临床上准确、安全用药的前提(龙春林, 2013)。由于传统的生药鉴定法有一定的局限性,主要依赖于经验,主观性较强,对于形态学上难以区分的样品,不同的人依据不用的性状得到的分类结果也不尽相同,因此结合分子系统学的证据会更有助于物种的鉴定。同时,系统发育分析有助于物种之间亲缘关系的梳理,既可能寻找到近缘替代种,又可为“傣百解”基原植物的种质资源综合评价、系统进化等研究提供有效的科学依据,这对品种整理、鉴定以及保护和开发利用具有重要的意义(段宝忠, 2017)。
猜你喜欢
小猕猴智力画刊(2022年3期)2022-03-29
小天使·一年级语数英综合(2021年4期)2021-05-08
世界科学技术-中医药现代化(2021年10期)2021-03-02
小天使·一年级语数英综合(2020年11期)2020-12-16
世界科学技术-中医药现代化(2020年2期)2020-07-25
科学大众(中学)(2019年3期)2019-05-17
汽车观察(2018年10期)2018-11-06
中国中药杂志(2017年13期)2017-07-31
科技知识动漫(2017年1期)2017-02-06
作文周刊·小学一年级版(2016年1期)2016-08-12
