
白念珠菌对氟康唑耐药机制的研究
2023-03-09 01:32:44封小川元航郑莹莹白江苗张欠欠
中国真菌学杂志 2023年6期
封小川 元航 郑莹莹,3 白江苗 张欠欠
(1.延安大学医学院,延安 716000;2.延安市人民医院,延安 716000;3.延安大学附属医院,延安 716000)
近年来,世界范围内以白念珠菌感染为主的侵袭性念珠菌病呈上升趋势,全球SENTRY抗真菌监测显示,在欧洲,白念珠菌引起的感染占念珠菌属的52.5%、亚太地区为46.0%、拉丁美洲为43.9%、美国和加拿大为42.7%[1],我国流行病学调查显示[2-3],白念珠菌引起的感染占念珠菌属的65%,个别地区甚至大于70%,在免疫功能低下者和植入医疗设备的健康人中白念珠菌的感染率居首位,感染病死率高达68.9%[4],美国每年也会因置入导管引发的白念珠菌感染造成约10万人死亡[5]。美国感染病学会(IDSA)指出氟康唑是既往非粒细胞缺乏念珠菌血症患者初始治疗及ICU中高危患者侵袭性念珠菌感染预防的首选药物[6],在发展中国家,三唑类药物(如氟康唑等)是治疗侵袭性念珠菌感染尤其是白念珠菌感染的主要药物[7]。但随着氟康唑在临床上的长期广泛应用,致使白念珠菌对氟康唑的耐药性不断增强,据美国疾控中心(CDC)统计,在感染念珠菌患者的血液样本中,约有7%对氟康唑耐药[8]。全球SENTRY监测显示[9],白念珠菌对氟康唑的耐药率为11.9%,我国白念珠菌对氟康唑的耐药率小于6%,剂量依赖性敏感(SDD)率为4.35%[10],这势必给临床治疗白念珠菌引发的疾病过程中带来严峻考验。
文献报道显示[11-13],白念珠菌对氟康唑耐药机制主要是靶酶编码基因ERG11的突变和外排泵编码基因过表达,但由于不同地区白念珠菌的感染及其耐药不同,其耐药机制亦不尽相同,因此,本文通过研究白念珠菌对氟康唑耐药相关基因ERG11、CDR1、CDR2和MDR1的表达水平及其与耐药的关系,探究延安地区白念珠菌对氟康唑的耐药机制,对管理目前有限的抗真菌药物,以及为患者提供精准治疗服务至关重要。
1 材料和方法
1.1 材料
菌株 收集延安市两家三甲医院送检并经鉴定为白念珠菌且对氟康唑耐药和敏感的菌株各10株,耐药菌株编号C1-C10,敏感菌株编号C11-C20。标准质控菌株:白念珠菌ATCC 90028。纳入标准:①经质谱鉴定为白念珠菌;②根据美国临床实验室标准化研究所(CLSI)判断标准[14],敏感(S):MIC≤8 μg/mL,耐药(R):MIC≥64 μg/mL,药敏结果复核为白念珠菌对氟康唑耐药及敏感菌株;③所有操作均符合实验室操作标准。
主要试剂和仪器 YeastOne plate真菌药敏板试剂盒(赛默飞世尔科技(中国)有限公司)(批号:280543)、Taq DNA聚合酶(北京百奥莱博科技有限公司)(批号:JN0015)、逆转录试剂盒(广州赛国生物科技有限公司)(批号:70060000)、罗丹明 6G (西亚化学试剂商店)(批号:20211115)、VITEK MS全自动快速质谱微生物检测系统(法国梅里埃公司)、ABI 7300 型全自动荧光定量PCR仪(美国ABI公司)、Cytomics 500流式细胞仪(美国贝克曼库尔特公司)、Nanodrop One 超微量分光光度计(赛默飞世尔科技(中国)有限公司)、Tanon3500型凝胶成像分析系统(上海天能有限公司)。
1.2 方法
ERG11基因扩增并测序 白念珠菌DNA的提取:菌株培养、离心后,提取出白念珠菌DNA,与5 μL 样品混合均匀后进行电泳约55 min,应用电脑成像系统进行凝胶相片拍摄。以GenBank公布的白念珠菌标准菌株ERG11基因序列数据为准,PCR引物设计:ERG11F:5’-ATGGCTATTGTTGAAACTGTCATTG-3’ (第1~25位) 、ERG11R:5’- TTAAAACATACAAGTTTCTCTTTTT-TCC-3’(第1560-1587位)。
罗丹明6G在试验菌株中外排情况的测定 ①实验菌株经离心、预冷,重悬于PBS使葡萄糖耗尽,加入罗丹明6G后将试管放于冰面上,然后在菌液中加入PBS混合均匀、离心,用流式细胞仪进行分析记录,此时菌株的荧光强度设为外排前的起始荧光强度;②洗3次预冷PBS,将多余的罗丹明6G去除后分为两管,一管50 μL菌液加入1 mL PBS,另一管50 μL菌液加入1 mL PBS,离心加入PBS混合均匀,运用流式细胞仪分析,将未用罗丹明6G处理的菌株细胞的荧光强度作为自身荧光对照;③将摄取罗丹明 6G 之后的细胞荧光强度作为基底,分别减去加与不加葡萄糖外排之后的细胞荧光强度,进行数据分析。
白念珠菌外排泵基因表达的检测 白念珠菌外排泵基因引物序列见表1。
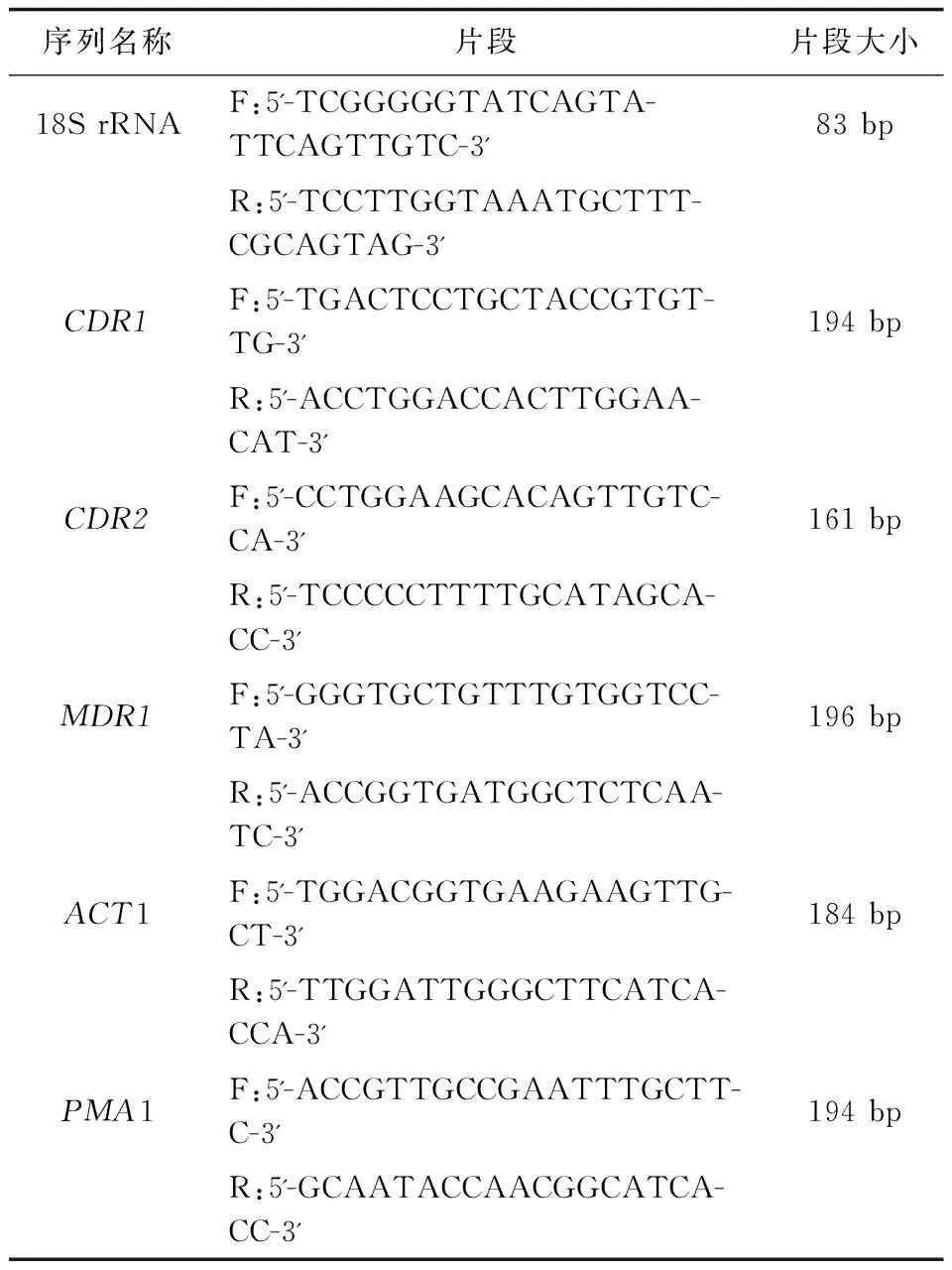
表1 白念珠菌外排泵基因引物
白念珠菌总RNA的提取 采用Trizol法提取白念珠菌总RNA,RT-PCR 扩增外排泵基因严格按照试剂盒说明书进行操作。PCR反应条件:94 ℃预变性3 min, 94 ℃变性10 s,55 ℃退火20 s,72 ℃延伸30 s,40个循环。收集荧光阈值(cycle threshold,Ct),将核糖体基因18S rRNA设定为内参基因,用相对定量△Ct表示(△Ct = Ct目的基因- Ct18S)并进行分析,其中△Ct值越小,相对基因表达量越高,反之则越低,见图1及图2。
1.3 统计学分析
2 结 果
2.1 ERG11基因的检测结果
实验菌株ERG11基因 PCR扩增的产物经纯化并测序,测序结果与 GenBank中的标准序列X13296进行比对分析,发现氟康唑耐药的C3号菌株第414位碱基发生了突变G→A,但并未引起第118位氨基酸甘氨酸(Gly)密码子的改变;氟康唑耐药的C7号菌株及对氟康唑敏感的C12号菌株的第518位碱基发生了突变C→G,引起第173位氨基酸密码子改变,致脯氨酸(Pro)变为精氨酸(Arg),见图3~6及表2。

图1 实验菌株RT-PCR溶解曲线

表2 白念珠菌ERG11基因突变及对应氨基酸改变
2.2 罗丹明6G的外排情况
结果显示,敏感株在摄取罗丹明6G后,未加葡萄糖几乎不发生外排,加葡萄糖后略有外排,但二者比较差异无统计学意义(P>0.05);耐药株在摄取罗丹明6G后,无论加葡萄糖与否均发生外排,加葡萄糖较未加葡萄糖的外排量增加,二者比较差异具有统计学意义(P<0.01);将耐药组株对罗丹明6G的外排与敏感组菌株对罗丹明6G的外排量进行分析,耐药组罗丹明6G的外排量明显高于敏感组,差异具有统计学意义(P<0. 01),见表3和图7。
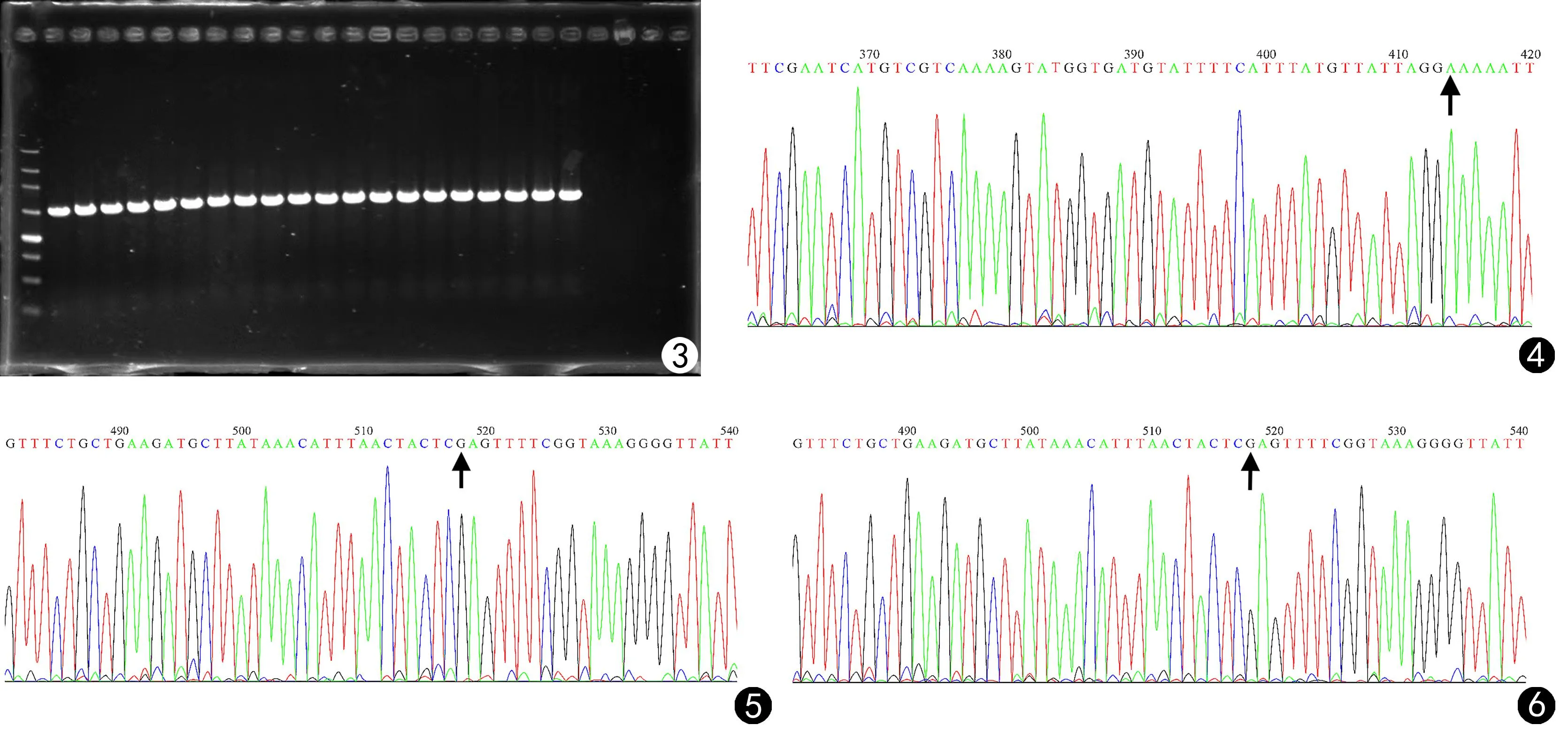
图3 白念珠菌ERG11基因扩增DNA凝胶电泳图
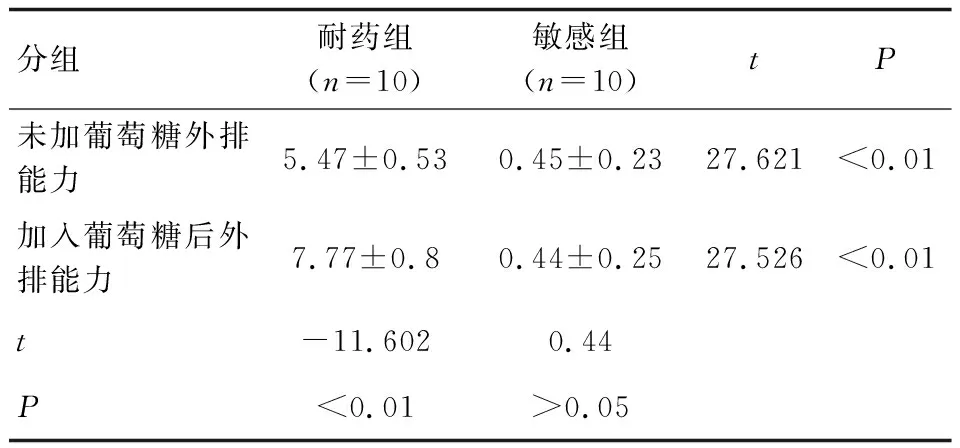
表3 白念珠菌耐药组与敏感组罗丹明6G外排实验结果比较
2.3 RT-PCR测定外排泵基因表达水平
统计学分析显示,耐药组和敏感组肌动蛋白相关基因ACT1和质膜H+-ATP酶编码基因PMA1的表达水平相比较,差异无统计学意义(P>0.05);耐药组CDR1和CDR2外排泵基因表达量较敏感组显著增高,差异有统计学意义(P<0. 01),见表4和图8。
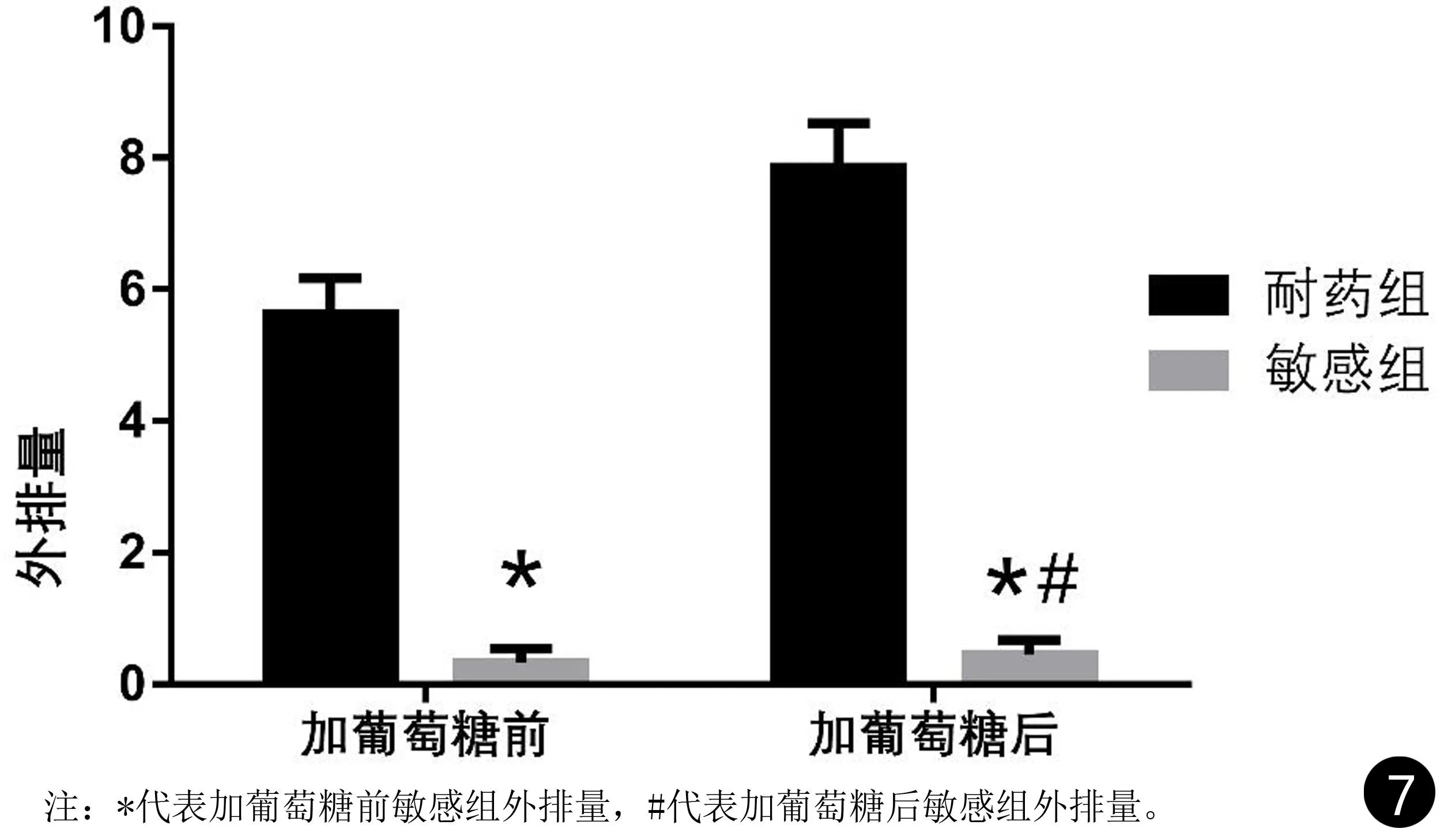
图7 敏感和耐药白念珠菌罗丹明6G外排实验结果

表4 白念珠菌主动外排泵基因在敏感组与耐药组相对表达水平的比较(△Ct值)

图8 敏感和耐药白念珠菌外排泵基因的表达量
3 讨 论
全世界每年由念珠菌引起的深部器官感染病例40万以上,病死率超过45%[15],尤其白念珠菌的发病率、致死率明显上升。唑类药物中的氟康唑是临床早期经验治疗白念珠菌感染尤其作为高危人群预防白念珠菌感染的首选药物,使其在人体内长期处于亚治疗水平,引起耐药性增加[16],白念珠菌耐药是目前临床治疗其感染失败的一个重要原因,研究报道,ERG11基因错义突变可使氟康唑抗真菌的作用靶点去甲基化酶CYP51的血红素表面结合位点发生变化,从而使氟康唑无法与白念珠菌有效结合而致耐药[17],而且白念珠菌细胞膜上的外排泵活力增强也是白念珠菌对氟康唑耐药的主要机制之一[18]。
本研究中靶酶编码基因ERG11的目的基因扩增测序中虽然引起氨基酸的改变,即耐氟康唑的C7号菌株及对氟康唑敏感的C12号菌株的第518位碱基发生了突变,引起第173位氨基酸密码子改变,致Pro变为Arg,但耐药及敏感菌株中均存在,虽为错义突变,但该突变没有发生在ERG11点突变的105-165、266-287、405-488氨基酸之间的3个“热点”区域[19],也不是既往文献报道的Y123F、K143R、F449V和G464S等多个与白念珠菌对氟康唑耐药有关的突变位点[20],推测这个位点的突变在白念珠菌对氟康唑耐药机制中为无意义突变,但此也不能表明该机制在本地区白念珠菌耐药性的产生中无作用,可能与样本量较少有关,尚需扩大样本量进行进一步研究。
罗丹明6G作为白念珠菌细胞膜上活性流出系统的底物,具有无毒、易检测、易吸收入射光的能量等优点,准确测量白念珠菌中罗丹明6G的积累浓度值有利于选择抗性菌株中CDR1、CDR2、MDR1等基因的过度表达。研究结果显示,加入荧光染料罗丹明6G后,耐药组罗丹明 6G的外排量明显高于敏感组,说明白念珠菌对氟康唑耐药为主动外排系统所致。为排除实验环境(如菌株的生长环境和RNA的抽提等)对外排泵基因的检测结果的影响,耐药组和敏感组加入肌动蛋白相关基因ACT1和质膜H+-ATP酶编码基因PMA1,比较其表达水平,差异无统计学意义(P>0.05),说明实验环境对外排泵基因的检测结果没有影响。耐药组CDR1和CDR2外排泵基因表达量较敏感组显著增高,差异有统计学意义(P<0.01),由此可知影响本地区白念珠菌对氟康唑耐药机制中最为重要的是其药物外排泵基因CDR1、CDR2过度表达,并且耐药组耐药基因CDR1、CDR2相对表达量均显著高于敏感株组,与文献报道结果相似[21-22]。分析原因主要是,CDR1的表达受顺式作用调控元件的控制,包括一个BEE(基础表达元件)、一个DRE(药物敏感元件)、两个SRE(甾醇调控元件)和一个NRE(负调控元件),而CDR2的表达仅受DRE的控制,在这些不同的元件中,只有DRE参与必需的CDR1和CDR2的高表达和对氟康唑外排的上调相反,MDR1基因不具有DRE元件,并且MDR1启动子也不直接与氟康唑反应[21]。
综上所述,本地区白念珠菌的耐药机制主要与外排泵基因过度表达相关,未发现靶酶编码基因有意义的突变,由于白念珠菌对氟康唑耐药现象的产生并非由于单一机制的调控,且临床很多患者存在滥用抗真菌药物的现象,导致白念珠菌对氟康唑的耐药性不断提高,而新型抗真菌药开发缓慢,改善白念珠菌对氟康唑的耐药问题迫在眉睫,因此,我们急需进一步探索白念珠菌致病机制和耐药机制,寻找新的药物治疗靶点,探索新的治疗体系,提高患者的生存率。
猜你喜欢
工程与建设(2019年3期)2019-10-10 01:40:36
基础医学与临床(2018年2期)2018-02-12 13:12:30
科技创新导报(2016年31期)2017-03-30 09:15:27
西南军医(2016年1期)2016-01-23 02:22:24
中国卫生标准管理(2015年5期)2016-01-14 05:17:06
山东医药(2015年13期)2016-01-12 00:39:39
化工进展(2015年3期)2015-11-11 09:09:13
中国医疗美容(2015年1期)2015-07-12 10:06:19
湖北理工学院学报(2015年1期)2015-02-27 15:02:28
山西大同大学学报(自然科学版)(2014年2期)2014-01-23 01:55:22
