
不同昼夜温差对赤霞珠葡萄果实香气的影响及转录组分析
2023-03-09 09:48:24张亚红乔振羽王亚楠黄嘉俊
核农学报 2023年4期
张 乐 张亚红 乔振羽 王亚楠 陈 璐 周 娟 黄嘉俊
(1宁夏大学农学院,宁夏 银川 750000;2宁夏大学食品与葡萄酒学院,宁夏 银川 750000)
香气作为葡萄果实品质评价中权重赋值最高的指标之一,对果实本身及其加工品影响重大,也是促进葡萄与葡萄酒市场消费的重要因素之一[1]。近年来,为筛选影响果实香气品质的关键气象因子,众多学者对国内主要酿酒葡萄产区的气候特征和果实品质特征进行了大量研究并总结,指出昼夜温差是影响果实香气品质的关键气象因子[2]。
葡萄果实香气物质合成途径主要有脂肪酸代谢途径、氨基酸代谢途径和异戊二烯代谢途径[3]。脂肪酸代谢途径是以不饱和的亚油酸和亚麻酸为前体物,在氧合酶的催化下合成C6醛及相应的醇、酯类[4];氨基酸代谢途径产生的物质一般包括脂氨基酸和芳香族氨基酸,其中脂氨基酸是果实香气成分中支链脂肪族醇、醛、酮和酯类物质的主要来源,芳香族氨基酸则是挥发性酚、醚类和某些含芳香环类香气物质的前体物;异戊二烯合成代谢途径主要以乙酰辅酶A为前体合成倍半萜类化合物为主[5]。大量研究表明,造成葡萄香气多样性的原因主要是不同地域的“风土条件”,包括气候条件、土壤条件、栽培措施等,这些因素及其相互作用都会影响葡萄和葡萄酒的质量,其中气候条件对葡萄香气影响的研究得到广泛关注[6]。研究发现,春季温度显著影响采收期美乐葡萄的3-异丁基-2-甲氧基吡嗪等吡嗪类和C6类香气化合物[7];低温在一定程度上提高了玛奎特葡萄浆果中具有强烈气味的芳樟醇和香叶醇等萜烯香气化合物的含量[8];葡萄园较冷的环境条件使得葡萄果实生长受到限制,从而抑制代谢物的积累和风味物质的形成;产地的昼夜温差是促进C6酯类和C6醇类合成的关键气象因子之一[9];昼夜温差与C6挥发物、3-异丁基-2-甲氧基吡嗪和(E)-b-达马斯基酮等挥发性化合物浓度之间存在显著相关关系[10],可能抑制巯基3-磺基己醇前体物的产生[11]。我国3个葡萄主产区中,新疆产地昼夜温差较大,而山东产地昼夜温差较小,宁夏葡萄产地昼夜温差介于新疆和山东产地之间。3个产地果实中挥发性香气物质组成有显著差异,山东产地果实中醛类和萜烯类物质种类和相对含量最高,新疆产地果实中酯类物质种类和相对含量最高,而宁夏产地果实中醇类物质种类和相对含量最高[12]。现有昼夜温差对果实香气影响的研究以通过对局部果实处理或者结合多气象因素为主,而各气象因素对葡萄的性能以及葡萄酒质量的影响存在复杂性,且多是各种因素相互作用,无法解释单一因素—昼夜温差对果实香气的影响机制[13],如在田间条件下,光照可促进温度升高,表明香味物质的合成受光照和温度的综合影响[14]。
本研究通过设施调控夜间温度改变昼夜温差,基于气相色谱法—质谱法联用技术(gas chromatographymass spectrometry,GC-MS)并进行转录组测序,研究昼夜温差对果实发育过程中香气物质积累的影响,以期为酿造者根据不同酿酒葡萄产区昼夜温差特征因地制宜地调整酿造工艺,打造风格特异的葡萄酒提供理论依据。
1 材料与方法
1.1 试验地概况与试验材料
试验于2021年7—9 月在宁夏吴忠青铜峡市西鸽酒庄葡萄园(北纬38.1°,东经105.8°)进行,该试验地位于贺兰山东麓偏南段,属中温带干旱气候区,平均海拔高度1 143 m,土壤类型为灰钙土,日照充足,年降水量在200 mm左右。
供试葡萄品种为25年生欧亚种酿酒葡萄(Vitis viniferaL.)赤霞珠(Cabernet Sauvignon),自根苗定植,南北行向栽培,株距为1.0 m,行距为2.5 m,整体采用单臂“厂”字形架式进行整形修剪,田间常规管理。
1.2 试验处理
在葡萄田间南北方向搭建3 间装配式钢架结构温室(图1),在每间温室内安装变频空调,于晚间20:00左右升起棉被打开空调,次日清晨6:00 降下棉被关闭空调,自转色前期至转色结束连续50 d 对温室进行夜间温度调控以改变昼夜温差,并利用ZDR-11 温度记录仪(杭州泽大仪器有限公司)和SS-H21400温室娃娃(杭州泽大仪器有限公司)监测温湿度等指标,查阅收集2015—2020年7—10 月气象资料(数据来源于宁夏气象局),计算该时期平均夜间温度,以此夜间温度为参考,设置4 组不同处理,即T-2.6℃(空调调控后夜间平均温度为17.0 ℃,温差为15.1 ℃)、T0.5℃(空调调控后夜间平均温度为20.1 ℃,温差为12.0 ℃)、T3.5℃(空调调控后夜间平均温度为23.1 ℃,温差为9.0 ℃)和CK(未经空调调控,夜间平均温度为19.6 ℃,温差为12.5 ℃)。夜间温度调控期间,温室内各项气象指标参数如表1所示。在最终采收期进行取样,每个处理各随机选取6 个果穗,带回实验室后随机剪取果粒,3 次重复,用液氮速冻后放入-80 ℃冰箱备用。
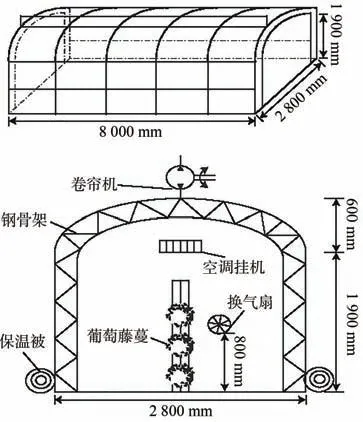
图1 夜间温室示意图[15]Fig.1 Nighttime greenhouse diagram[15]
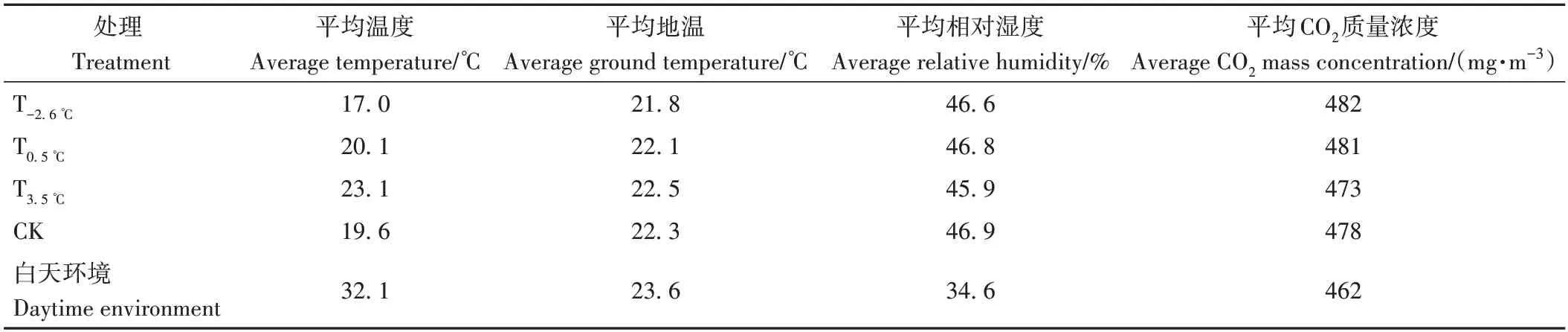
表1 调控夜间温度期间温室内气象参数Table 1 Meteorological parameters in greenhouse during night temperature regulation
1.3 试验方法
1.3.1 挥发性香气成分检测 参照孙磊等[16]的方法并加以改进。称取适量样品和内标(2-辛醇)置于顶空瓶中密封,置于80 ℃水浴中平衡30 min,用固相微萃取针萃取30 min,待萃取结束后,萃取针在进样口解吸5 min。色谱柱:HP-5MS(30 m×0.25 mm×0.25 μm);分流比:不分流;载气流速:1.0 mL·min-1;进样口温度:250 ℃;扫描方式:全扫描;离子源温度:230 ℃;四级杆温度:180 ℃;升温程序:起始温度为45 ℃,保持4 min,以6 ℃·min-1的速率升至130 ℃,保持6 min,再以10 ℃·min-1的速率升至230 ℃,保持8 min。采集到的质谱图利用美国国家标准与技术研究所(National Institute of Standards and Technology,NIST)谱库进行检索,鉴定样品中的挥发性成分,并利用内标法对各成分进行定量分析。
1.3.2 RNA 提取及转录组测序 委托上海美吉生物医药科技有限公司完成样品RNA提取和转录组测序,且每个样品3个生物学重复。样品RNA的提取通过Omini Plant RNA Kit(DNase I)试剂盒(康为世纪生物科技有限公司,泰州)完成,RNA检测合格后经反转录和PCR扩增得到cDNA文库,库检合格后,于Illumina平台测序。参考基因来源:Vitis_vinifera;参考基因组版本:12X;参考基因组来源:http://plants.ensembl.org/Vitis_vinifera/Info/Index。
1.3.3 差异基因的功能分析 基于表达量定量结果,采用DESeq2软件进行组间差异基因分析,获得两组间发生差异表达的基因[17],筛选阈值为:|log2FC|≥1,padjust<0.05,将筛选出的差异表达基因与基因本体(gene ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库进行比对,获得不同样品中的功能注释及相关代谢通路信息。
1.3.4 实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)验证分析 根据富集结果筛选出参与香气合成等代谢通路的差异表达基因,并从中选取9个基因进行qRT-PCR 验证分析(表2),内参基因为ACTIN1(GenBank:XP_008654957.1),引物序列通过Primer 平台设计并由上海生工生物工程有限公司合成(表2)。使用一步法反转录荧光定量试剂盒(B639277-0100,上海生工生物工程有限公司),反应程序为:95 °C预变性10 min,95 ℃变性30 s,60 ℃退火30 s,72 °C延伸40 s,40个循环。3次重复,采用2-ΔΔCT法计算基因相对表达量。

表2 qRT-PCR引物序列Table 2 Primer sequence of qRT-PCR
1.4 数据处理
利用SPSS 26.0软件进行数据统计分析,采用Duncan多重比较进行显著性方差分析,最低显著水平P<0.05;采用Origin Pro2021软件进行主成分分析和绘图。
2 结果与分析
2.1 不同昼夜温差对葡萄果实可溶性固形物、可溶性糖和可滴定酸含量的影响
由图2 可知,随着采样时间的延迟,不同昼夜温差处理下赤霞珠浆果可溶性固形物和可溶性糖含量整体呈现上升趋势,可滴定酸含量则呈下降趋势。采收时,T-2.6℃、T0.5℃、T3.5℃、CK 处理的可溶性固形物含量分别为26.1%、25.2%、24.8%和25.3%,可溶性糖含量分别为224.00、218.70、211.39 和221.12 mg·g-1,可滴定酸含量分别为8.45、6.45、5.15 和6.70 g·L-1。由此可见,昼夜温差与可溶性固形物、可溶性糖含量呈正相关,与可滴定酸含量呈负相关。
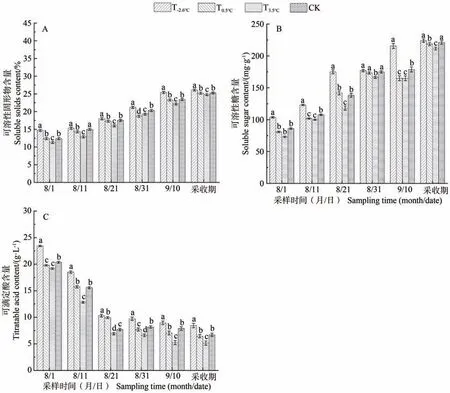
图2 不同昼夜温差对浆果可溶性物质的影响Fig.2 Effect of different diurnal temperature treatments on soluble substances in berries
2.2 不同昼夜温差对葡萄果实挥发性香气的影响
2.2.1 葡萄果实香气成分、种类比较 由表3 可知,不同昼夜温差处理下的赤霞珠葡萄果实香气总量差异显著,从高到低依次为T0.5℃>T-2.6℃>T3.5℃>CK,T-2.6℃、T0.5℃、T3.5℃的香气总量分别是CK的2.86、4.58、1.18倍;T-2.6℃、T0.5℃、T3.5℃、CK 的香气成分种类依次为24、24、29、14种。
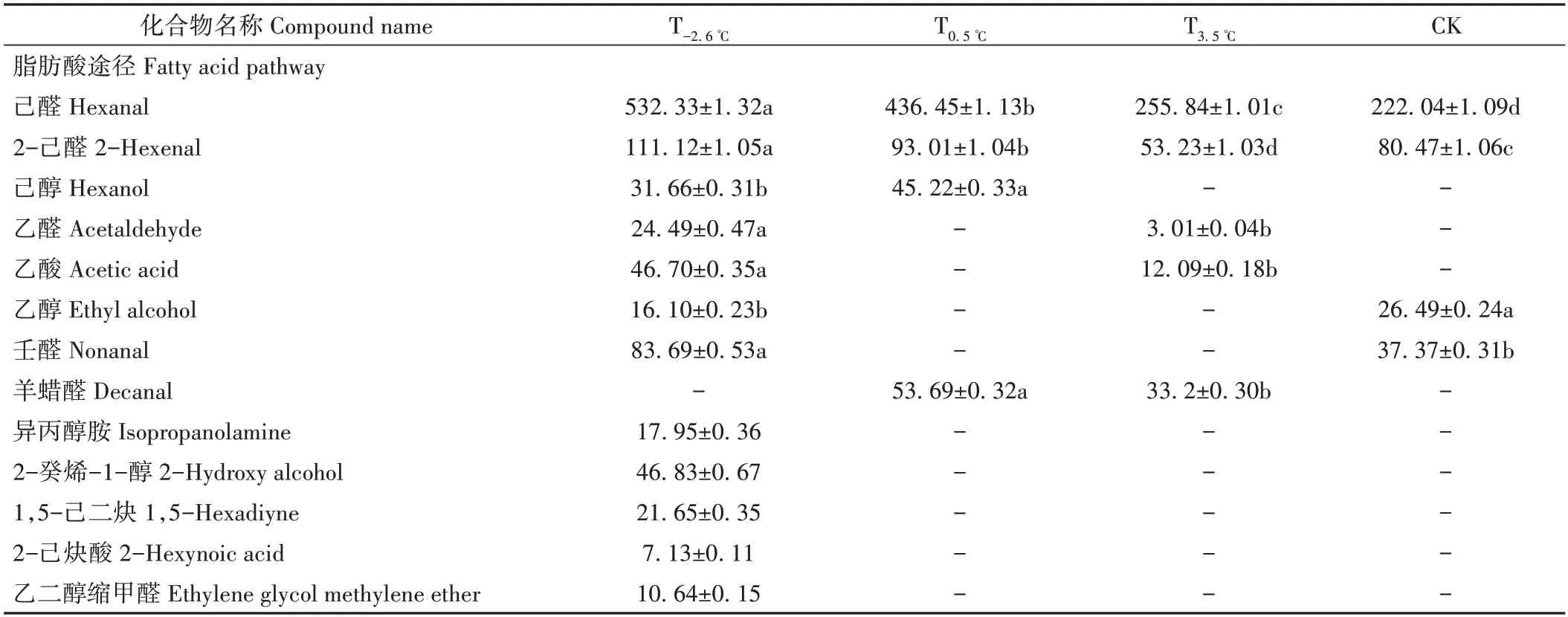
表3 不同昼夜温差处理下的葡萄果实香气成分种类和含量Table 3 Types and contents of aroma components of grape fruits under different diurnal temperature differences /(ng·kg-1)

表3(续)

表3(续)
T-2.6℃处理的葡萄果实在脂肪酸途径中的香气总含量高于其他处理的样品,分别是T0.5℃、T3.5℃、CK 处理的1.44、2.58、2.59 倍,其香气成分主要包括乙醛、己醛、2-己醛和乙酸等,是柑橘、苹果、番茄、紫罗兰等香气的主要贡献者。T0.5℃处理的葡萄果实在氨基酸途径中的香气总含量高于其他处理的样品,分别是T-2.6℃、T3.5℃、CK处理的3.04、6.84、10.38倍,其香气成分主要包括茴香脑、大茴香基丙酮、2-正庚呋喃、间茴香醛等,是风信子、水仙、苹果、草莓、树莓等香气的主要贡献者。与T-2.6℃和T0.5℃处理的葡萄相比,T3.5℃处理的样品在异戊二烯途径中的香气含量最高,分别为T-2.6℃、T0.5℃和CK处理样品的7.97、2.68和2.66倍,其香气成分主要包括大马士酮、香叶基丙酮、蘑菇醇等,主要贡献烤烟叶、香料、薄荷、埌香、药草等香气。
综合来看,T-2.6℃和T0.5℃处理提高了赤霞珠果实中的花香和果香,后续在酿酒过程中可提高葡萄酒一类香气的含量,在酿造干红的同时,也宜于酿造起泡桃红类葡萄酒;T3.5℃处理提高了果实中的烟草香,为后续酿造葡萄酒提供了更多的三类香气,即陈酿香,因此,在较小温差环境下生长的葡萄酿酒后更适宜储存较长时间以更好地形成陈酿香。
2.2.2 主成分分析 为了直观地展示不同昼夜温差对赤霞珠葡萄果实香气的影响情况,进行了主成分分析(图3)。其中,脂肪酸途径得到2 个主成分,解释了总方差的85.5%,第1主成分(PC1)和第2主成分(PC2)分别占总变异性的59.1%和26.4%,T-2.6℃和T0.5℃的点集中在第一、二、四象限,T3.5℃则集中在第三象限(图3-A);氨基酸途径得到2 个主成分,解释了总方差的75.9%,PC1 和PC2 分别占总变异性的46.2%和29.7%,T-2.6℃和T0.5℃的点集中在第一、二象限,T3.5℃则集中在第四象限(图3-C);异戊二烯途径得到2个主成分,解释了总方差的83.3%,PC1和PC2分别占总变异性的59.8%和23.5%,T-2.6℃和T0.5℃的点集中在第三、四象限,T3.5℃则集中在第一象限(图3-E),从视觉上说明不同昼夜温差对赤霞珠果实香气成分的影响存在较大差异。
由载荷图可以看出,脂肪酸途径中,T-2.6℃和T0.5℃葡萄果实积累了比较丰富的乙醛、己醛、乙醇、己醇、乙酸等,T3.5℃对庚醛等的积累较丰富(图3-B);氨基酸途径中,T-2.6℃和T0.5℃葡萄果实积累了较丰富的苯甘氨醇、苯乙醇、安息香醛、间茴香醛等,T3.5℃对苯甲醇、苯乙醛等的积累较丰富(图3-D);异戊二烯途径中,T-2.6℃和T0.5℃葡萄果实积累了较丰富的1,4-戊二烯等,T3.5℃积累了较丰富的大马士酮、香叶基丙酮、蘑菇醇、肉桂烯等(图3-F)。
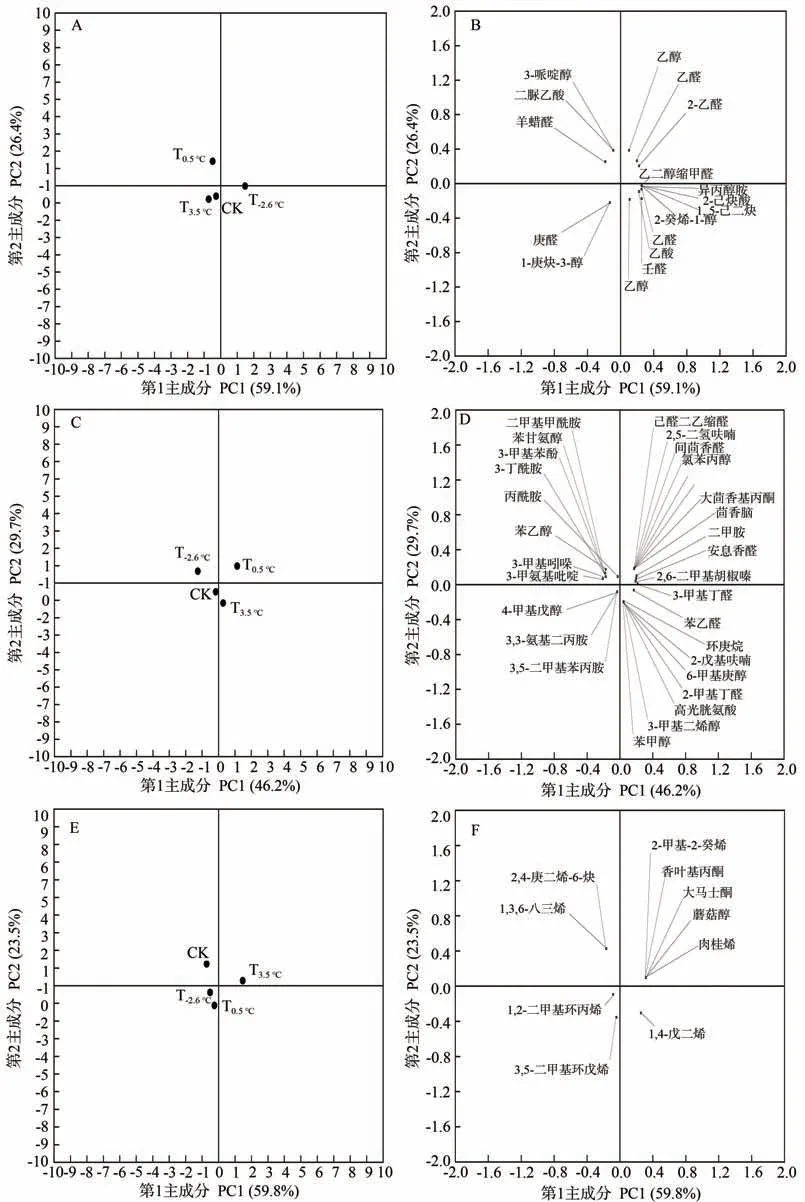
图3 葡萄果实香气成分含量主成分分析Fig.3 Principal component analysis of grape fruit aroma components
2.3 不同昼夜温差对葡萄果实香气相关基因表达的影响
2.3.1 RNA-seq 测序质量评估 由表4 可知,通过Illumina平台完成T-2.6℃、T0.5℃、T3.5℃和CK(分别包含3次重复在内)共12个样品的转录组分析,共获得80.03 Gb Clean Data,各样品Clean Data 均达到6.22 Gb 以上,Q30碱基百分比在92.51%以上,说明测序质量良好且可以进行后续比对分析;分别将各样品的Clean Reads 与指定的参考基因组进行序列比对,比对率从87.39%到90.14%不等,其中比对到多个位置的为3.41%~4.57%,比对到唯一位置的为83.68%~86.57%。
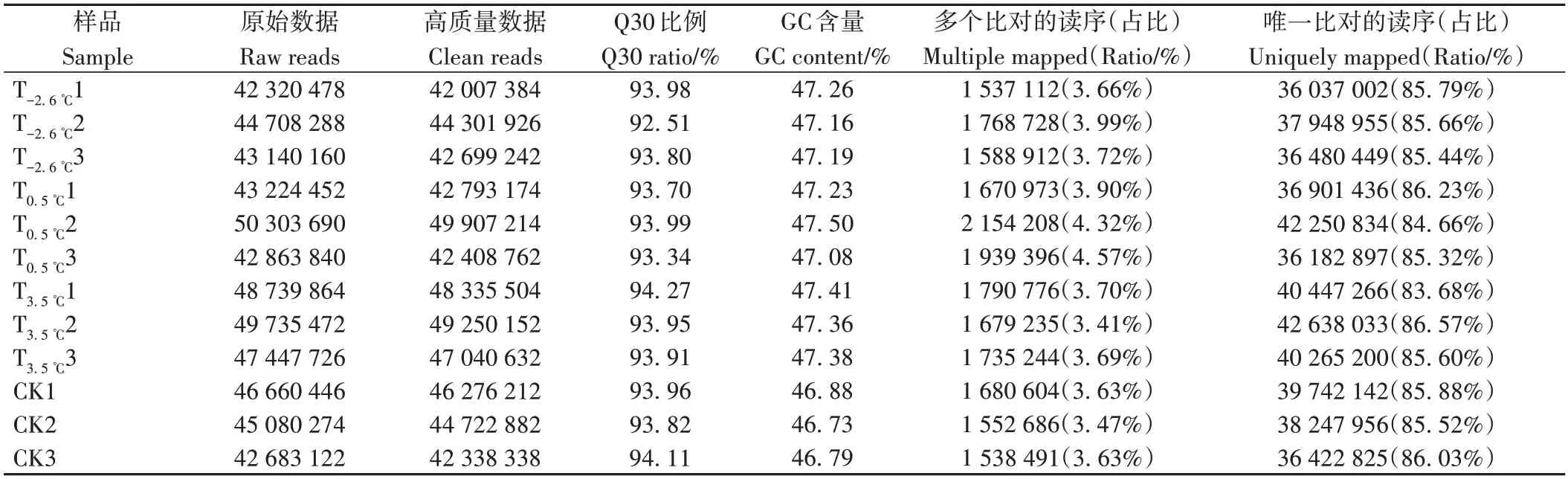
表4 不同昼夜温差处理葡萄果实转录组测序结果及比对统计Table 4 Transcriptome sequencing results and comparative statistics of grape fruit treated with different diurnal amplitude differences
2.3.2 差异表达基因的筛选及直系同源蛋白(clusters of orthologous groups,COG)分析 由图4-A 可知,不同昼夜温差处理下,共检测到5 111 个差异表达基因,其中,T-2.6℃与CK 之间差异表达基因为1 064 个,包括上调基因671个和下调基因393个;T0.5℃与CK之间差异表达基因为830 个,包括上调基因599 个和下调基因231个;T3.5℃与CK之间差异表达基因为3 107个,包括上调基因1 911个和下调基因1 196个。由图4-B可知,不同昼夜温差处理共有差异表达基因132 个,T-2.6℃特异表达差异基因有488 个,T0.5℃特异表达差异基因有263 个,T3.5℃特异表达差异基因有2 123个。
由图4-C 可知,比对差异表达基因与COG 数据库后,在COG 的21 个分类中,T-2.6℃、T0.5℃、T3.5℃注释到S(function unknown,未知功能)(586、1 622、456)的基因数最多,较多为K(transcription,转录相关基因)(53、166、39)、O(posttranslational modification,protein turnover,chaperones,翻译后修饰,蛋白质折叠和分子伴侣)(50、138、55)等功能条目,而注释到A(RNA processing and modification,RNA的加工和修饰)(1、11、0)、F(nucleotide transport and metabolism,核苷酸的转运和代谢)(7、15、3)等功能条目的差异表达基因数较少。
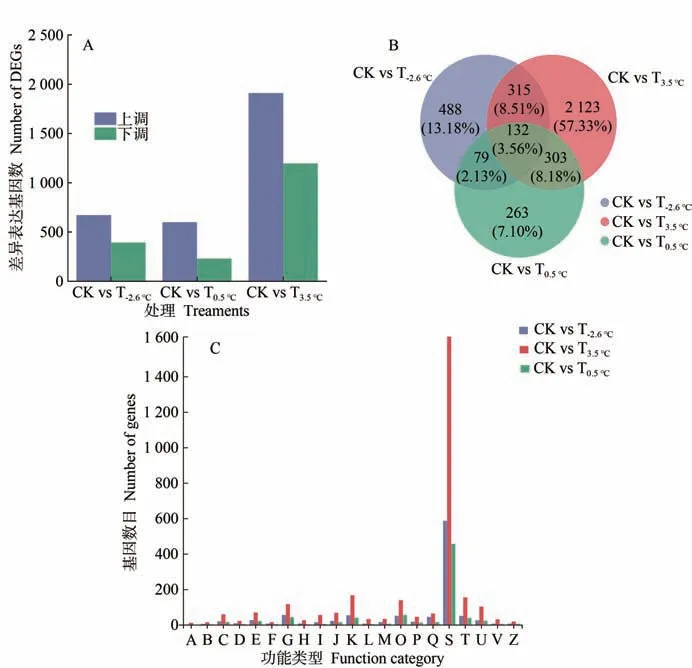
图4 差异表达基因的筛选及COG分析Fig.4 Screening of differentially expressed genes and COG analysis
2.3.3 差异表达基因GO和KEGG 富集分析 对差异表达基因进行GO 注释分析(图5)可知,在细胞成分、分子功能、生物过程三大条目的差异表达基因存在较大差异,图5 仅展示丰度前20 的GO 富集条目,不同昼夜温差处理与CK 之间富集到的条目数一致,在细胞成分中主要富集到的条目有代谢过程、细胞过程、生物调控等;在分子功能中主要富集到的条目有结合、催化活性、转运活性等;在生物过程中主要富集到的条目有细胞部分、膜组件、薄膜、细胞器、含蛋白质复合物等。
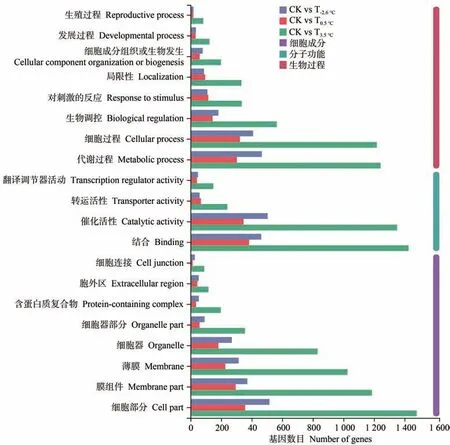
图5 差异表达基因GO分析Fig.5 GO analysis of differentially expressed genes
对处理间的差异表达基因进行KEGG 富集分析,结果表明(图6),注释到KEGG数据库的差异表达基因共有1 252 个,代谢通路119 个,图6 仅展示丰度前10的KEGG 富集条目,在CK vs T-2.6℃中,差异表达基因在植物激素信号转导、硫代谢、氰基氨基酸代谢、油菜素类固醇生物合成、光合作用-天线蛋白、光合作用、芪类、二芳基庚烷类和姜辣素生物合成、半乳糖代谢、柠檬烯和蒎烯降解、昼夜节律-植物途径中明显富集;在CK vs T0.5℃中,差异表达基因在淀粉和蔗糖代谢、色氨酸代谢、半乳糖代谢、柠檬烯和蒎烯降解、氮代谢、苯丙烷生物合成、黄酮类生物合成、昼夜节律-植物、亚油酸代谢途径中明显富集;在CK vs T3.5℃中,差异表达基因在芪类二芳基庚烷类和姜辣素生物合成、类胡萝卜素生物合成、内质网中的蛋白质加工、类固醇生物合成、酪氨酸代谢、色氨酸代谢、柠檬烯和蒎烯降解、氮代谢、苯丙烷生物合成、昼夜节律-植物途径中明显富集。其中,关于香气的代谢合成,差异表达基因主要富集在亚油酸、苯丙氨酸、酪氨酸代谢、柠檬烯和蒎烯降解途径,据此推测,上述几种代谢途径在各处理香气形成中起着重要作用,因此,通过KEGG 富集对香气合成途径相关的亚油酸、苯丙氨酸、酪氨酸代谢、柠檬烯和蒎烯降解途径中的相关基因进行进一步分析。
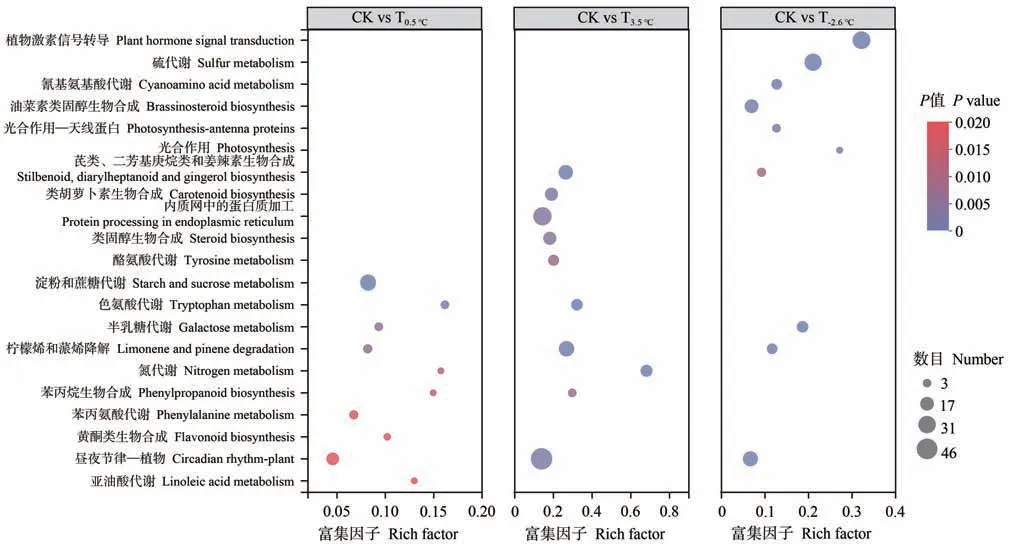
图6 差异基因KEGG富集散点图Fig.6 Enrichment scatter plot of differential gene KEGG
2.3.4 香气物质相关候选基因的筛选 亚油酸代谢途径属于脂肪酸途径,由图7 可知,在此代谢通路上共注释到8个差异表达基因(differentially expressed genes,DEGs),其中包括5个脂氧合酶(lipoxygenase,LOX,VIT_14s0128g00780、VIT_09s0002g01080、VIT_00s0265g00 170、VIT_13s0064g01480 和VIT_06s0004g01510),2 个乙醇脱氢酶(alcohol dehydrogenase,ADH,VIT_07s0005g04 610和VIT_18s0001g15410),1个丁酰辅酶A脱氢酶(butylcoa dehydrogenase,HBD,VIT_19s0014g02370)。LOX 和HBD相关基因在T-2.6℃和CK处理下大多下调表达,在T0.5℃和T3.5℃处理多上调表达;ADH相关基因在T-2.6℃和CK处理下大多上调表达,在T0.5℃和T3.5℃处理下多下调表达。
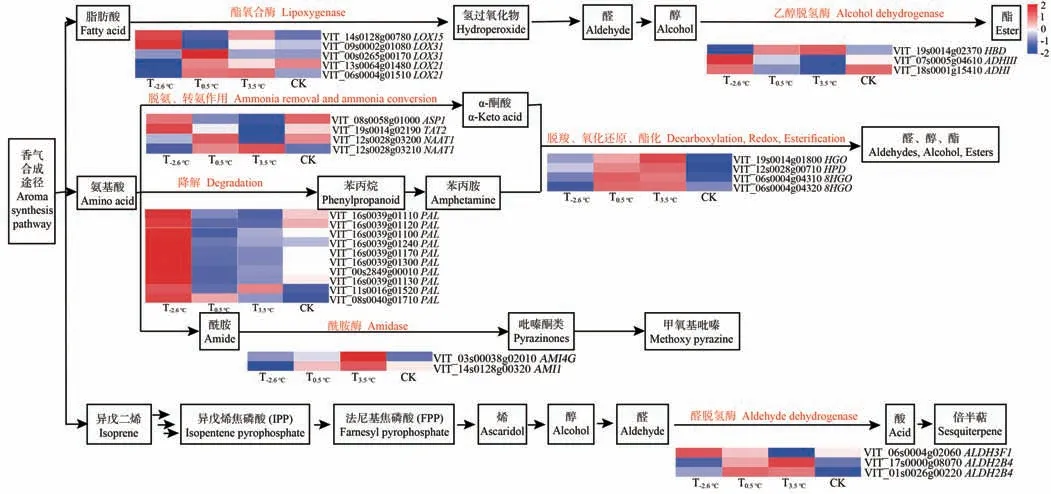
图7 香气物质代谢合成途径相关基因Fig.7 Genes related to aroma substance metabolic synthesis pathway
苯丙氨酸和络氨酸代谢途径属于氨基酸途径,在这两个代谢通路上共注释到20个DEGs,主要包括1个天冬氨酸氨基转移酶(aspartate aminotransferase,ASP,VIT_08s0058g01000)、1个转氨酶(transaminase,TAT,VIT_19s0014g02190)、2 个烟酰胺氨基转移酶(niacinamide aminotransferase,NAAT,VIT_12s0028g03200 和VIT_12s0028g03210)、10 个苯丙氨酸裂解酶(phenylalanine lyase,PAL)、2个酰胺酶(Amidase,AMI,VIT_03s0038g02010和VIT_14s0128g00320)、1个双加氧酶(dioxygenase,HGO,VIT_19s0014g01800)、1 个丙酮酸双加氧酶(pyruvate dioxygenase,HPD,VIT_12s0028g00710)、2个香叶醇氧化还原酶(geraniol oxidoreductase,8HGO,VIT_06s0004g04310和VIT_06s0004g04320)。T-2.6℃和CK 处理促进了ASP、TAT、PAL多数相关基因的上调表达;NAAT、AMI、HGO、HPD、8HGO多数相关基因在T0.5℃和T3.5℃处理上调表达。
柠檬烯和蒎烯途径属于异戊二烯途径,在此代谢通路上共注释到3 个DEGs,主要为醛脱氢酶(aldehyde dehydrogenase,ALDH,VIT_06s0004g02060,VIT_17s0000g 08070和VIT_01s0026g00220),T0.5℃和T3.5℃处理促进了ALDH 相关基因的上调表达;而T-2.6℃和CK 处理则降低了ALDH相关基因的转录水平。
综上可知,较大的昼夜温差促进了ADH、ASP、TAT、PAL 相关基因的上调表达,较小的昼夜温差促进了LOX、HBD、NAAT、AMI、HGO、HPD、8HGO、ALDH相关基因的上调表达。
2.3.5 qRT-PCR 验证 为检验转录组测序结果的真实性,本研究从香气代谢合成途径相关基因中随机挑选了9个候选基因LOX(VIT_06s0004g01510)、HBD(VIT_19s0014g02370)、HPD(VIT_12s0028g0071)、HGO(VIT_19s0014g01800)、ADHIII(VIT_07s0005g04610)、HPPD(VIT_12s0028g00170)、ADHI(VIT_18s0001g15410)、ALDH2B4(VIT_17s0000g08070)、ALDH2B4(VIT_01s0026g00220)进行qRT-PCR验证(图8)。将qRT-PCR验证结果与RNAseq的数据进行相关性分析,可知两者的相关系数较高且表达趋势基本相同,表明RNA-seq的分析结果可靠。
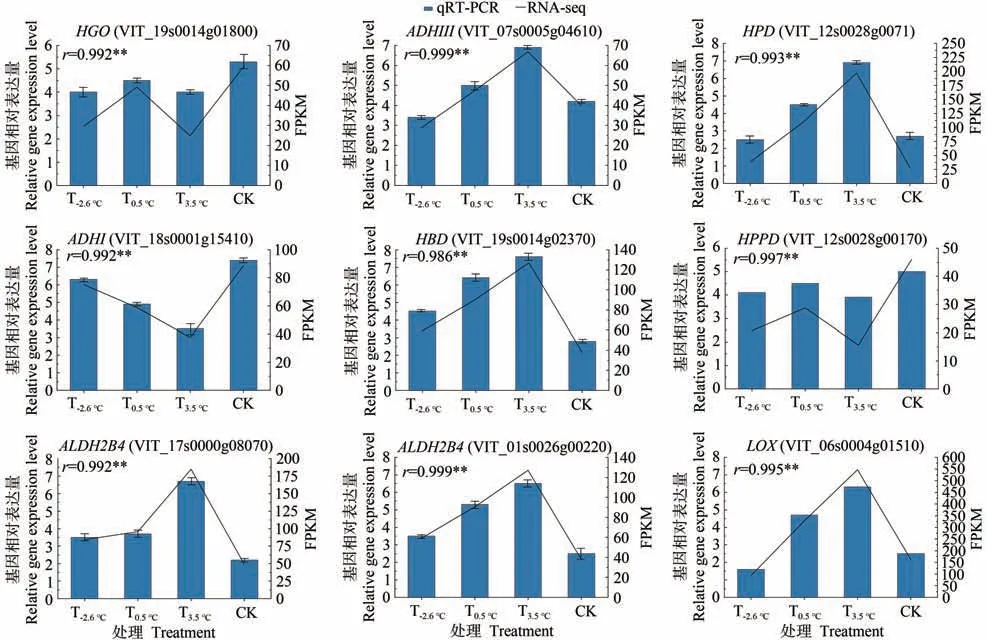
图8 部分基因的qRT-PCR验证Fig.8 qRT-PCR validation of some genes
3 讨论
刘玉兰[18]研究发现果实着色期气温平均日较差对葡萄果实总糖含量有重要影响,其中,最低气温影响最大,而本研究发现降低夜间温度增大昼夜温差,可溶性固形物和可溶性糖含量呈上升趋势,造成这一现象的原因可能是由于夜间低温减弱了呼吸作用,使得浆果内有机物质的消耗减少,从而促进含糖量等的升高[18]。
香气是葡萄的重要品质特征之一,是葡萄酒消费者、葡萄种植者和葡萄酒酿造者共同关注的焦点。赤霞珠作为在中国各大葡萄产区均有种植的酿酒品种之一,对于其品质尤其是香气品质的提升也受到广泛关注。一般认为,冷凉环境有利于香气物质的产生,短暂的高温会加速果实成熟,提高葡萄酿酒后的品质,然而长期高温或强光照会降低吡嗪类物质的含量,导致香气品质改变。炎热的气候会促进雷司令葡萄果实和葡萄酒中产生一种带有煤油味的物质—1,1,6-三甲基-1,2-二氢萘(1,1,6-trimethyl-1,2-dihydronaphthalene,TDN),但气候过冷会导致果实香气物质含量下降,青草味加强,因此,温度过高或过低都不利于香气物质的积累[19]。本研究发现,降低夜间温度增大昼夜温差提高了赤霞珠葡萄果实中醛类、酯类、醇类、萜烯类挥发性香气物质的含量,这可能是由于冷凉环境有利于香气物质的形成,如生长季后期的低温环境显著提高了一年两收栽培的夏黑葡萄冬果C6醛类香气物质的含量,主要特征香气为花果香[20],同时,冷凉环境在一定程度上也会使萜烯类含量上升[21];升高夜间温度减小昼夜温差可促使赤霞珠葡萄果实中酮类和酸类挥发性香气物质的积累,是由于适度的高温会加速果实成熟,从而提高葡萄果实部分香气物质的含量[22]。
脂氧合酶(lipoxygenase,LOX)和乙醇脱氢酶(alcohol dehydrogenase,ADH)是脂肪酸途径合成挥发性香气的关键酶[23],LOX 与果实成熟过程中香气物质释放的关系密切,如LOX 活性与不饱和脂肪酸及脂肪酸来源香气成分的积累显著相关[24];挥发性酯类化合物是由醇酰基转移酶催化醇和羧酸酯化生成的,脂肪酸代谢途径为酯类化合物的生物合成提供醇等底物,LOX 在巨峰和87-1 葡萄浆果中较高水平的表达提高了挥发性酯类化合物的含量[25];雷司令、维多利亚和优株玫瑰葡萄冬果成熟过程中LOX 的高转录水平对冬果中C6醛类物质的高积累起主要作用[26],在葡萄果实发育过程中,ADH 的严密调控可能为改变葡萄和葡萄酒香气提供契机,如利用植物育种或分子遗传方法调控葡萄浆果中的ADH 水平以促进醇类(果香)的产生[27]。本研究发现,降低夜间温度增加昼夜温差降低了LOX 相关基因的转录水平,促进了ADH 相关基因的上调表达,这与刘孟龙[28]的研究结果基本一致。苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)是苯丙烷途径的关键酶,催化苯丙烷类代谢第一步反应,在植物果实着色、次生代谢以及发育过程中起重要作用[29]。大量PAL 在小果粒葡萄果实中显著上调使得二苯乙烯在果实开始成熟时大量积累[30]。前人研究发现,适当降低温度可提升PAL 活性,调控PAL相关基因上调表达[31-32],这与本研究中降低夜间温度增大昼夜温差可升高PAL 相关基因转录水平的结果基本一致。此外,本研究还发现香气代谢合成途径中LOX、ADH 相关基因与葡萄果实香气物质存在密切关系,较大的昼夜温差条件下,乙醛、己醛等物质含量增加,同时LOX 相关基因下调表达而ADH 相关基因上调表达。并且,PAL 相关基因转录水平的提高促进了2-己烯醛和乙酸等香气物质的积累,为葡萄贡献了柑橘、苹果、番茄、紫罗兰等香气。
4 结论
昼夜温差与可溶性固形物、可溶性糖含量呈正相关,与可滴定酸含量呈负相关,较大的昼夜温差可为葡萄酒提供更多品种香,并促进ADH、ASP、TAT、PAL 相关基因的上调表达;较小的昼夜温差可为葡萄酒提供更多陈酿香,并促进LOX、HBD、NAAT、AMI、HGO、HPD、8HGO、ALDH相关基因的上调表达。
猜你喜欢
小猕猴智力画刊(2023年11期)2023-12-28 09:13:12
阅读(低年级)(2020年11期)2020-12-28 02:26:35
女报(2020年10期)2020-11-23 01:42:42
河南畜牧兽医(2020年23期)2020-01-10 03:19:52
小资CHIC!ELEGANCE(2019年16期)2019-06-15 08:43:22
儿童故事画报·发现号趣味百科(2019年4期)2019-05-10 07:09:38
电子制作(2018年23期)2018-12-26 01:01:20
Coco薇(2017年8期)2017-08-03 20:46:09
小青蛙报(2016年2期)2016-10-15 05:05:56
Coco薇(2016年8期)2016-10-09 02:08:33
