
紫茉莉β-谷甾醇对镉处理的浓度响应及其化感效应
2023-03-07 04:46张文亭王宏镔王海娟郭思宇
生态与农村环境学报 2023年2期
张文亭,王宏镔,王海娟,彭 瑜,郭思宇
(昆明理工大学环境科学与工程学院/ 云南省土壤固碳与污染控制重点实验室,云南 昆明 650500)
镉(Cd)因具有重要的工业用途,对其需求日益增加,Cd 污染成为一个严重的环境问题。一方面,排入土壤中的 Cd 具有高移动性、毒性和污染广泛性等特征,作为环境中各类污染物质最终归属地的土壤成为一个主要污染场所[1];另一方面,Cd 易通过食物链在生物体内积累,对动、植物及人体健康产生巨大风险[2]。作为农业大国,Cd 污染土壤的修复在我国尤显重要且迫切。
作为一种从土壤中去除有毒重金属和其他污染物的有效、廉价和环境友好的技术,植物修复(phytoremediation)技术常用于转移或稳定污染土壤中的有毒金属和其他污染物[3]。该技术的关键是找到合适的修复植物,并且这类植物应具有分布广泛、地上部生物量高、生物富集系数高、生命周期短、繁殖速率高等特点[4]。因此,在所发现的超富集植物种质资源有限、生物量低、生长缓慢等现实条件下,寻求利用外来入侵植物(alien invasive plant,AIP)修复重金属污染土壤是一个值得考虑的思路。
云南是有名的“植物王国”,根据《云南省外来入侵物种名录(2019版)》,云南省共查明外来入侵植物321种和4个变种[5]。若AIP具有较强的重金属富集能力,可望用于重金属污染土壤的植物提取(phytoextraction)修复;若其重金属富集能力不强,则可以利用这类植物较强的抗逆性,在广大重金属矿区将其作为先锋植物,用于重金属污染土壤的植物固定(phytostabilization)修复,增加矿区植被覆盖率,防止水土流失造成重金属扩散,从而化害为利[6]。近来有研究发现,尽管外来入侵植物紫茉莉(Mirabilisjalapa)在野外生长条件下未表现出 Cd 超富集植物特征[7],但在盆栽条件下已接近 Cd 超富集植物的标准[8-9]。三叶鬼针草(Bidenspilosa)[10]和假苍耳(Ivaxanthifolia)[11]等 AIP 都具有较强的富集Cd的能力,但紫茉莉因花形美观、花色多样且在我国各地均有栽培,具有其他植物不可比拟的公众可接受性。此外,紫茉莉对其他重金属(如铅和铬)也具有较强的富集能力,即具有富集多种重金属的功能[12-13]。LIU等[14]综述了国内外运用观赏植物修复污染土壤的研究进展,认为作为一种观赏性修复植物,紫茉莉在修复 Cd 污染土壤方面具有潜在用途。
化感作用是植物种间关系的一种重要类型,是由植物通过释放化感物质到环境中而对另一种植物产生积极或消极作用的现象[15]。有研究认为,外来入侵种具有化感作用,紫茉莉植株体内已被检测到化感物质甾醇(sterol)[16-19]。甾醇是以环戊烷全氢菲(甾核)为骨架,大多以C—5位双键、C—3位羟基作为重要活性基团的醇类化合物,以及以此为基础形成的各种衍生物。因此,各甾醇在结构上非常相似,主要区别在于侧链基团不同[20]。植物甾醇种类繁多,紫茉莉中甾醇主要有β-谷甾醇〔β-sitosterol,(24R)-24-ethyl-cholest-5-en-3β-ol〕和豆甾醇(stigmasterol,22-dien-3β-ol)[21-22],紫茉莉β-谷甾醇是自然界中以游离态或结合态存在的一类甾醇类物质,具有分子量较高、含有极性基团、沸点较高、不易气化等特点[20]。β-谷甾醇的合成主要有3个阶段:首先乙酰辅酶 A(acety-l CoA)分子在 6C 一系列酶作用下形成 5C 的异戊烯焦磷酸(isopentenyl pyrophosphate,IPP)分子,然后 IPP 分子聚合形成 C30 的鲨烯(squalene),最后形成植物体内的环阿屯醇,再经过一系列酶的作用最终完成β-谷甾醇合成[20]。紫茉莉β-谷甾醇结构与动物胆甾醇结构很相似,因而具有许多与胆甾醇相同的生理功能,如抗癌、延缓衰老、预防疾病等。Cd污染下,β-谷甾醇可在紫茉莉抵抗环境不利因素过程中发挥重要作用,如稳定细胞膜、具有较强抗氧化活性等[23]。但有研究表明,甾醇形成修饰过程复杂,若紫茉莉的生长环境正常,则β-谷甾醇的生物合成也是正常的;若受到环境胁迫,甾醇作为植物在生物逆境胁迫过程中重要的化学物质之一,能迅速调节自身合成机制。而其合成过程有可能产生具有生物活性的物质,也可能产生有毒性、刺激性或激素活性的物质[20],这些物质均有可能在抗逆过程中释放到环境,此时产生甾醇的植物体本身获利,释放到环境中的甾醇则对周围植物正常的生理活动、代谢过程、植物内源激素平衡维持构成巨大威胁,从而导致周围植物无法正常生长[24-26]。因此,若运用外来入侵植物紫茉莉修复 Cd 污染农田土壤,其可能会通过释放化感物质抑制作物的生长发育,并促进自身扩张成为本地生境的单优种群[27]。然而,这些研究大多在无重金属污染的情况下进行,当重金属污染对作物和紫茉莉均构成选择压力时,Cd 处理下紫茉莉是否会产生或分泌甾醇类物质,甾醇类物质对作物产生的化感潜力会发生怎样的变化,目前还鲜见报道。
因此,若将紫茉莉运用于重金属污染土壤的植物修复实践,重金属污染下其是否仍会产生化感作用并排挤农作物等生态风险问题不容忽视。玉米、大豆是当前全国种业研究的重点对象,云南又是西南地区黄豆和玉米的主要产地[28],超富集植物研究常将玉米或其他农作物与其进行间作[29-31],化感研究也常用玉米和黄豆作为受试植物[32-33]。以紫茉莉、黄豆、玉米作为供试植物,采用室内水培实验,探究 Cd 处理下紫茉莉根系产生的β-谷甾醇对黄豆和玉米种子萌发和幼苗生长的影响。笔者研究的目的是:(1)探明 Cd 处理下紫茉莉体内和根系分泌物中主要的甾醇类物质——β-谷甾醇的含量;(2)进一步确认β-谷甾醇对2种作物化感效应的强弱。该研究可为深入探究 Cd 胁迫下紫茉莉植株与作物的种间关系、利用外来入侵植物进行土壤环境修复实践提供科学依据。
1 材料与方法
1.1 供试植物
紫茉莉种子经φ=75% 乙醇表面消毒后,进行萌发生长实验。紫茉莉在温室的清洁土壤中自然光照条件下培养45 d后,选择大小一致的幼苗进行水培实验。紫茉莉种子购自江苏绿友种业有限公司,玉米和黄豆种子均购于云南京滇种业有限公司。
1.2 紫茉莉培养及根系分泌物收集
植物培养:用容积为 2.5 L 的塑料容器设置不同浓度含Cd 营养液(0、1、3、5 mg·L-1)进行培养,Hoagland 营养液参照 ZHANG 等[34]介绍的方法配制,Cd 以 CdCl2·2.5H2O 形式添加,Cd2+浓度以纯Cd 计。紫茉莉移入水培装置后,每个容器放置 6 株,且保证植株的大小和鲜重较为一致,植物放置在温室自然光照条件下培养。植物生长期间用氧气泵持续充氧,每隔 3 d 更换1次培养液,每处理均设置 3 个重复,培养14 d后收获。
根系分泌物收集:用章家恩[35]介绍的方法收集紫茉莉根系分泌物,根据Cd处理浓度依次收集,即紫茉莉水培14 d后,将其取出,用EDTA·Na2和去离子水先后分别冲洗根系数次以去除根表面附着的Cd,再将根放入 0.2 mol·L-1CaCl2溶液2 h以抑制细菌生长。随后将植株(只有根浸没在去离子水中)放入盛有 500 mL 灭菌去离子水的玻璃瓶中(玻璃瓶外包有锡箔纸避光),置入光照培养箱 6 h后将植株取出,收集的水溶液即为相应 Cd 处理下的紫茉莉根系分泌物。再将根系分泌物收集液经布氏漏斗抽滤、减压浓缩,连续收集的根系分泌物定容至 500 mL,一部分放入 4 ℃ 冰箱备用,另一部分进行作物种子萌发和幼苗生长的生物学测试。
1.3 甾醇处理液的制备
为进一步明确 Cd 处理下紫茉莉根系分泌物中β-谷甾醇的化感作用,并排除其他化感物质对作物的干扰,采用外源添加β-谷甾醇进行作物的化感作用实验。Cd 处理质量浓度为 3 mg·L-1,准确称取25 mgβ-谷甾醇,先用 2 mL 甲醇溶解后再用去离子水稀释至 250 mL,用RE 52-99 型旋转蒸发器(上海亚荣生化仪器厂)于 50 ℃ 条件下旋转蒸发,待溶液体积减少约 25% 后加100 mL 去离子水,重复操作 5 次,以去除溶液中的甲醇,然后用去离子水定容至 500 mL。为保证外加β-谷甾醇浓度接近紫茉莉根系分泌物的浓度,用去离子水分别稀释至 0、0.125、0.5、1 和 2 mg·L-1作为外加β-谷甾醇处理液,进行β-谷甾醇对作物的影响实验。
1.4 样品处理及测定
1.4.1样品前处理
紫茉莉根系分泌物收集完成后,再次先后用自来水、去离子水、EDTA·Na2溶液、去离子水分别清洗紫茉莉植株数次。测量植物高度及根长,将植株分为根、茎、叶3个部分,用滤纸吸干植物表面水分并称量鲜重后,置于烘箱 105 ℃ 杀青30 min,然后在 70 ℃ 下烘干至恒重,磨碎后得到待测样品,室温下保存。紫茉莉β-谷甾醇采用超声波辅助法[36]进行提取,结晶法进行提纯[37],至少结晶 3 次,以保证得到纯度更高的β-谷甾醇,最终得到待测液。提取、结晶操作流程为:紫茉莉样品→甲醇提取剂超声辅助提取→过滤→滤液 50 ℃ 旋转蒸发浓缩→ 50 ℃ 蒸干,加 1 mL 甲醇溶解→β-谷甾醇粗制品→乙醇重结晶 3 次→旋转蒸发仪去除乙醇→β-谷甾醇精制品→超高效液相色谱分析。
1.4.2种子萌发及幼苗生长实验
参照文献[38]介绍的方法进行化感作用实验。首先进行不同 Cd 处理浓度下紫茉莉根系分泌物对黄豆和玉米的化感效应实验。其次,由于收集到的紫茉莉根系分泌物是混合物,为进一步研究Cd处理下紫茉莉根系分泌物中β-谷甾醇的化感效应大小,外源添加 0、0.125、0.5、1 和 2 mg·L-1β-谷甾醇溶液,进行 2 种作物的化感效应实验。
(1)种子萌发实验
选取饱满、大小一致的黄豆和玉米种子,用75%酒精浸泡1 min,φ=1% NaClO 溶液消毒 3 min,然后用去离子水清洗干净,吹干备用。取洗净的种子 50 粒放入铺有双层滤纸的培养皿(直径90 mm)中,分别加入不同 Cd 处理浓度下的紫茉莉根系分泌物或不同浓度β-谷甾醇溶液20 mL,每个处理重复3次。种子于恒温培养箱中25 ℃ 暗培养 7 d, 3 d时在每个培养皿中补加 10 mL紫茉莉根系分泌物或β-谷甾醇。每隔 12 h 统计1次种子发芽数(以芽突破种皮为发芽标准),记录数据。
(2)幼苗生长实验
取胚根刚突破种皮且较为一致的种子 25 粒,均匀摆放在铺有滤纸的培养皿中,加 20 mL不同Cd处理浓度下的紫茉莉根系分泌物(对照为蒸馏水)或不同浓度β-谷甾醇处理液(对照为不加β-谷甾醇处理),每处理设置 3 个重复,每隔 3 d在每个培养皿中补加紫茉莉根系分泌物或β-谷甾醇溶液 10 mL。培养 15 d 时,在每个培养皿随机选取 10 粒种子,测定其胚根、胚芽长度及鲜重,每处理计算 3 次重复的平均值。
1.4.3样品测定
植物Cd含量测定:植物样品采用 HNO3-H2O2加热消化,用火焰原子吸收光谱仪(AA240FS,美国 Varian公司)测定植株各部位 Cd 含量。
紫茉莉样品中甾醇的分析测定:对紫茉莉提取结晶所得供试液进行测定,并用豆甾醇、β-谷甾醇标准品进行对照分析。采用超高效液相色谱法(Agilent 1200 型液相色谱仪)进行甾醇含量分析测定,色谱柱:Nova-Pak C18Column (150 mm×4.6 mm×5 μm);流动相:V(甲醇)∶V(水)=70∶30;检测波长 210 nm;柱温 25 ℃;流速 1.0 mL·min-1;进样量 20 μL。甾醇组分鉴定选用保留时间法定性和峰面积外标法定量[39]。
对紫茉莉甾醇提取液样品进行定性与定量分析,得到甾醇提取液的液相色谱图,经与标样对比,色谱定性结果如图1 所示。图1中,1为豆甾醇,2 为β-谷甾醇,3、4 为未知峰,并且 2.622 min 处为β-谷甾醇的检测峰。设x为β-谷甾醇的物质的量浓度,y为相应的峰面积,得到相应直线的回归方程为y=54 900x-35 300,r= 0.999 8,然后根据该方程计算紫茉莉β-谷甾醇含量。
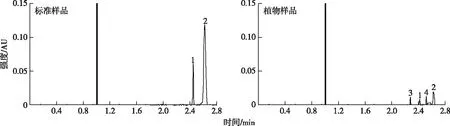
图1 紫茉莉甾醇色谱
种子发芽率=测试种子发芽数/测试种子总数×100%;发芽势=发芽初期 3 d的发芽种子数/测试种子总数×100%。
采用WILLIAMSON 等[40]提出的化感响应指数(allelopathic response index,RI)表征化感效应强弱,计算公式为
IR=(T/C-1)×100%。
(1)
式(1)中,IR为化感响应指数;C为对照值;T为处理值。IR>0说明化感效应表现为促进作用,IR<0说明为抑制作用,IR=0说明无影响,IR绝对值表征化感作用强度大小。综合效应(synthesis effect,SE)是供体对同一受体所有测试项目的化感作用抑制(或促进)百分率的算术平均值[41]。
生理生化指标测定:作物幼苗生长 15 d 后进行幼苗叶片生理生化指标测定,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光化还原法[42],过氧化物酶(POD)活性测定采用愈创木酚法[35],过氧化氢酶(CAT)活性测定采用过氧化氢还原法[43]。SOD活性以抑制NBT光化还原50%作为1个酶活性单位(U),POD活性以每分钟O.D.470(470 nm的光密度)增加0.01为1个酶活性单位,CAT活性以每分钟O.D.240减少0.1的酶量为1个酶活性单位。丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)比色法[43],叶绿素含量测定采用乙醇和丙酮等量浸提比色法[44]。
1.5 数据处理
数据先用 Microsoft Excel 2010 进行简单处理,用平均值±标准差(SD)表示。各处理之间的显著性差异程度运用 SPSS 20.0 软件进行方差分析,显著性差异水平取 0.05,极显著差异水平取 0.01。用 Origin 2018 软件绘图。
2 结果与分析
2.1 不同 Cd 处理浓度对紫茉莉生长、Cd 吸收和富集影响
紫茉莉在水培条件下培养 14 d时,不同 Cd 处理浓度之间其株高、根长、生物量无显著差异(P>0.05)。除根外,紫茉莉茎和叶片中的 Cd 含量随着 Cd 处理浓度增加而显著增加(P<0.05)。在相同 Cd 处理浓度下,除对照外,同一植株中紫茉莉根部吸收的 Cd 含量显著高于茎和叶片(P<0.05),但是茎和叶片之间 Cd 含量无显著差异(P>0.05,表 1)。
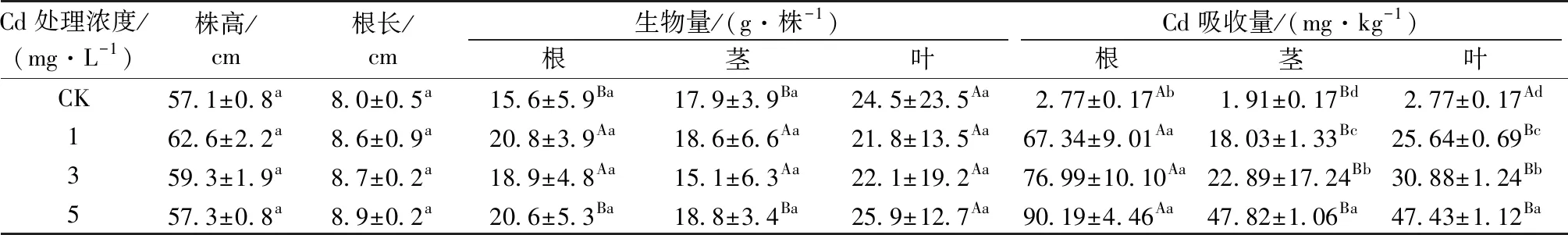
表1 不同 Cd 处理浓度对紫茉莉生长和Cd 含量的影响
2.2 不同 Cd 处理浓度下紫茉莉根系分泌物对作物的化感效应
不同 Cd 处理浓度下收集的紫茉莉根系分泌物对 2 种作物的化感效应不同(表 2)。随着 Cd处理浓度升高,紫茉莉根系分泌物使黄豆的发芽势、发芽率、根长均呈逐渐降低趋势,苗高仅在3 mg·L-1Cd 处理下显著降低,鲜重则无显著变化(P>0.05);玉米的发芽势、鲜重也均呈下降趋势。3~5 mg·L-1Cd 处理下的紫茉莉根系分泌物使玉米根长显著增加(P<0.05),说明紫茉莉根系分泌物对黄豆和玉米的影响不同(表 2)。
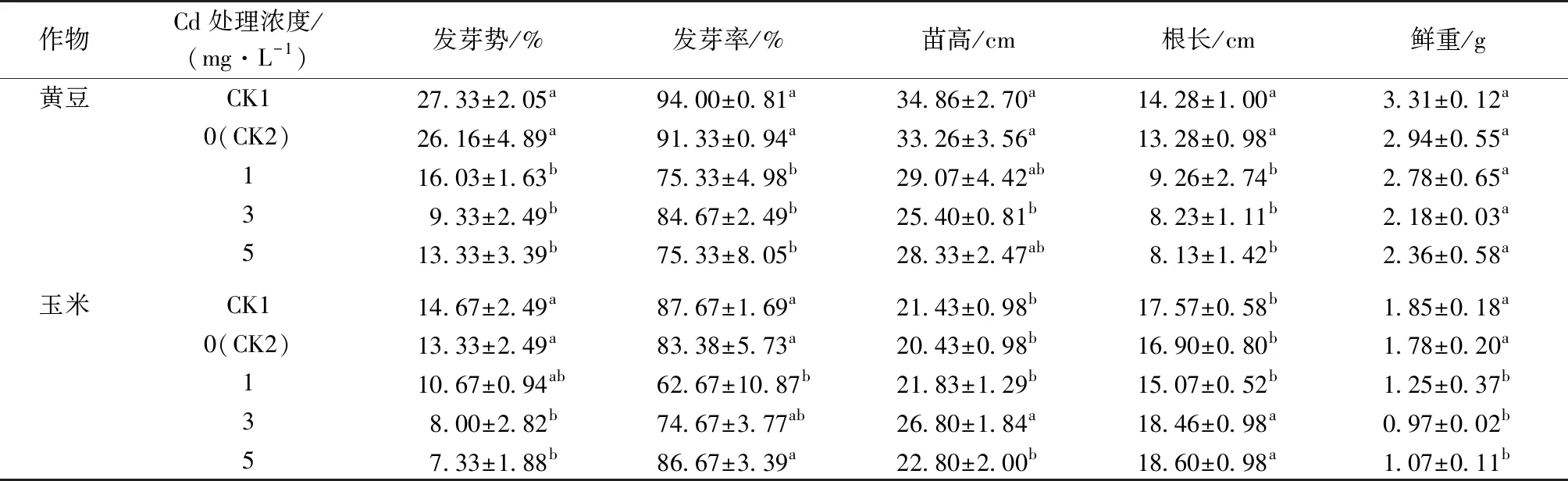
表2 不同 Cd 处理浓度下紫茉莉根系分泌物对黄豆和玉米种子发芽和幼苗生长的影响
从化感响应指数值(RI)看,不同 Cd 处理浓度下紫茉莉根系分泌物对 2 种作物发芽势均产生化感抑制作用,对发芽势的影响主要表现为延迟其发芽时间,由原来的12 h 延长至 24 h 甚至更久(图 2)。随着 Cd 处理浓度升高,紫茉莉根系分泌物对黄豆和玉米种子发芽势的RI值均显著低于对照(P<0.05);同一Cd 处理浓度下,除对照外,紫茉莉根系分泌物对玉米发芽势的 RI 值均显著高于黄豆,说明玉米发芽势受到的化感抑制作用显著低于黄豆。1 和3 mg·L-1Cd处理下,紫茉莉根系分泌物对黄豆和玉米种子发芽率的 RI 值均显著低于对照,且玉米的 RI值显著低于黄豆,但5 mg·L-1Cd处理下已升高至正值。中浓度(3 mg·L-1) Cd 处理下,紫茉莉根系分泌物对黄豆幼苗苗高的 RI值显著低于对照(P<0.05),玉米则显著高于对照且RI值>0,说明此时紫茉莉根系分泌物对玉米苗高起促进作用;随着 Cd 处理浓度增加,紫茉莉根系分泌物对玉米根长的 RI 值呈先下降后升高趋势,黄豆根长则均下降;同一 Cd 处理浓度下,黄豆幼苗苗高、根长的 RI 值显著低于玉米。随着 Cd 处理浓度升高,玉米幼苗鲜重的 RI 值均显著低于对照(P<0.05),黄豆则无显著变化。不同 Cd 处理浓度下,紫茉莉根系分泌物对黄豆和玉米的化感综合作用(SE)值均显著低于对照(P<0.05),且同一 Cd 处理浓度下,黄豆的 SE 值显著低于玉米(P<0.05),说明不同 Cd 处理浓度下,紫茉莉根系分泌物对黄豆的化感抑制作用大于玉米(图2)。
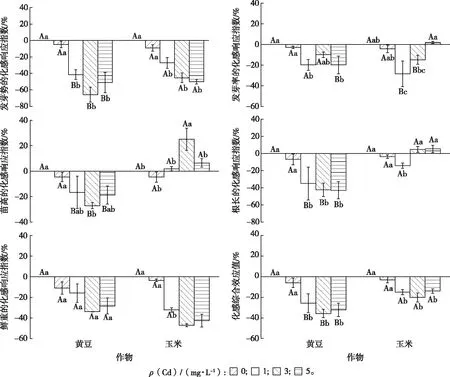
同一幅图中直方柱上方小写字母不同表示同一作物在不同Cd处理浓度间化感响应指数差异显著(P<0.05),大写字母不同表示同一 Cd 处理浓度下2个作物之间化感响应指数差异显著(P<0.05)。
2.3 不同 Cd 处理浓度下紫茉莉β-谷甾醇含量分析
先用豆甾醇、β-谷甾醇标准品配制一定浓度梯度的标准溶液,并与紫茉莉体内提取及根系分泌物收集所得供试液进行超高效液相色谱测定分析,发现紫茉莉体内及根系分泌物中均检测到β-谷甾醇,而豆甾醇并未检测到,因此对β-谷甾醇进行重点分析。
由图3可见,不同 Cd 处理浓度之间以及同一Cd处理浓度下紫茉莉不同部位之间β-谷甾醇含量均存在显著差异。同一 Cd 处理浓度下,紫茉莉根部β-谷甾醇含量均显著高于茎和叶片(P<0.05),而茎和叶片之间β-谷甾醇含量无显著变化;5 mg·L-1Cd 处理使紫茉莉根和茎部β-谷甾醇含量显著低于对照,但叶片β-谷甾醇含量与对照相比差异不显著。在高浓度(5 mg·L-1) Cd 处理下,紫茉莉根分泌物中β-谷甾醇含量较对照显著增加,但1和3 mg·L-1Cd处理下与对照无显著差异(图3)。上述结果说明,在5 mg·L-1Cd处理下,紫茉莉植株根和茎部的β-谷甾醇含量显著减少,但其根系分泌物中β-谷甾醇含量显著增加(P<0.05)。
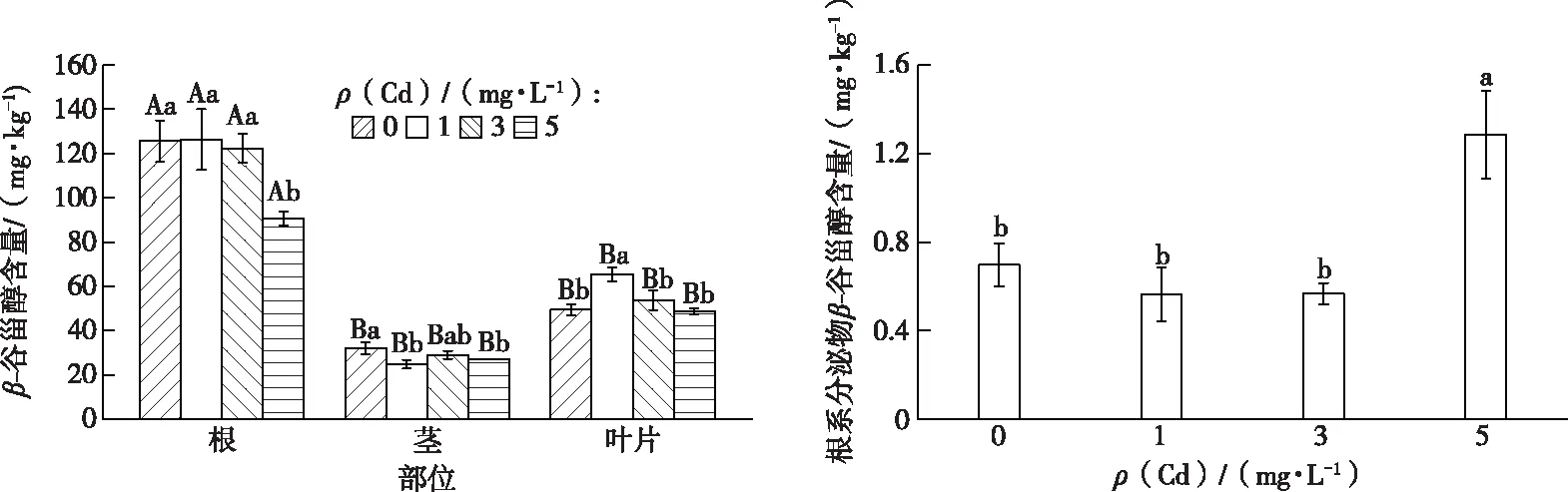
同一幅图中英文小写字母不同表示不同Cd处理浓度间β-谷甾醇含量差异显著(P<0.05),大写字母不同表示同一Cd处理浓度下植物不同部位β-谷甾醇含量差异显著(P<0.05)。
2.4 Cd处理下不同浓度β-谷甾醇对作物种子萌发和幼苗生长的影响
从种子发芽势可以看出,β-谷甾醇处理对黄豆和玉米种子萌发的影响同样表现在延缓萌发时间上,其中以黄豆种子表现较为明显。当β-谷甾醇质量浓度升至 2 mg·L-1时,黄豆发芽势比对照显著下降 37.5%(P<0.05),发芽率和苗高也显著低于对照,而此时玉米的发芽势和发芽率显著高于对照(P<0.05,表3)。随着β-谷甾醇处理浓度的增加,黄豆的根长、鲜重以及玉米的苗高、根长、鲜重均与对照无显著差异(P>0.05,表3)。
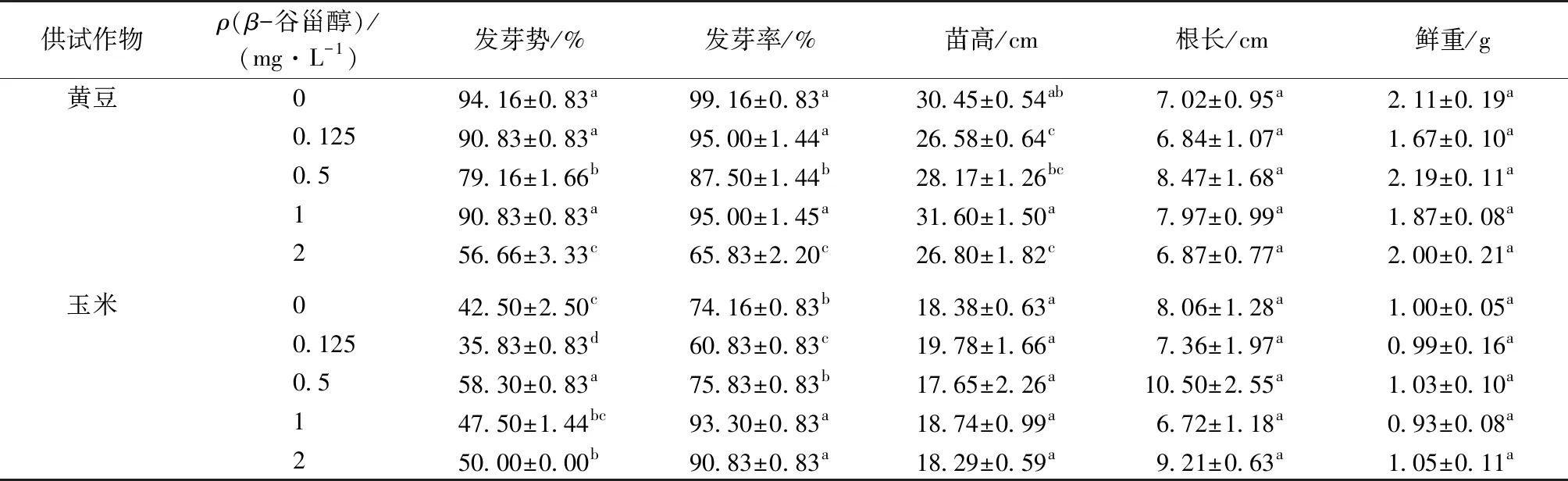
表3 3 mg·L-1 Cd 处理下β-谷甾醇对黄豆、玉米种子萌发和幼苗生长的影响
从2种作物受β-谷甾醇的化感效应(表 4)看,低浓度β-谷甾醇(0.125 mg·L-1)处理下,玉米种子发芽势和发芽率的 RI 值显著低于对照(P<0.05),但黄豆种子发芽率和发芽势的RI值均与对照无显著差异,说明低浓度β-谷甾醇处理下玉米较敏感。当β-谷甾醇浓度为 2 mg·L-1时,黄豆种子发芽势和发芽率的RI值均显著低于对照,分别为-39.82% 和-33.61%,但玉米表现为RI>0,说明此时黄豆发芽率受到的化感抑制作用高于玉米。0.5~2 mg·L-1β-谷甾醇处理下,玉米种子发芽势和发芽率的RI>0且均显著高于黄豆(P<0.05),表现为化感促进作用。在β-谷甾醇处理浓度为 0.125和 2 mg·L-1时,黄豆幼苗苗高的 RI 值均小于0并显著低于对照和玉米。0.5 mg·L-1β-谷甾醇处理下,2种作物根长均受化感促进作用;1和2 mg·L-1β-谷甾醇处理下,黄豆幼苗鲜重的 RI 值显著低于对照,但玉米无显著变化。
总体而言,Cd 处理下β-谷甾醇对2种作物的化感综合作用存在显著差异(P<0.05),随着β-谷甾醇处理浓度升高,黄豆的化感综合效应(SE)值呈先下降再升高后回落的趋势,且在β-谷甾醇低浓度和高浓度处理时SE<0,表明受化感抑制作用;玉米幼苗的SE值无显著变化。因此从化感综合效应看,β-谷甾醇对黄豆的化感抑制作用大于玉米。
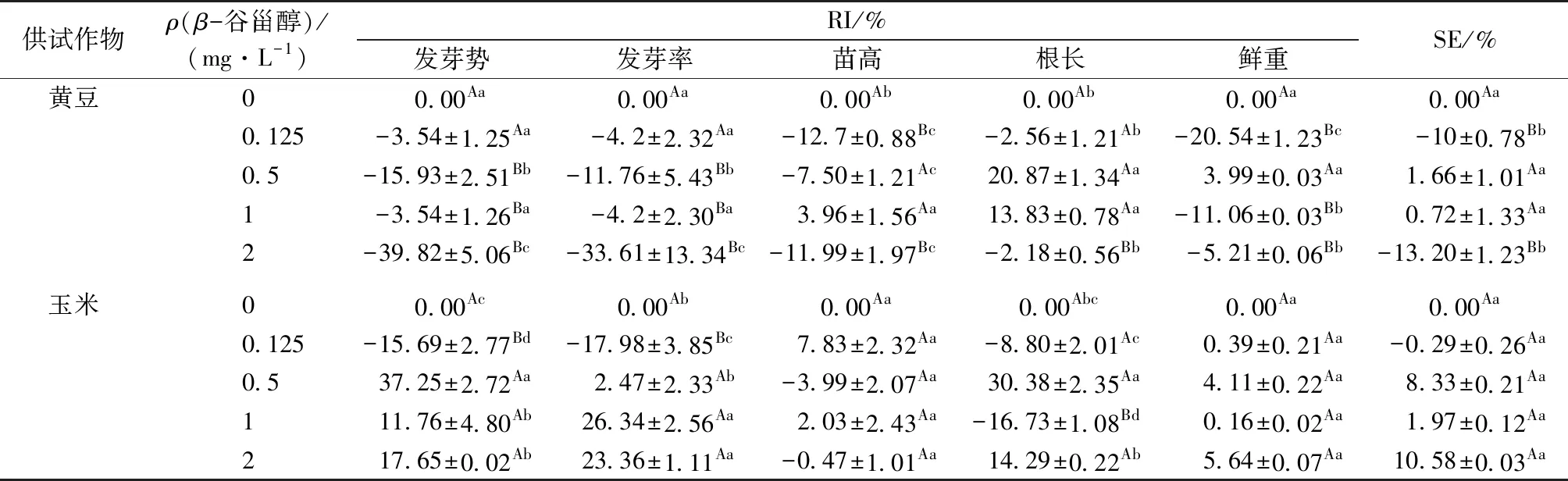
表4 3 mg·L-1 Cd 处理下β-谷甾醇对黄豆、玉米的化感响应指数(RI)及化感综合效应(SE)
2.5 Cd 处理下不同浓度β-谷甾醇对黄豆和玉米幼苗叶片抗氧化能力的影响
随着β-谷甾醇处理浓度升高,黄豆和玉米幼苗叶片的过氧化物酶(POD)、过氧化氢酶(CAT)、超氧物歧化酶(SOD)活性均发生不同变化(图4)。
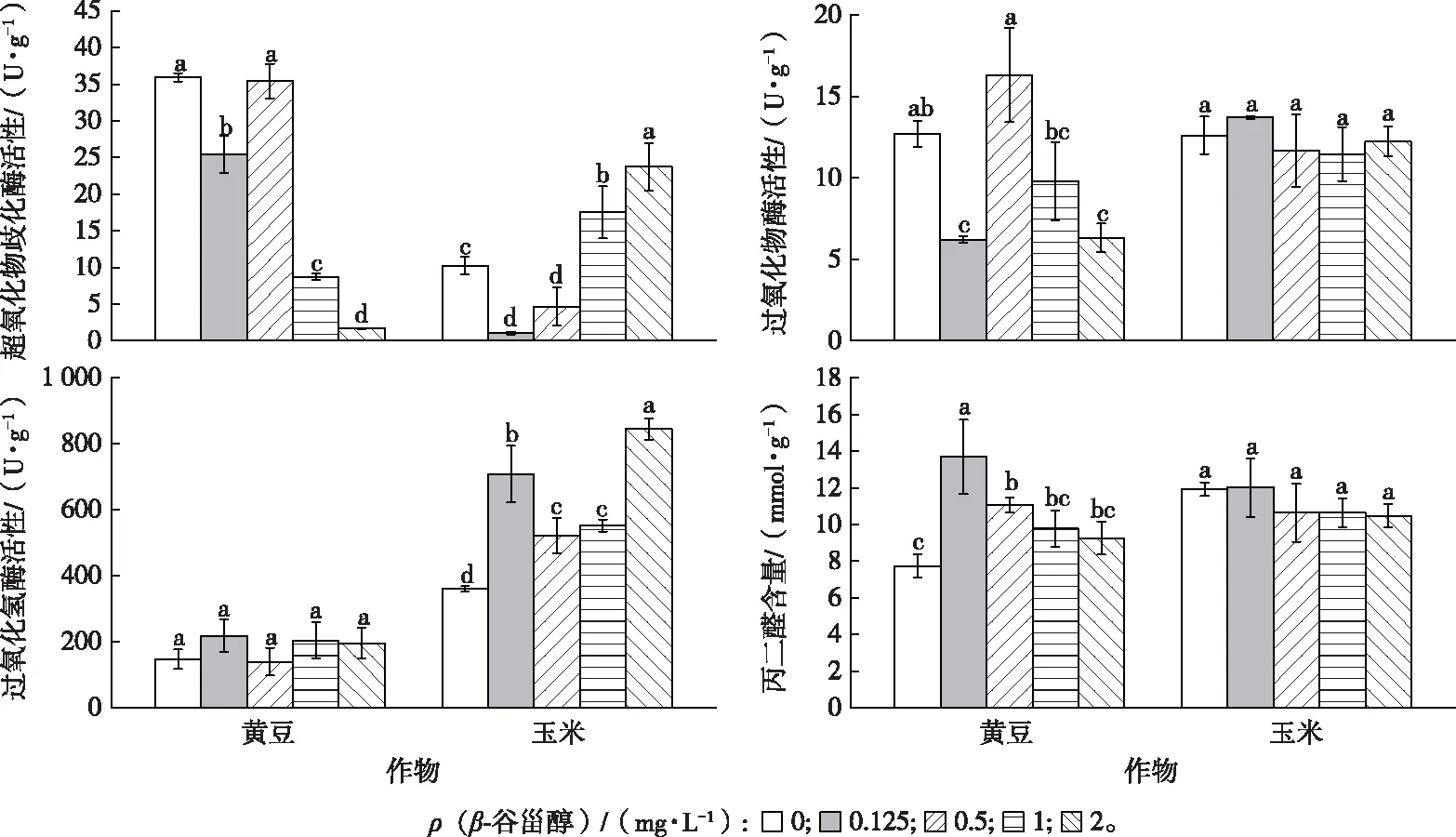
同一幅图中直方柱上方英文小写字母不同表示不同β-谷甾醇浓度处理下同一植物幼苗叶片中某指标差异显著(P<0.05)。
黄豆幼苗叶片 SOD 和 POD 活性呈先升高后下降的趋势(图4),而 CAT 活性与对照相比变化不显著(P>0.05);玉米幼苗叶片的SOD和CAT活性随着β-谷甾醇浓度的升高而升高,而 POD 活性与对照相比差异不显著。当β-谷甾醇浓度增至 2 mg·L-1时,黄豆幼苗苗高受化感抑制作用,有可能使黄豆幼苗细胞遭受损伤,抗氧化酶系统遭到破坏,导致酶活性降低。这说明经β-谷甾醇处理后,2 种作物受试幼苗的抗氧化酶系统均受到了影响(图4)。
随着β-谷甾醇处理浓度的升高,黄豆幼苗叶片细胞膜脂过氧化产物丙二醛(MDA)含量呈先升高后下降的趋势,玉米则无显著变化,表明 2 种作物幼苗叶片细胞膜受损情况不同,β-谷甾醇对黄豆幼苗叶片细胞膜的影响大于玉米。
2.6 Cd处理下不同浓度β-谷甾醇对黄豆和玉米幼苗叶片光合色素含量的影响
低浓度(0.125 mg·L-1)与高浓度(2 mg·L-1)β-谷甾醇处理下,与对照相比,黄豆和玉米幼苗叶片的叶绿素 a 和类胡萝卜素含量均显著下降(P<0.05);不同β-谷甾醇处理浓度下,2 种作物幼苗叶片的叶绿素 b 含量与对照相比均无显著变化,叶绿素总量均呈现先增加后减少的趋势(图5)。
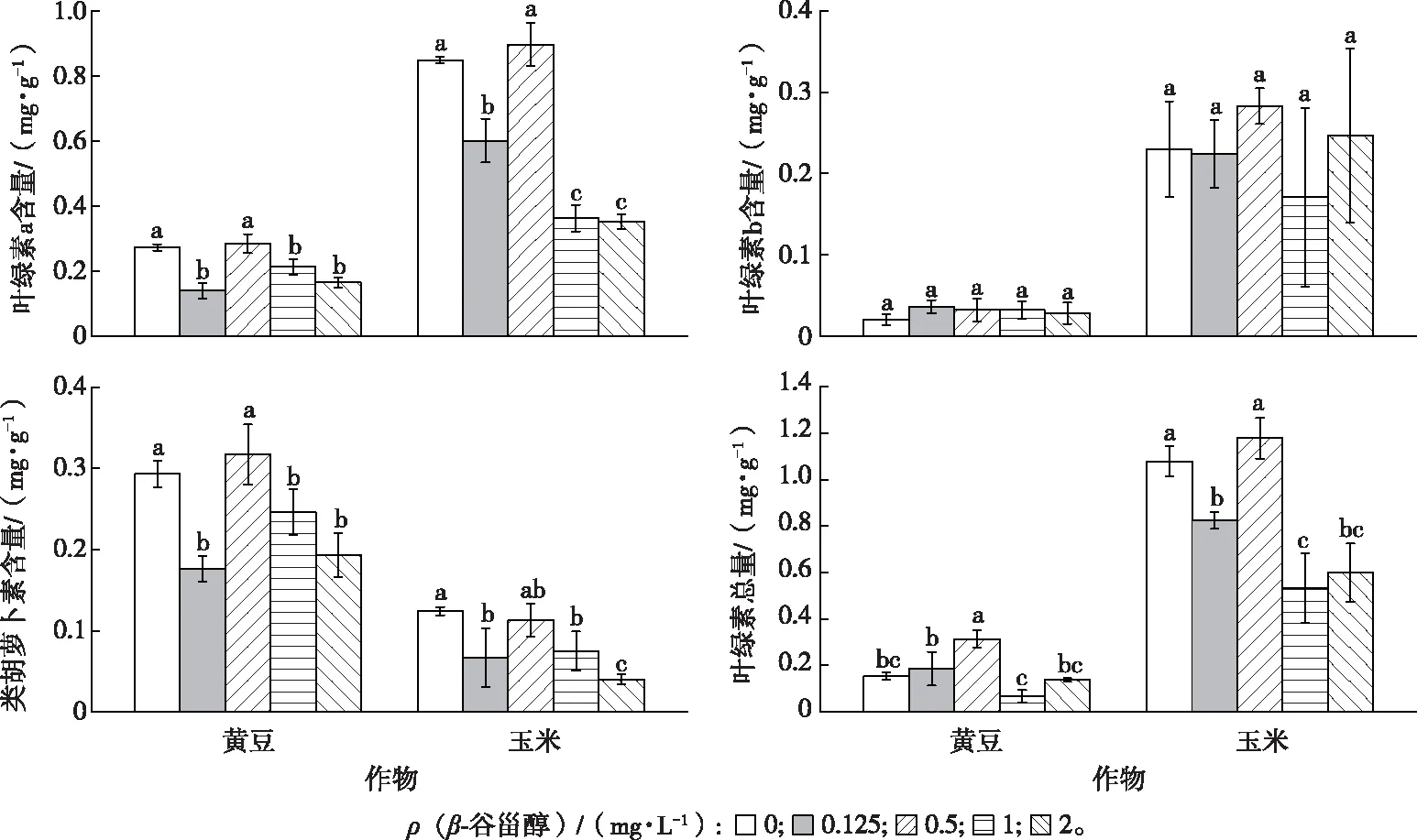
同一幅图中直方柱上方英文小写字母不同表示不同β-谷甾醇浓度处理下同一植物幼苗叶片中光合色素含量差异显著(P<0.05)。
3 讨论
3.1 不同 Cd 处理浓度下紫茉莉β-谷甾醇含量变化
对于入侵植物而言,它们大多具有较强的化感作用,即通过分泌次生化学物质以增加其在逆境下的入侵成功机率[45]。非重金属胁迫下,植物根部能分泌化感物质早已被证实,且根系分泌物中检测到的甾醇已被报道为一类化感物质[46-47]。有关紫茉莉β-谷甾醇的研究[21-22]主要集中在鉴定其成分是否存在,鲜见对其含量进行定量分析的报道,且从紫茉莉体内检测到甾醇类物质的相关研究基本在无重金属条件下进行[48]。而笔者研究主要在于明确紫茉莉β-谷甾醇化感作用强弱,并对其进行定量分析,以便测定Cd 处理下其含量变化以及由此所产生的化感效应情况。Cd处理条件下,紫茉莉根系分泌物中甾醇类的变化情况是笔者研究的一个重点。结果表明,在不同 Cd 处理浓度下,紫茉莉不仅表现出较高的耐Cd性,同时在其植株和根系分泌物中也检测到化感物质β-谷甾醇,可见紫茉莉β-谷甾醇在 Cd 处理条件下仍能正常合成。在植物甾醇与抗病性相关研究中,尤其是环境胁迫下,植物必然产生一系列的生理和化学响应,环境、生理和化感物质三者之间必然存在着联系[20,45]。因此,紫茉莉β-谷甾醇化感物质的产生和释放是紫茉莉在Cd污染的选择压力下形成的结果。紫茉莉根部作为直接接触Cd的部位,也是β-谷甾醇的主要合成和储存部位。对照中紫茉莉根和茎的β-谷甾醇含量最高,高浓度(5 mg·L-1)Cd 处理时其含量则显著降低,即无 Cd 处理最有利于紫茉莉β-谷甾醇合成和储存,高浓度 Cd 处理下则相反,且在高浓度 Cd (5 mg·L-1) 处理下紫茉莉根系分泌物中β-谷甾醇最多,其他处理则无显著变化(图3)。紫茉莉为外来入侵植物,有研究认为,生长在不利于自身环境条件下的植物,其分泌用于防御的次生代谢物质数量增加[49]。因此,低浓度 Cd 处理下紫茉莉能够正常生长,此时其分泌甾醇物质的相对成本较高,因而其分泌量较少;而高浓度 Cd 胁迫时,紫茉莉生长的潜在速度受到抑制,其生长的相对成本较高,必然会分泌较多甾醇类次生化感物质用于防御。另外,Cd 对酶活性具有一定抑制作用,且呈现“低促高抑”的情况[50]。笔者研究表明,不同 Cd 浓度处理下,虽然紫茉莉植株β-谷甾醇含量与根系分泌量呈相反趋势,但是紫茉莉植株β-谷甾醇的合成总量几乎保持不变。可见不同 Cd 处理下,紫茉莉根部β-谷甾醇合成所需的酶并未受到影响,而是通过增加其根系β-谷甾醇的分泌量,来减少紫茉莉体内β-谷甾醇的储存总量。另一方面,可能是外来入侵植物紫茉莉在高浓度 Cd 处理下,需要通过提高抗逆性等生理作用,进一步强化防御机制中重要的化学机制,从而增强了相对竞争力,根系则分泌较多化感物质来获得更加有利的生态位,通过改变自身生理生化反应来适应周边环境的变化,产生化感作用来影响邻近植物的生长发育,改造环境使其更有利于自身生长。现行的GB 15618—2018《土壤环境质量 农用地土壤污染风险管控标准(试行)》中,农用地土壤 Cd 污染风险管制限值最高为4 mg·kg-1。据WANG等[51]报道,当土壤(pH值为6.5)中Cd含量为10.23 mg·kg-1时,土壤孔隙水中Cd质量浓度仅为80 μg·L-1。因此,实际土壤溶液中 Cd 质量浓度很难达到 5 mg·L-1,加之土壤的吸附作用、缓冲性能和土壤微生物的分解作用,就甾醇类物质而言,紫茉莉根系分泌的β-谷甾醇很难达到很高的浓度,所引起的化感作用有限。但需要注意的是,笔者研究仅测定了外源不同 Cd 浓度处理 14 d后紫茉莉根系分泌的β-谷甾醇变化,未考虑植物根系分泌化感物质的连续释放性或瞬间性,今后需跟踪监测实际土壤环境中紫茉莉根系分泌的β-谷甾醇随生长发育时间的动态释放规律。
3.2 Cd 处理下不同浓度β-谷甾醇对黄豆和玉米种子萌发和幼苗生长的影响
研究结果表明,不同 Cd 浓度处理下,紫茉莉根系分泌物β-谷甾醇含量达 0.4~2 mg·L-1,β-谷甾醇作为紫茉莉根系分泌物中主要的甾醇类化感物质,其在紫茉莉成功入侵方面发挥了一定作用。受到环境胁迫时植物化感作用会明显增强,这种增强作用对产生化感物质的植物是有利的,但对受化感抑制作用影响的受体植物则是雪上加霜[52]。已有研究表明,在无重金属胁迫下,紫茉莉对作物种子萌发和幼苗生长具有化感作用,对西瓜表现为化感抑制作用,而对枸杞则为促进作用[15]。同时,有研究表明,紫茉莉浸提液对小麦等作物地上部的化感作用抑制率达-85%~-60%[53]。因此,笔者选取中等浓度Cd污染水平(3 mg·L-1),结合根系分泌物中的β-谷甾醇含量,通过外源添加β-谷甾醇进一步确认β-谷甾醇对黄豆和玉米作物的化感作用。结果表明,Cd 处理下β-谷甾醇对黄豆和玉米的种子萌发、幼苗生长均产生了化感作用,且黄豆受β-谷甾醇的化感抑制作用大于玉米。有研究指出,紫茉莉的化感物质会降低作物的有丝分裂指数,诱导各种染色体畸变,进而推测紫茉莉的化感作用可能是其成功入侵和快速扩散的机制之一[54]。笔者研究结果与其一致,在β-谷甾醇处理下,黄豆种子萌发率有可能因细胞分裂受阻、种子胚的生长受抑制而降低,并且这种抑制具有浓度和时间效应。因此在Cd 胁迫下,β-谷甾醇的分泌会增加黄豆生长压力,对受化感影响的作物细胞膜系统造成伤害,使膜功能失调,幼苗生长期呼吸作用受损并降低对水分的吸收能力,光合作用、激素含量、细胞超微结构等也受影响[55]。然而,这种化感效应因受试作物的种类及其对化感物质的耐受性而异,与黄豆相反,玉米则表现为促进作用。
笔者研究中,Cd 处理下黄豆和玉米受β-谷甾醇的化感抑制效应与植物化感作用通常表现形式不尽相同,即无重金属污染下的“低促高抑”现象[56]。不同之处在于,虽然2种作物均受到了β-谷甾醇的化感作用,但从两者所受化感综合作用来看,黄豆表现为β-谷甾醇低浓度与高浓度处理下的化感抑制现象,玉米则表现为低浓度抑制、高浓度促进现象(表4)。相同之处在于,同一化感物质对不同作物具有不同的作用,这与辛福梅等[57]在非重金属条件下对紫茉莉化感作用的研究结果基本吻合,这是因为同种植物中次生代谢物质种类相同,因而化感作用具有相似性。此外,不同作物对β-谷甾醇的化感抑制作用敏感程度不同,这可能是种属间的遗传组成差异所致。
3.3 Cd处理下不同浓度β-谷甾醇对黄豆和玉米幼苗主要生理生化指标的影响
化感作用打破了植物细胞内原有自由基产生和清除之间的平衡,导致自由基积累和膜脂过氧化损伤,细胞正常生理功能受到影响或破坏,诱导生物膜损伤是许多化感物质抑制受试植物生长的主要机制之一[58]。受试植物的抗氧化酶活性会随植物浸提液的处理而变化,即植物的 MDA 含量会随化感物质的浓度升高而上升。当植物受到逆境胁迫时,植物细胞膜系统会发生膜脂过氧化。在该研究中,当β-谷甾醇处理浓度为 0.125 mg·L-1时,黄豆幼苗叶片 MDA 含量显著高于对照,这可能是因为在该处理浓度下黄豆幼苗叶片受到的化感抑制作用较强,引起 MDA 含量急剧升高(图4);而在该β-谷甾醇处理浓度下玉米的 MDA 含量则无显著变化,说明在 Cd 处理下玉米幼苗叶片对β-谷甾醇具有更强的阻止膜脂过氧化的能力。而黄豆幼苗 MDA 含量的积累可能是β-谷甾醇处理抑制受试黄豆生长的主要原因。植物细胞膜系统损伤的直接后果是膜通透性发生改变。一旦细胞膜的功能遭受损伤,将有可能影响植物幼苗对水分和营养的吸收,从而影响植物的正常新陈代谢。
在正常条件下,活性氧的产生和清除处于平衡状态时,植物受到的危害较小。 SOD作为超氧自由基清除剂,在适度逆境条件下,其活性升高可以增加植物的抗氧化能力; CAT主要存在于细胞的过氧化体中,能够清除细胞内过多的 H2O2,以使细胞内 H2O2维持在正常水平,从而保护细胞膜系统。POD 是植物体内常见的氧化还原酶,具有清除H2O2、有机氢的过氧化物等功能[59]。黄豆幼苗在相应β-谷甾醇浓度下的综合化感效应与SOD和POD活性先上升后下降的变化趋势保持一致,当β-谷甾醇进入黄豆幼苗体后,对黄豆幼苗有害的过氧化物含量增加。玉米幼苗的 SOD 和 CAT 活性随β-谷甾醇处理浓度的升高而逐渐升高,从而使玉米幼苗获得较好的抗逆能力,这可能是玉米幼苗为抵御β-谷甾醇化感物质诱发的氧化损伤而产生的生理应答,以减轻其受到的化感抑制作用,与玉米幼苗在β-谷甾醇高浓度处理下受到的化感促进现象保持一致。当然,因为黄豆幼苗的 CAT 活性和玉米幼苗的POD活性变化不显著,所以β-谷甾醇对作物是否通过化感作用产生较多的活性氧,还需进一步测定其幼苗中活性氧含量来加以验证。
Cd处理下2种作物幼苗在不同浓度β-谷甾醇处理后,其抗氧化酶系统均受到一定程度的影响,细胞清除自由基的能力降低,促使细胞降低了抵御β-谷甾醇化感作用的能力。同样,随着β-谷甾醇处理浓度升高,黄豆及玉米幼苗的叶绿素总量存在不同的变化趋势,与β-谷甾醇对这 2 种作物幼苗苗高的抑制作用一致(表 4)。因此,紫茉莉根系分泌物中存在的β-谷甾醇会对作物产生化感作用,降低其叶绿素含量,减弱其光合作用,从而影响作物的生长发育。综上所述,通过β-谷甾醇对 2 种作物生理生化指标的影响来看,外来入侵植物紫茉莉根系分泌物中β-谷甾醇的化感作用影响了作物的生理生化过程。
该研究结果表明,不同 Cd 处理浓度下紫茉莉根分泌物β-谷甾醇含量不同,而不同浓度β-谷甾醇处理下 2种作物幼苗对β-谷甾醇的抗性也不同。可以肯定的是,虽然β-谷甾醇仅占紫茉莉化感物质的一小部分,但其化感抑制作用的确存在。由此可见,在 Cd 处理下,外来入侵植物紫茉莉自身的竞争能力中化感作用并没有减弱,并可能随其根分泌物β-谷甾醇含量增加而提高。因此,在将紫茉莉作为先锋物种修复不同Cd污染程度土壤时可选择不同作物进行间作。虽然其对共存作物产生的化感作用不容忽视,但这种化感作用的强弱与土壤溶液的Cd浓度有关,可利用其可能产生的促生作用实现与作物的互利共生,如β-谷甾醇对玉米的化感综合效应多表现为促进作用(表4)。
Cd处理下外来入侵植物紫茉莉仍会通过释放化感物质影响其他植物生长,为自身迅速生长、扩散创造条件,导致生物入侵现象出现。单纯运用紫茉莉等外来入侵植物修复某一农田土壤时,容易造成生态结构单一和生物多样性减少,进而引发生态危机。因此,若能够找到合适的共生作物与其间作或套作,不仅能够增加物种多样性,稳定生态系统,还能实现“边生产边修复”。例如,QIN等[60]将Cd富集植物续断菊(Sonchusasper)与玉米间作,发现续断菊根系分泌的草酸和柠檬酸含量比单作时分别升高43.8%和75.4%,但玉米根系分泌的这2种有机酸含量则分别降低18.5%和18.7%,进而影响到根际土壤的生物有效态Cd含量,使续断菊对Cd的富集增加,而玉米对Cd的富集减少。笔者研究结果表明,紫茉莉对黄豆的化感抑制作用高于玉米,因此可优先选择玉米与紫茉莉进行Cd污染土壤的间作修复。同时,笔者课题组前期已筛选出一些重金属低积累玉米品种,如“雅玉 69”“盛农玉10”等[61],这样就更降低了玉米的食品安全隐患,既能实现作物的良好生长,又能发挥外来入侵植物紫茉莉进行Cd污染土壤修复的生态功能。
4 结论
(1)不同 Cd 处理浓度下收集的紫茉莉根系分泌物对黄豆和玉米均能产生化感抑制作用,且黄豆受到的化感抑制作用大于玉米。
(2)在高浓度 Cd (5 mg·L-1)处理下,紫茉莉根和茎叶β-谷甾醇含量显著减少,但其根系分泌物中β-谷甾醇含量显著增加,即随着外源 Cd 处理浓度增加,入侵植物紫茉莉根系会分泌更多β-谷甾醇,引起更强的化感作用。
(3)在不同Cd 处理浓度下,紫茉莉根系分泌物中β-谷甾醇含量范围为 0.4~2 mg·L-1。外源添加中(1 mg·L-1)、高浓度(2 mg·L-1)β-谷甾醇时,玉米受化感促进作用。不同浓度β-谷甾醇处理条件下,黄豆和玉米幼苗叶片中抗氧化酶、丙二醛以及叶绿素含量发生了不同程度的变化,与其受到的化感作用一致,玉米幼苗表现出相对较强的抗氧化调节机制以应对β-谷甾醇产生的化感效应。因此,可选择玉米与紫茉莉进行Cd 污染土壤的间作修复。
猜你喜欢
课外生活(小学1-3年级)(2021年8期)2021-08-28
基层中医药(2021年1期)2021-07-22
中国粮油学报(2018年12期)2018-03-19
现代园艺(2017年23期)2018-01-18
中国粮油学报(2017年2期)2017-08-07
金色少年(奇趣科普)(2017年6期)2017-07-25
新疆农垦科技(2016年10期)2016-06-15
中国粮油学报(2016年5期)2016-01-23
杂草学报(2015年2期)2016-01-04
西藏大学学报(自然科学版)(2015年1期)2015-12-25
