
天水麦积山油松树轮宽度对气候变化的响应及其机制
2023-03-07 09:04姚岱均惠俞翔王凯欣
干旱区研究 2023年1期
姚岱均, 刘 康, 惠俞翔, 王凯欣
(西北大学城市与环境学院,陕西 西安 710127)
北半球中高纬度地区树木生长受到气候因子的强烈影响,其年轮中记录了大量气候信息[1]。这些信息定年准确、分辨率高、复本易得,使树轮在树木生长对气候响应和区域气候重建等研究中得到广泛应用[2]。树木年轮学研究中往往认为区域影响树木生长的因子是稳定的[3],通过研究主要限制因子与树木生长的关系可以建立两者间线性转换函数[1],并基于此重建气候。但是,20 世纪中叶以来,北半球高纬度地区许多树木年轮记录中发现了被称为“分离效应”的异常现象,即树木生长对气温的敏感性发生了异常下降[3-7],这一现象说明树木生长与气候关系不一定是稳定的。青藏高原[8]、川西卧龙山[7]、秦岭牛背梁[6]的研究也发现了“分离效应”的存在,证明这一现象在中低纬度和高海拔区域也有体现。Babst 等[9]基于全球2710 个树轮样点的研究更是指出1930—1960 年和1960—1990 年干冷地区树木生长的气温响应显著降低,而几乎全球树木生长都受到了更强烈的水分胁迫,持续的气候变化可能导致全球树木生长的气候驱动因素的再分配。因此,深入理解气候变化背景下树木生长与气候响应关系必须考虑多因子、非线性的作用过程[1,9],采用树轮生理模型是一个可行的解决方法[10-11]。
Vaganov-Shashkin 模型(VS 模型)基于逐日气温、降水和太阳辐照度模拟季节性形成层活动和细胞生成过程[1,10],得出树木逐日生长速率、生长主要限制因子和生长季日期等树木生长的重要参数,并可以对比实际采集的树轮样本验证其可靠性,因此,在探讨树木生长过程和影响因素的研究中得到了广泛应用。目前,国内外基于VS模型已经开展了大量的研究,例如,Chen等[12]应用VS模型指出气温导致的干旱是加拿大西部白云杉生长的主要限制因子,陈兰等[10]运用VS模型探究了秦岭太白红杉径向生长对气候因子的响应,发现生长季(4—8月)的气温、生长初期及7 月、8 月的降水是限制秦岭太白红杉生长的主要气候因子。VS 模型可以模拟树木生长的逐日特征和各因素影响,运用VS模型有望解释树木生长与气候因子响应关系的变化机制,但目前相关研究还较少。
本文选择位于西秦岭北坡,兼受东南季风、西南季风和高原季风影响,气候系统独特复杂、对气候变化异常敏感[13]的天水麦积山地区作为研究区,以常用于树木年轮学研究的油松(Pinus tabulaefor⁃mis)作为树轮样本来源,探讨油松生长对气候变化的响应,并基于VS 模型探讨其机制,以期为理解气候变化背景下该区域油松对气候的响应机制和评估气候重建的可靠性提供依据。
1 研究区概况和方法
1.1 研究区概况
甘肃省天水麦积山风景名胜区(34°07′~34°28′N,105°56′~106°10′E)位于西秦岭北支东端(图1),海拔1450~2100 m,毗邻黄土高原,北邻渭水,东接嘉陵[14],是黄河水系和长江水系的分水岭;气候上属于暖温带湿润半湿润气候区,年平均气温9~13 ℃,年降水量420~660 mm,多集中在6—9 月,无霜期170~220 d,土壤以山地棕壤为主[15],植被主要属暖温带针阔叶混交林[16]。

图1 麦积山采样点及气象站点分布Fig.1 Distribution of tree-ring sampling site in Maijishan Mountain and meteorological station
1.2 样芯采集及年表建立
分别于2021年3月和5月两次在麦积山北坡闫山瀑布(34.34°N,106.02°E,海拔1720 m)设立采样点,在胸高1.3 m处使用生长锥在不同方位采集2根样芯,共采集24棵油松,获得48棵样芯。
在实验室对采集的树芯按照通用的树轮处理方法进行预处理,利用LinTAB 年轮宽度测量仪在0.01 mm 水平上测定轮宽,并使用COFFECHA 对测量结果进行检验,剔除误差较大的样芯,最后保留30 根样芯用于构建年表。利用ARSTAN 程序的负指数函数对树轮宽度数据进行去趋势和标准化,最终得到差值年表(RES)、自回归年表(ARS)及标准化年表(STD)。标准化年表主要排除了树木自身生长趋势造成的干扰,仅保留大部分高、低频环境信号[17],选用标准化年表进行研究(图2)。

图2 树轮指数与样本量Fig.2 Tree-ring index and sample size
1.3 气象资料
气象数据选用的是离采样点最近的麦积气象站(34.57°N,105.87°E,海拔1085.2 m)的逐日观测数据,数据来源于资源环境科学数据中心(https://www.resdc.cn/Default.aspx)提供的中国气象要素站点观测逐日数据集。根据麦积气象站1980—2019 年观测资料(图3),麦积站多年平均年降水量523.13 mm,8 月最多(92.94 mm),12 月最少(3.04 mm);多年平均气温11.45 ℃,7 月最高(23.33 ℃),1 月最低(-1.71 ℃)。

图3 1980—2019年多年平均月均温和月降水量Fig.3 Monthly averaged temperature and precipitation during the period 1980 to 2019
已有研究指出20世纪90年代西秦岭区域普遍发生了气温突变[18-20]。通过Mann-Kendall检验[18]方法对麦积山1980—2019 年的年平均气温进行分析(图4),1997年麦积山气温发生了突变,这与已有研究结果基本一致[18]。Babst 等[9]指出树木生长与气候因子的响应模式与气温有关,因此,本文将1997年作为研究油松生长与气候因子关系的时间分界点(1980—1997年,1998—2019年)。

图4 1980—2019年麦积山年平均气温的M-K检验Fig.4 The M-K test of the annual average temperature in Maijishan Mountain from 1980 to 2019
1.4 VS模型
VS模型自诞生以来已经开发和改进多个版本,本文采用的是VS-GENN(http://vs-genn.ru/)提供最新的VS-oscilloscope,其与过去版本在基本原理上一致[1],即将程序分为4 个模块,分别为数据输入模块、基本模块、用来估计各季节细胞数量和大小的模拟形成层模块和数据输出模块。VS-oscilloscope计算树木生长速率采用的公式如下:
式中:Gr(t)为树木年轮总体生长速率;GrE(t)、GrT(t)、GrW(t)分别为光照E、气温T和土壤水分W引发的逐日生长速率。VS-oscilloscope与过去版本的VS模型主要差别是改进了GrE的计算公式[1],在不考虑融土的条件下,采用的水平面上每日接收的光照E的计算公式为:
式中:Isc是太阳常数;r是地球大气层外正常入射的太阳辐射强度与太阳常数的比值,无量纲;L是纬度(°);δ是太阳斜角(°);ω是日落时角(弧度)。
2 结果与分析
2.1 年表统计特征分析
从油松标准化年表统计参数表中可以看出(表1),年表有较高的平均敏感度,表明油松对气候要素变化敏感;一阶自相关系数较低,表明油松受前一年气候要素“滞后效应”的影响较小;样芯平均、树间和树内相关系数均显著(P<0.05),信噪比较高,第一主成分为0.377,总体代表性大于90%,这些都表明年表具有较高的可信度。同时,以0.85作为总体代表性的最低阈值[17](图2),以此为依据建立可靠的年表(1980—2019年)。

表1 年表统计特征及公共区间分析Tab.1 Chronology statistics and the results of common interval analysis
2.2 树轮指数与气候要素的波动特征
将1997 年作为研究油松生长与气候响应时间的分界点(1980—1997 年,1998—2019 年),并通过独立样本t检验比较两个时段气候因素和树轮指数差异[5]。由麦积山平均气温、降水和树轮指数在两个时段的箱线图可以发现(图5),1998—2019 年的气温显著的高于1980—1997年(P<0.05),降水和树轮指数的变化则不显著。从气候要素上,1980—1997 年与1998—2019 年最明显的差异是气温的显著增加。

图5 1980—2019年树轮指数与气候要素的箱线图Fig.5 Boxplots of tree-ring chronology and climatic variables in 1980-2019
从气温与降水的变化趋势上可以看出(图6),除1月、10月、11月外,其他各月平均气温和全年平均气温都至少在90%的显著性水平上显著增加;而降水仅有2月在90%的显著性水平上增加。1980—2019年麦积山暖干化趋势显著,这意味着树木生长可能受到更强的水分胁迫。

图6 1980—2019年气候要素变化趋势Fig.6 The trend of climatic variables change in 1980-2019
2.3 树轮指数对气候要素的响应
由于标准化年表的一阶自回归系数较低,麦积山油松生长受前一年气候要素“滞后效应”影响较小。因此,选取当年逐月气温、降水与标准化年表进行响应分析。将1980—2019年、1980—1997年和1998—2019年3个时间段的气候要素与标准化年表分别进行相关分析(图7),1980—2019 年油松生长与5月气温(P<0.01)、7月气温(P<0.05)呈显著的负相关,与6月降水(P<0.1)、7月降水(P<0.05)呈显著的正相关,表明5—7月高温导致的干旱可能限制了油松的生长,此时降水使土壤湿润程度增加可以缓解干旱,促进油松生长。1980—1997年油松生长与5 月气温(P<0.01)、6 月气温(P<0.1)、8 月气温(P<0.1)呈负相关,6 月降水(P<0.05)呈显著正相关,整体上与1980—2019年区别不大,均是末春与夏季气温导致的干旱对油松生长产生了限制,而降水有助于缓解干旱,促进油松生长。1998—2019年的响应模式较1980—1997年则有了较大变化,5月、6月、8月的气温和6月降水与油松生长的关系均不显著,7月气温、降水与油松生长的关系均由不显著变为显著(P<0.05);秋冬季节的气象要素与油松生长表现出了相关性:10月气温(P<0.1)表现为正相关,12月气温(P<0.05)、10 月降水(P<0.01)表现为负相关。油松生长与气候的响应模式在1997 年后发生了明显改变。

图7 油松标准化年表与各月气候要素的相关性Fig.7 Correlations coefficients between he standardized chronology and monthly climatic variables
为了进一步分析油松生长与气候要素相关性的变化,选择1997年前后表现出明显变化的5月、7月、10月、12月气温和6月、7月、10月降水进行10 a窗口的向前滑动相关分析(图8),5月气温与油松生长的相关性不断下降,10 月、12 月气温,7 月、10 月降水的相关性不断增加,而6 月降水、7 月气温的相关性则表现出趋势相似的震荡。这表明麦积山油松生长与气候响应模式变化有相关性下降、相关性上升和相关性震荡3种模式。

图8 年表序列与气候要素的向前滑动相关分析Fig.8 Correlation analysis of forward evolutionary intervals between chronology and climatic variables
2.4 VS模型模拟
VS模型需要输入麦积山逐日气温降水数据、采样点的纬度和生长参数,通过调整合适的生长参数使模型模拟结果接近实测数据[1,10-11]。生长参数可通过实测资料和模型模拟调整确定[10],模型模拟是指根据已有的参数范围给出估计的参数初始值,根据模型模拟的结果与实测值进行对比,不断调整模型参数,当模拟值与实测值一致性够高时认为此时的各个参数的数值基本合理。由于一些生长参数对树木生长的作用相互拮抗,这种方法获得的参数存在错误的可能性[1,21]。因此,本文查阅了研究区的植被[22-23]、土壤[24]、气候[25]等资料,结合实测结果和模型模拟共同确定合适的生长参数(表2)。
采用表2 中列出的参数对油松生长进行模拟(图9),1980—2019 年模型模拟与实测值的相关性为0.59(P<0.05),1980—1997 年和1998—2019 年也均达到了95%显著性水平,说明模拟拟合程度较好,可以真实地反映麦积山油松生长情况[10-11]。

图9 模拟轮宽序列和实测轮宽序列对比Fig.9 Comparison of simulated and measured radial growth series

表2 麦积山油松生理过程模拟参数Tab.2 Physiological process simulation parameters of Pinus tabulaeformis in Maiji Mountain
2.5 树轮指数对气候要素的响应及变化的解释
由VS 模型模拟的逐日平均总体生长速率(Gr)和土壤湿度(GrW)、气温(GrT)及光照(GrE)引发的生长速率可以看出(图10),由气温引发的生长速率均呈双峰型,在150~250日(5—8月),气温对油松生长速率的贡献下降,这是由于气温已经超过VS模型所给出的油松最适生长气温上限(23 ℃)。当5月气温过高时,就有可能使气温引发的生长速率的峰值提前,对油松生长起到负面作用,这解释了为何1980—2019 年整体上5 月气温与油松生长呈负相关。8月气温在1980—1997年与油松生长的负相关原因可能是高温时气温超过油松最适生长气温上限,高温对油松生长起到抑制作用,推迟了气温引发的生长速率的第二个峰值所致。

图10 模拟的麦积山油松生长速率Fig.10 Simulated growth rate of Pinus tabulaeformis in Maijishan Mountain
气温引发的生长速率谷底在6—7 月200 日前后。由于干旱缺水,土壤湿度引发的生长速率也在100 日左右开始下降,并在250 日左右下降到最低值。此时高温可以进一步降低气温引发的生长速率,加剧土壤的干旱,而降水可以缓解干旱,提升土壤湿度引发的生长速率。这解释了6月、7月油松生长与气温的负相关、与降水的正相关。
为了分析油松生长过程的变化,分析了麦积山油松1980—1997 年与1998—2019 年逐日生长速率的差值和1980—2019 年生长季的变化(图11),1998—2019年和1980—1997年相比,全年土壤湿度引发的生长速率都有所减少,反映油松受到了更强的水分胁迫。气温引发的生长速率在30~150 日和270~330日,即2—5月的末冬和春季、9—11月的秋季有所增加;在150~270 日,即5—9 月的春末和夏季有所下降;而在年初到30日、330日到年末变化不大。这些变化是研究区变暖所导致,2—5月、9—11月没有充足热量供油松生长,因此,2—5月和9月的增温使油松生长的热量供应相对更加充足,最终导致生长季延长(P<0.05);5—9月的气温往往已经超过VS 模型所给出的油松最适生长气温上限(23 ℃),此时继续增温对油松的限制更强;非生长季的12 月和1 月增温并不显著,因此,对油松生长影响不大。总体生长速率的变化趋势与气温引发的生长速率基本一致,总体生长速率差值与气温引发的生长速率差值的相关性(0.66)也要大于与土壤湿度引发的生长速率差值的相关性(0.45),这反映了气温引发的生长速率的变化是导致总体生长速率变化的主要因素。但总体上生长速率总体变化与气温引发的生长速率的变化也有区别,主要体现在冬末和春季的增加较气温引发的生长速率结束得更早(90日左右),秋季的增加则开始较晚(290日左右),这反映了总体上生长速率受到土壤湿度引发的生长速率减少的影响,增温导致的干旱抵消了部分充分热量对油松生长的促进作用。
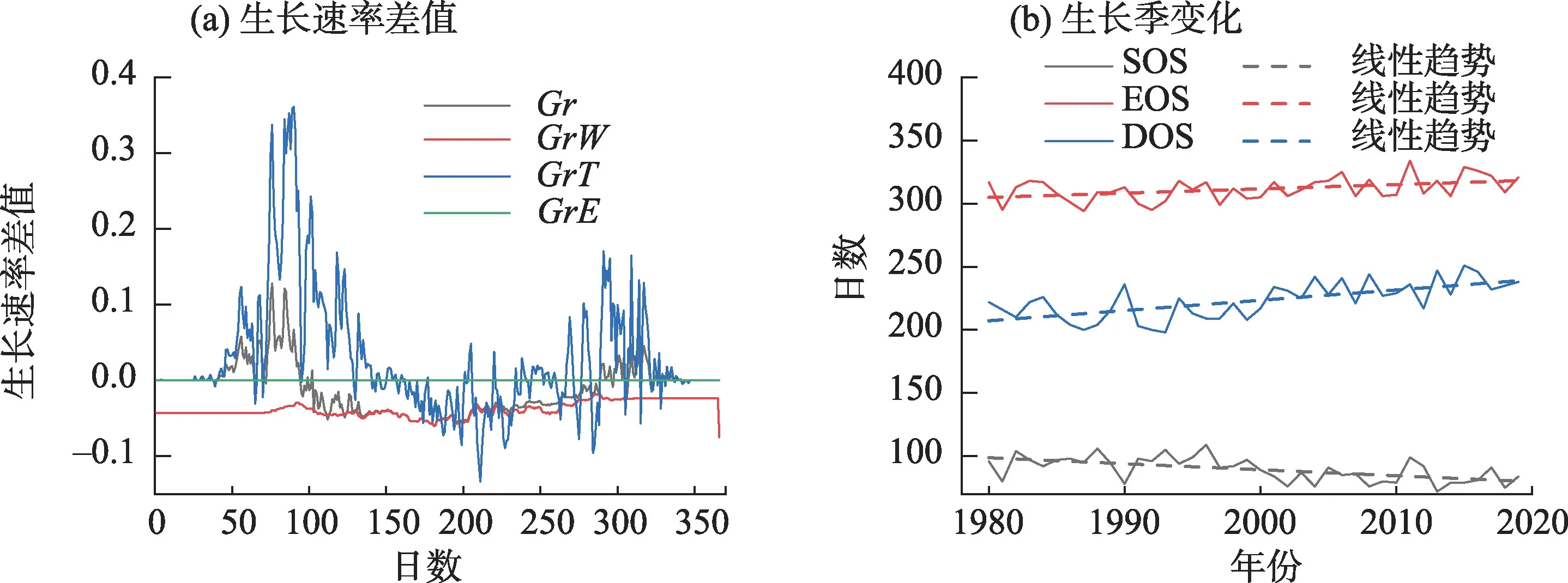
图11 麦积山油松生长速率差值及生长季变化Fig.11 Growth rate difference and change of growing season of Pinus tabulaeformis in Maijishan Mountain
气候变化导致的油松全年生长速率的变化使油松生长与气候响应模式发生了改变。5月气温与树轮指数负相关性下降的原因可能是2—5 月变暖导致油松生长的热量供应更充分,气温引发的生长速率更高,抵消了5月高温的负面影响。7月降水与树轮指数正相关性的增加可能是7月暖干化使高温和土壤湿度低对油松生长的胁迫更严重,气温和土壤湿度引发的生长速率都更低,降水可以降低气温,增加土壤湿度,增加气温和土壤湿度引发的生长速率,最终促进了油松的生长。10月气温与树轮指数的正相关、降水与树轮指数负相关性增加的原因可能是10 月气温引发的生长速率相对增加对油松生长起到促进作用,促进作用超过了土壤湿度引发的生长速率相对减少对油松生长的抑制作用,导致总体生长速率差值由减少转变为增加;此时更高的气温可以增加气温引发的生长速率,促进油松生长;降水伴随的降温使气温引发的生长速率下降,延缓了总体生长速率差值由减到增的转换,对油松生长产生负面影响。12月植物处于休眠状态,气温很难对油松生长过程产生影响,因此,12 月气温与树轮指数负相关性的增加可能仅是相关关系而非因果关系。6月降水、7月气温与树轮指数相关性的震荡变化则可能反映了研究区气候存在年际波动和周期性变化。
3 讨论
理解树轮-气候响应关系不稳定的“分离效应”和更本质的树木生长的气候驱动因子的变化有助于预测气候变化对全球生态系统物质循环和能量流动的影响[9]。本文研究了天水麦积山区域1980—2019年油松生长对气候响应的变化,发现1997年前后油松生长与气候响应关系发生了明显改变。
树轮-气候响应关系可以与基于树轮的气候重建结果相对照,通过验证历史时期树轮-气候响应关系的稳定性以评估气候重建的科学性[3]。Chen等[26]、Song等[13]分别基于天水石门山采集的油松样本分析了其与1952—2011年、1953—2008年气象要素的响应关系,均发现与5月、6月、7月的气温存在相关关系,并分别重建了天水1666—2008 年5—6月最高气温和1630—2011 年5—7 月平均气温。这表明5 月、6 月、7 月气温是天水油松生长的主要限制因子。本研究发现,5 月气温与油松生长的相关性在1991—2000年后才有比较明显的下降,1998—2007年后相关性才下降到95%显著性水平以下,此前两者相关性比较稳定。因此,树轮气候重建中树轮-气候响应关系保持稳定的假设基本成立,Chen等[26]、Song等[13]所进行的气候重建比较可靠。
Wu等[27]基于VS模型模拟了天水在内的甘肃东部3 个样点的油松生长,本研究中VS 模型参数与Wu等[27]选择的参数主要区别在油松根深及相关的土壤湿度参数上,Wu 等[27]选择的油松根系深度为500 mm,本研究选择的油松根系更深(1600 mm)。天水[22]与麦积山生境类似的甘肃武都五凤山[28]油松生理的研究中均已证明油松根深可以达到1500~1600 mm,更深的根系可以使油松吸收深层土壤的水分,对油松的生理过程产生影响。因此,将模型中油松根系深度设置为1600 mm可以更好的反映油松的实际生长情况。本研究也对一些土壤湿度参数进行了调整。史江峰等[29-30]基于VS 模型对贺兰山油松的研究中,也对油松根系深度和土壤湿度参数进行过类似的修正。
基于VS模型,本研究发现气候变化使油松全年生长速率发生了变化,进而改变了油松生长与气候响应的关系。麦积山变暖使春秋季油松生长的热量供应充分,显著地提前和延后了生长季开始日和结束日;同时夏季油松生长受到更强的高温和干旱胁迫。生长季开始和结束时间附近几日的气温、降水往往对油松生长产生较为显著的影响,生长季变化改变了各月气候因子与油松生长的相关性,突出表现为5 月气温与树轮指数负相关性的下降、10 月气温与树轮指数正相关性和降水与树轮指数负相关性的上升。由此可见,麦积山油松生长与气候响应模式的变化主要是由当地气候变化(特别是变暖)导致;如果变暖持续,响应模式可能发生进一步的变化。考虑到气候变暖和油松分布在我国的普遍性,特别是一些区域油松生长模式与研究区较为相近[27],类似的油松生长与气候响应模式变化在其他区域可能也同样存在[31]。
尽管利用VS模型探讨树木生长过程的研究已经非常常见,但是模型模拟并不能完全代替实际观测,也不能保证模型模拟的所有生理参数都合理[1,21]。因此,如果要进一步研究麦积山油松生长及其与气候因子的响应模式,需要对油松生长开展长期定位试验和连续监测,利用高精度实际观测数据校正模型[11]。
4 结论
通过采用树轮气候学的方法研究了气候变化背景下,天水麦积山区域油松树轮在两个时间段(1980—1997年,1998—2019年)对气候变化的响应差异,并基于VS 模型对其变化机制进行了探讨,主要得出如下结论:
(1)在天水地区气候显著暖干化的背景下,研究区油松与气候的响应关系发生了明显改变,变化存在相关性下降、上升和震荡3种模式,说明研究区油松生长与气候的关系不完全稳定。麦积山5月气温与油松生长的相关性在1991—2000 年后才有比较明显的下降,前人基于天水石门山油松树轮样本重建的1666—2008 年5—6 月最高气温和1630—2011年5—7月平均气温比较可靠。
(2)VS模型模拟的结果表明,麦积山油松生长与气候响应模式的变化主要是由气候变暖和生长季的变化引起的,如果变暖持续,响应模式可能发生进一步的变化。类似的变化在有油松生长且生境类似、气候有变暖趋势的其他区域可能同样存在。
(3)基于VS 模型模拟的麦积山油松的生长过程对真实生长过程拟合效果较好,模拟的油松生长的逐日特征和各因素影响也较好地解释了油松生长与气候因子间响应关系的变化机制。因此,可以尝试将VS 模型用于解释其他区域的树木生长与气候因子间响应关系及其变化。
猜你喜欢
南方农业·下旬(2022年4期)2022-05-24
山西林业(2021年2期)2021-07-21
宝藏(2021年1期)2021-03-10
天津农林科技(2020年3期)2020-08-13
电子制作(2019年15期)2019-08-27
中国水土保持科学(2019年6期)2019-04-26
敦煌学辑刊(2018年1期)2018-07-09
现代园艺(2017年22期)2018-01-19
星星·散文诗(2017年10期)2017-12-30
高原山地气象研究(2016年4期)2016-02-28
