
慢性心力衰竭合并肌少症发病机制的研究进展
2023-03-04 01:41汪英男
包头医学院学报 2023年2期
高 婕,汪英男
(内蒙古医科大学 ,内蒙古 呼和浩特 010059)
肌少症是一种与年龄相关的老年综合征,已被公认为一种新的疾病,引起了世界范围内的广泛关注,其特征是肌肉质量、力量和功能逐渐丧失[1]。慢性心力衰竭(chronic heart failure,CHF)是另一种重要的老年综合征,一种心血管系统的终末疾病。这两种疾病常常同时出现于老年患者这个群体,有文献报道CHF患者的骨骼肌具有多种组织学异常,2/3的CHF患者有肌原纤维萎缩和肌肉毛细血管密度降低[2]。随着中国步入老龄化社会,CHF合并肌少症作为一种难治病严重影响了老年人的生活质量,给家庭和社会带来了沉重负担。越来越多的证据表明,肌少症和CHF是相互影响的临床综合征,目前研究表明CHF合并肌少症可以通过蛋白质代谢失衡、激素水平紊乱、炎症、氧化应激、活动量降低、胰岛素抵抗、线粒体功能障碍这几种机制(如图1)促进疾病的进展,导致老年患者生理功能降低和死亡率增加[3]。所以,CHF合并肌少症作为一个亟待解决的医学难题越来越受到医学界重视。但是CHF合并肌少症的发病机制十分复杂,目前为止,CHF合并肌少症的发生机制仍在探索,给临床诊断和治疗造成了巨大阻碍。因此,我们根据现有的研究对CHF合并肌少症的发病机制作一综述,为该类疾病的临床诊断和治疗奠定理论基础。
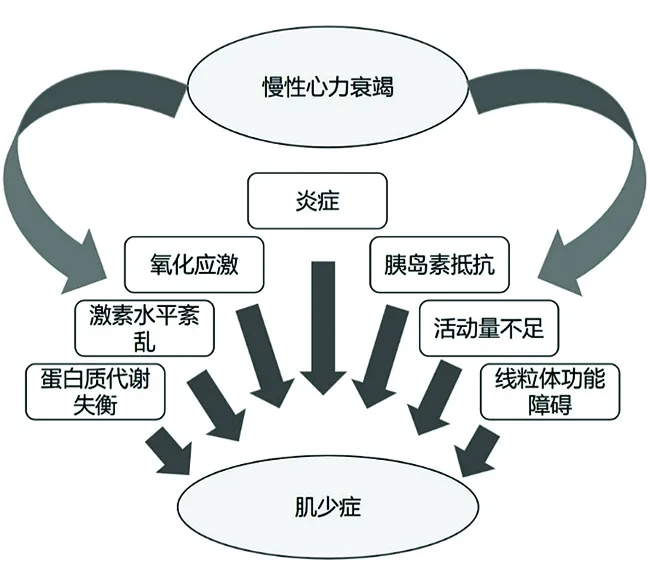
图1 慢性心力衰竭合并肌少症的发病机制
1 CHF通过蛋白质代谢失衡促进肌少症
1.1CHF通过蛋白质合成减少促进肌少症 在肌少症患者中肌肉量的减少主要是由于合成代谢反应减弱引起的,这被称为合成代谢阻力。这是由于老年人对环境因素的刺激(如食物摄入量和运动)效应表现弱化而引起。而对于CHF的患者来说,由于营养不良、炎症的刺激以及本身的高龄因素也会导致合成代谢变钝。磷脂酰肌苷3激酶(PI3K)-蛋白激酶B (Akt)-雷帕霉素靶蛋白(mTOR)信号通路是机体蛋白质合成的重要途径,它的激活可以控制蛋白质合成和氧化代谢[4]。具体而言,Akt的去磷酸化(失活)通过抑制mTOR信号级联反应来降低蛋白质翻译的启动,激活雷帕霉素复合物 1 (mTORC1)靶蛋白,抑制糖原合成酶激酶3 (GSK3),从而导致蛋白质合成减少。在CHF合并肌少症患者中,参与调节蛋白合成的PI3K/Akt/mTOR信号通路被抑制,促进蛋白分解的泛素-蛋白酶体系统(ubiquitin proteasome system,UPS)、自噬、凋亡过度激活,骨骼肌合成分解之间的动态平衡被打破,最终导致骨骼肌减少[5]。
1.2CHF通过蛋白质分解增加促进肌少症 骨骼肌中的蛋白质降解是由各种激素和代谢刺激诱导,分解代谢通过UPS、自噬和凋亡3个途径促进蛋白质降解,其中一些大蛋白被UPS降解,而线粒体和一些可溶性蛋白优先被自噬降解。
1.2.1UPS途径促进蛋白质降解 UPS是蛋白质降解的主要机制,通过刺激肌肉萎缩中肌肉无名指1(MuRF1)和肌生成素结合F盒蛋白(myogenin-engaged F-box protein,MAFbx)的表达诱导蛋白质降解,其中MuRF1是一种肌肉特异性泛素E3连接酶,在与骨骼肌萎缩相关的临床条件下被激活,MURF1已在一些肌萎缩模型中得到证实[6]。在CHF患者的骨骼肌中有UPS的过度表达,Adams等[7]研究表明UPS的激活和随后的蛋白质降解是CHF患者肌肉萎缩的主要决定因素,这促进了肌少症的发生发展。UPS的蛋白质降解过程主要由p38丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路和核因子κB (NF-κB) 两个通路激活[8]。MAPK p38负责激活肌肉特异性E3连接酶MuRF-1和MAFbx的表达。氧化应激或炎症因子也可以通过诱导NF-κB通路激活MuRF-1和MAFbx的表达。这些途径激活是CHF合并肌少症患者肌肉萎缩的关键步骤。
1.2.2自噬途径促进蛋白质降解 自噬存在于所有真核细胞中,代表着一种分解代谢过程,是一种保守的细胞保护机制,受损的细胞成分可通过溶酶体降解转化,这一过程涉及通过自噬小体和溶酶体消化的结合来大量降解细胞质成分以达到对细胞的保护。自噬在CHF中发挥重要作用,自噬溶酶体通路的功能受损导致机体蛋白质不足,是人类CHF的致病因素之一。Cho等[9]的研究结果报道,在HF动物模型中可以观察到骨骼肌自噬受损,这进一步引起了骨骼肌损伤和变性,促进了肌少症的形成。自噬过度或者减少均与肌少症相关,GSK-3α是细胞内骨骼肌自噬的显著调节剂,在生理条件下,特别是在年轻肌肉中,GSK-3α通过增强TSC2来调节mTORC1活性,从而抑制mTORC1,促进Atg12、Bnip3、LC3等自噬蛋白的合成;然而,肌少症中,较低的GSK-3α水平对TSC2的激活减弱,因此mTORC1超激活,从而抑制自噬依赖的蛋白降解[10]。因此,平衡的结构性自噬活性对CHF患者中肌细胞活力和预防肌减少症的发展至关重要,适当调节自噬信号传导可以维持CHF患者的骨骼肌质量和数量。
1.2.3凋亡通路激活促进蛋白质降解 凋亡通路的激活已被发现与肌肉萎缩有关,对凋亡通路的抑制可达到预防肌少症的作用。Liao等[11]发现运动和白藜芦醇联合治疗可以激活AMPK/Sirt1的抗凋亡信号通路,降低乙酰化p53的表达并增加Bcl-2/Bas的表达率,抑制肌细胞凋亡从而改善肌肉质量预防肌肉减少症。一种被称为“肌核凋亡”的凋亡样过程可加速消除在骨骼肌中肌核,这是一种与肌肉减少症有关的机制。鉴于心肌细胞的多核性质,细胞凋亡表现出独有的特征,凋亡途径导致肌肉纤维萎缩而不会导致其他细胞的死亡。Filippatos等[12]表明与相同年龄的健康人相比,CHF患者的肌肉中也发现了更高频率的肌核凋亡,说明CHF通过增加肌核的凋亡促进了肌少症的病程进展。
2 CHF通过激素水平紊乱促进肌少症
CHF和肌少症患者体内会出现相同种类的激素水平紊乱,这提示激素的改变可促进CHF患者中肌少症的发生发展。
2.1CHF通过胰岛素样生长因子-1缺乏促进肌少症进展 下丘脑GH-IGF-1轴是一个重要的人体内分泌代谢轴,贯穿了人类的一生对生长发育的调控,生长激素(growth hormone,GH)和胰岛素样生长因子-1(insulin like growth factor-1,IGF-1)水平的降低与肌肉质量和功能的减少有关,GH可以促进肌肉前体细胞与肌管的融合,IGF-1是骨骼和肌肉生长的关键调节因子,二者的缺乏最终导致肌肉减少症的发生[4]。IGF-1缺乏症是CHF患者中最常见的荷尔蒙缺乏症之一,在早期HF合并肌少症患者中观察到IGF-1缺乏症。一项回顾性观察研究[13]表明HF患者血浆中的人胰岛素样生长因子结合蛋白-1(insulin-like growth factor-binding protein-1,IGFBP-1)和IGFBP-1 / IGF-1比值与健康人相比,显著降低。IGF-1是骨骼肌中主要的合成代谢信号,IGF-1主要通过激活PI3K- Akt-mTOR通路控制蛋白质合成和氧化代谢,以影响肌肉生理学[4]。PI3K-Akt-mTOR通路的激活可以磷酸化失活O型叉头盒(FOXO)转录因子来阻止骨骼肌中的蛋白降解,该机制在细胞周期调控、凋亡和代谢中起着至关重要的作用[4]。FOXO转录因子在磷酸化后无法进入细胞核,从而阻止了蛋白水解相关基因的上调,如E3连接酶肌肉环指蛋白1 (MuRF-1)和肌肉萎缩F-Box (MAFbx)[4]。因此,IGF-1促进蛋白合成并且抑制蛋白降解。除此之外,骨骼肌IGF-1基因可以被切割以产生三种IGF-1亚型,IGF-1Ea,IGF-1Eb和IGF-1Ec,其中IGF-1Ec也称为机械生长因子(MGF)。当骨骼肌细胞受到外部因素的刺激或损伤时,MGF表达升高,这是激活骨骼肌卫星细胞(satellite cell,SC)以启动骨骼肌细胞修复并促进其增殖的主要因素[14]。CHF患者中IGF-1水平降低通过这些机制进一步加剧了肌少症的症状,促进肌少症的发生。
2.2CHF通过睾酮缺乏促进肌少症 睾酮水平降低是CHF患者的常见现象,不仅影响CHF的预后而且也造成了CHF患者骨骼肌功能的下降。Yoshihisa等[15]证明在男性CHF患者中,血清睾酮降低与心肌损伤、运动能力降低和死亡率较高有关。这表明睾酮水平的降低是造成运动能力降低的因素之一,睾酮降低可以改变外周血管阻力、增加心脏后负荷,引起肌肉质量损失和功能损害。睾酮对骨骼肌的积极作用涉及多种分子机制。睾酮会增加钠与中性氨基酸转运体和L型氨基酸转运体2通道的结合,可以提高对细胞内氨基酸的再利用率,激活PI3K-AKT-mTOR通路,增加蛋白质合成,同时增加I型和Ⅱ型肌纤维以及每根肌纤维的肌核数量,促进肌肉生长[16]。睾酮还可以直接激活Wnt信号通路中促进干细胞更新的细胞内信号转导子β-连环蛋白,增强成肌细胞培养系统中卫星细胞的有丝分裂活性,这同样是肥大的肌肉纤维中加入新肌核的主要来源[17]。Pronsato等[18]最近研究表明睾酮也可以通过诱导小鼠C2C12骨骼肌细胞中线粒体基因表达的上调,诱导核呼吸因子1和2(Nrf-1和Nrf-2),线粒体转录因子A(Tfam)和B2(TFB2M)以及视萎缩1(OPA1)的表达,改善肌肉线粒体的生物发生和功能,最终对骨骼肌细胞产生有益作用。除此之外,睾酮还可以通过抑制凋亡通路的激活促进肌肉的生长。睾酮可以通过激活p53和FoxO3a转录因子抑制氧化应激诱导C2C12肌肉细胞凋亡[19]。总之,睾酮可以通过多种机制调节CHF患者中骨骼肌的生长,这提示睾酮的补充治疗具有重要的意义,目前大量的临床相关试验也正在投入研究。
2.3CHF通过生长激素释放肽缺乏促进肌少症进展 生长激素释放肽(ghrelin)是一种主要在胃底区域产生的多肽,具有调节食欲、促进食物摄入和释放GH多种作用,ghrelin通过一氧化氮依赖机制激活下丘脑中的生长激素释放激素发挥作用[20]。老年ghrelin水平的降低与肌少症的发生有关。与此同时,在心血管疾病的治疗中,ghrelin也发挥一定的作用。Yuan等[21]研究发现ghrelin可以调节交感神经活动和高血压症状、增强血管活动和血管生成、抑制心律失常、减少HF发生、抑制心肌梗死后心脏重塑。除此之外,Barazzoni等[22]通过对大鼠的持续外周酰化ghrelin治疗和保留食物摄入,发现外周酰化ghrelin治疗可以减轻 CHF 诱导的组织特异性骨骼肌线粒体功能障碍、炎症反应和胰岛素抵抗。因此,ghrelin是CHF相关肌肉分解代谢改变的潜在治疗方法,对患者的预后具有潜在的积极影响,这同时也表明ghrelin可能通过线粒体功能障碍、炎症、胰岛素影响肌少症的发展。
2.4CHF通过肌肉生长抑制素升高促进肌少症 肌肉生长抑制素(myostatin,MYO),又称生长分化因子8,是转化生长因子β家族的一员。MYO是一种主要的肌肉萎缩的诱导剂,MYO基因不光在老年男性中的表达增加,在CHF患者中也显著上调。Meloux等[23]发现,在急性心肌梗死患者中 MYO与肌钙蛋白I峰值相关,这表示MYO反映了心肌损伤的程度。在CHF患者中,高表达MYO通过多种机制促进肌少症的进展。首先MYO诱导激活素二型受体(ActRIIA/B)磷酸化,激活转录因子SMAD2和SMAD3,通过激活SMAD转录因子,MYO下调参与肌源性分化的基因如MyoD、myogenin和myf5,抑制肌肉SC的激活从而损害肌肉再生能力[24]。另外,SMAD2和SMAD3的激活也会诱导MuRF-1和MAFbx的表达促进蛋白降解。除此之外,MYO被认为是肌管中产生活性氧(Reactive Oxygen Species,ROS)的有效诱导剂,可激活机体的炎症反应并诱导骨骼肌的恶化。Sriram等[25]证明在衰老过程中增加的MYO通过刺激NF-κB信号通路诱导TNF-α的产生,从而损伤骨骼肌。MYO是促进CHF患者肌少症发生的重要因素,是目前CHF合并肌少症研究热点。
2.5CHF通过血管紧张素Ⅱ升高促进肌少症 CHF患者中持续交感神经活动导致血管紧张素Ⅱ(AngⅡ)增加,最终增强醛固酮的分泌,促进盐和水的潴留,使CHF的进展进一步恶化。有文献报道Ang Ⅱ也可加重HF患者肌少症的进展,Katano等[26]在动物模型中发现,AngII输注可通过改变IGF-1信号、增加细胞凋亡以及过度激活UPS来诱导肌肉萎缩,而对肾素血管紧张素系统抑制可以降低HF患者肌肉萎缩患病率。这表明可能通过靶向降低病理性升高的血浆肾素活性进而对CHF患者产生有益的结果。一项研究表明,CHF患者血浆肾素活性降低至正常水平可通过改善心脏收缩功能,减少系统性水肿,恶病质或者肌肉减少症的发生来延缓CHF的进展[27]。
3 CHF通过炎症激活促进肌少症
炎症因子对肌肉质量和运动能力产生重要作用。最近Tuttle等[28]在一项包含168篇文章的荟萃分析发现,高水平的循环炎症标志物如C-反应蛋白(C-reactive protein,CRP)、白细胞介素6 (interleukin 6,IL-6)和肿瘤坏死因子-α (tumor necrosis factor-α,TNF-α)与握力和伸膝力显著负相关,其中较高的CRP水平与社区人群中较低的肌肉质量显著相关。IL-10是一种抑制炎症和免疫反应的细胞因子,2018年Rong等[29]进行了一项横断面研究,并报告了老年肌少症患者炎症细胞因子IL-6、抗炎细胞因子IL-10和IL-6/IL-10比值增加46倍。在CHF患者中,炎症反应可引起骨骼肌蛋白水解和消耗过多,降低CHF患者的运动能力。Koshikawa等[30]发现CHF患者血液中升高的CRP、IL-6、脑钠肽水平与骨骼肌肌肉蛋白水解呈正相关,这表明与CHF相关的炎症状态可能导致肌肉蛋白水解过多,造成骨骼肌的消耗促进肌少症的发展。这充分提示炎症是CHF和肌少症之间重要的一条共通机制。这些炎症因子以多种方式在CHF患者中引起骨骼肌消耗。Wang等[31]发现在正常、健康和衰老动物中,TNF-α的表达与卫星细胞的数量以及肌细胞减少症的发生和发展有关,系统性消融TNF-α可防止肌纤维表型的年龄相关变化和肌减少症的发生。Yalcin等[32]在一项老年人群的研究中发现,另一种抗炎因子IL-15信号的降低通过促进TNF-α介导的细胞凋亡进一步导致骨骼肌质量损失和肌肉功能下降。另外炎症因子水平的升高进一步激活了与炎症相关的NF-κB信号通路引起蛋白质的降解[33],促进肌少症的发生。除此之外IL-10作为一种抗炎细胞因子,Rong等[29]发现它通过抑制人类单核细胞和巨噬细胞的功能以及阻断促炎细胞因子(包括IL-6)的产生减轻炎症反应,是预防老年人肌肉减少症的有希望的治疗候选药物。总之,全身炎症反应的激活,调控了细胞因子的释放,其中促炎细胞因子水平的增高和抗炎因子的降低共同参与CHF患者的肌肉损失,最终炎症导致CHF和肌少症互相作用形成恶性循环,加速了疾病的恶化。
4 CHF通过氧化应激促进肌少症
氧化应激是由于ROS的产生与抗氧化防御之间的不平衡而发生的。ROS主要由线粒体产生,ROS的过量产生会导致线粒体功能和氧化能力降低,最终引起细胞发生一系列的病理变化。ROS的产生随着年龄的增长而增加,而肌肉的酶清除系统随着年龄的增长而下降,研究表明氧化损伤分子在肌肉收缩活动中的积累可加速骨骼肌的退化。ROS可以通过限制mTOR机制的靶点磷酸化真核翻译起始因子4e结合蛋白1(4E-BP1)的能力来加速蛋白水解和自噬,抑制蛋白合成[34]。CHF的特点是心肌氧化还原调节的变化,有文献报道心脏的氧化应激刺激会加重CHF患者的骨骼肌障碍[35]。据报道,黄嘌呤氧化酶(XO)是缺血组织中ROS过度产生的重要介质,Hideo等[36]证明心肌梗死后急性期的XO抑制可以预防HF小鼠的骨骼肌异常和运动不耐受。这表明氧化应激的适当调节可能对HF中的肌肉萎缩产生积极作用,氧化应激是导致CHF患者肌肉萎缩的重要信号分子,加速了肌少症的发生。
5 CHF通过活动量减少促进肌少症
活动量不足可引起肌肉的萎缩,Santos等[37]的横断面研究发现,不太热衷于运动的老年人表现出更高的肌少症风险,活动量不足可以减少机体代谢,导致减弱肌肉蛋白质的合成。此外,长时间不活动的老年人会导致胰岛素敏感性降低和较高的炎性状态,进一步对肌肉内稳态产生负面影响并且促进肌肉分解代谢。CHF由于呼吸和运动的不耐受,导致缺乏运动和长时间卧床,从而导致肌肉损耗和功能障碍。总体而言,尽管CHF相关性肌少症与体力活动之间的关系仍未得到充分研究,但现有证据表明,充分的体力活动可以预防CHF相关肌肉损失。运动可通过促进卫星细胞的增殖、线粒体和毛细血管密度产生的能量增加并且可以下调慢性炎症和抑制骨骼肌细胞凋亡等多种机制影响肌肉的变化[38]。
6 CHF通过胰岛素抵抗促进肌少症
胰岛素抵抗可导致肌少症的发生。研究表明,在老年大鼠中肌肉质量的增加会提高骨骼肌葡萄糖摄取,并提高胰岛素敏感性[39]。同时,胰岛素抵抗也是CHF发生的因素之一。Wamil等[40]的研究表明胰岛素抵抗患者与胰岛素敏感的患者相比,更容易发生CHF或死亡,这说明胰岛素抵抗可加重CHF。这是由于胰岛素抵抗时可通过抑制PI3K-Akt-mTOR通路信号抑制肌肉的增长[4],除了PI3K-AKT通路的抑制外,胰岛素抵抗后肌力也会因线粒体功能下降而降低并且触发运动神经元和肌纤维死亡。过氧化物酶体增殖体激活受体γ协同激活因子-1α(PGC1-α)是线粒体生物发生和呼吸的主要调节因子,是核编码线粒体基因(nuclear-encoded mitochondrial genes,NEMGs)的主要调节分子,它通过调控线粒体自噬改变和细胞色素C氧化酶的活性发挥作用。研究表明,胰岛素敏感性的降低以及Akt和mTOR表达的降低可通过PGC-1α信号通路的降低导致线粒体功能的损伤,受损线粒体的积累已被证明会触发运动神经元和肌纤维死亡[41]。除此之外,胰岛素抵抗还可以通过肌肉中葡萄糖的代谢紊乱引发肌肉质量降低。Consitt等[42]最近的研究表明肌肉胰岛素抵抗导致葡萄糖转运蛋白4(GLUT4)从细胞内GLUT4储存囊泡向质膜的转运减少,导致肌肉葡萄糖利用率降低。总之,在心脏和骨骼肌的肌肉细胞中葡萄糖代谢的破坏造成的能量稳态失衡导致CHF症状更加严重,胰岛素抵抗会同时危害CHF和肌少症。当身体发生胰岛素抵抗时,体内的胰岛素分泌减少,葡萄糖稳态被破坏导致葡萄糖利用障碍,而肌肉是人体吸收和利用葡萄糖的重要器官,使其对肌肉的能量供应显著减少从而导致骨骼肌含量下降和肌肉减少症。
7 CHF通过线粒体功能障碍促进肌少症进展
线粒体是细胞能量代谢的主要调节器,骨骼肌的代谢紊乱与线粒体功能障碍有关。最近的一项研究比较了年轻人和老年人的线粒体呼吸能力,发现线粒体呼吸能力和耦合控制能力随着年龄的增长而下降[43]。线粒体功能障碍是胰岛素抵抗和CHF发病机制的关键因素,最近的研究显示在CHF患者中发现骨骼肌线粒体的含量和耦合效率降低、电子传递链活性受损、线粒体膜电位降低、ETC复合物活性受损以及线粒体生物发生异常导致ATP合成减少和心脏重塑[44]。这些研究表明线粒体功能障碍是骨骼肌萎缩的基本特征,而CHF患者可能会通过线粒体障碍的恶化进一步导致肌少症的进展。因此,线粒体质量控制对于维持骨骼肌的正常功能至关重要,可用作改善骨骼肌功能的治疗靶点,其中AMPK/PGC-1α信号通路通过对线粒体的调控对肌肉发挥重要作用。Liang等[45]表明终身有氧运动可以通过AMPK/PGC-1α信号通路挽救自噬/线粒体的异常功能状态、刺激线粒体生物发生、优化线粒体动力学最终抑制老年小鼠骨骼肌质量的下降。所以有氧运动可以通过AMPK/PGC-1α信号通路活性改善能量代谢、挽救自噬不足、抑制过度凋亡和蛋白质泛素化,最终修复功能失调的线粒体、强化线粒体质量控制,改善肌原纤维的超微结构从而防止或延缓骨骼肌萎缩。
CHF与肌少症共存是一个常见的现象,肌少症作为一种老年性慢性疾病,仍常常被人们视为一种正常的衰老现象而忽视,所以需要提高对它的认识和重视程度。相对于肌少症,CHF的研究虽已有数百年的历史,但它仍然是心血管领域难以攻克的难题。二者病理生理学机制十分复杂,现有证据表明,他们之间存在共同的途径,通过以上综述笔者发现HF和肌少症通过调控蛋白质代谢、激素水平、炎症、氧化应激、活动量减少、胰岛素抵抗、线粒体功能障碍这些共同的发病途径互相作用,促使病情加重。在分子水平上PI3K/Akt/mTOR信号通路控制骨骼肌质量的合成代谢和分解代谢信号,是骨骼肌肌肉质量和功能的重要调控机制。本综述可为CHF合并肌少症的发病机制研究提供思路,并为将来两种疾病的共同治疗提出有益的启发。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
河北医科大学学报(2021年12期)2021-12-29
中华骨与关节外科杂志(2021年12期)2021-08-31
中老年保健(2021年3期)2021-08-22
海洋通报(2021年1期)2021-07-23
生物学通报(2021年4期)2021-03-16
国际放射医学核医学杂志(2020年4期)2020-07-27
天然产物研究与开发(2018年2期)2018-04-04
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
