
广东鼎湖山河岸林水翁蒲桃+蒲桃群落的物种和功能多样性及群落稳定性分析
2023-03-03 07:23:40杨桂梅周友秀杨钰华黄柳菁
植物资源与环境学报 2023年1期
杨桂梅, 周友秀, 杨钰华, 玄 锦, 黄柳菁
(福建农林大学风景园林与艺术学院, 福建 福州 350002)
物种多样性与功能多样性是生物多样性研究的重要内容,对生态系统功能和结构起着决定性作用[1-2]。物种多样性反映了群落内部结构和生物资源上的丰富度[3],不同植物群落或植被类型,由于建群种或伴生种的不同,群落内部组成、结构和功能等存在差异[4-5]。功能多样性考虑了植物群落的冗余特性和种间互补作用,表现群落内部物种功能性状的总体差异与变化范围[6]。而群落稳定性受到群落结构与功能的共同作用,表现出群落对干扰的抵抗能力及其演变与发展趋势[7]。
一直以来,多样性-稳定性间的关系备受争议。MacArthur[8]与Elton[9]形成的多样性-稳定性假说认为物种多样性与群落稳定性具有正效应,物种多样性越复杂,越能抵抗对外界的干扰,对群落及生态系统稳定就越有利。但一些学者对该假说提出了质疑,认为越复杂的生态系统(即物种多样性越高,种间联结越强)将削弱物种间的平衡,导致稳定性下降,尤其是对于自然界的一般系统,种群的稳定性不一定与植物区系的多样性相关[10-12]。20世纪90年代以来,一系列的稳定性实验结果表明:物种多样性可以促进生态系统功能的稳定性[13]。但由于每个物种对群落的功能贡献都是独特的,物种多样性及物种丧失均可能引起群落内物种功能性状的组成或结构发生改变,因此,物种及其所具有的功能属性对于维持生态系统功能及群落稳定十分重要[14-15]。姚天华等[15]对高寒草甸群落稳定性的研究结果表明:物种多样性是影响生态系统稳定性的关键,但功能多样性相较于物种多样性能更好地预测生态系统稳定性的变化。董世魁等[3]认为,物种多样性和功能多样性对不同类型群落稳定性的影响机制各不相同。物种影响功能性状的丰富度,功能决定了物种多样性与生态系统功能的关系[16]。因此,群落稳定性不仅需要通过物种的数量来反映,更需要考虑其内部不同物种所具有的功能属性[17]。目前,有关物种多样性与群落稳定性的研究已非常深入,但长期以来群落内物种及功能多样性与群落稳定性的关系的研究仍较少。因此,综合研究群落的物种、功能多样性与群落稳定性特征,有助于揭示群落的结构、演变规律及稳定性维持机制,可为群落稳定性提高和物种多样性保护提供理论基础。
水翁蒲桃(SyzygiumnervosumCandolle)+蒲桃〔Syzygiumjambos(Linn.) Alston〕群落是广东鼎湖山河岸林早期演替中的优势树种,属南亚热带地带性常绿阔叶林,是经多年演化而形成的原始次生林[18-19]。近年来,由于极端天气频发和人类活动加剧,生态环境恶化,自然保护区被波及,如旅游业的不断发展和热带台风频发,直接影响植物群落的生存与发展。自2000年黄忠良等[19]对该群落多样性调查后,越来越多的学者开始对鼎湖山不同海拔梯度或区域的物种组成、区系、物种多样性和功能性状等进行相关研究[20-21]。然而,经多年的自然演替,群落稳定性处于何种程度尚未有文献报道。鉴于此,为明晰该群落稳定性维持机制,本研究以广东鼎湖山河岸林水翁蒲桃+蒲桃群落为研究对象,进一步解释物种多样性、功能多样性和群落稳定性之间的关系,以期为鼎湖山物种多样性保护及森林资源可持续管理提供参考意见。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于广东省肇庆市鼎湖山国家级自然保护区(东经112°31′、北纬23°10′),属南亚热带季风湿润型气候,冬夏季节交替明显,年均温20.9 ℃,年降水量1 927 mm,年蒸发量1 115 m[20]。自然植被丰富,森林覆盖率达78.8%,是华南地区生物多样性最富集的地区之一,植被类型包括南亚热带典型常绿阔叶林、沟谷常绿阔叶林、山地常绿阔叶林、河岸常绿阔叶林、针叶林、针阔叶混交林、稀树灌草和灌草丛8种植被类型。样地区域海拔约60~100 m,属河岸常绿阔叶林,生物多样性丰富,群落类型为水翁蒲桃+蒲桃群落,主要植物种类有水翁蒲桃、蒲桃、鱼尾葵(CaryotamaximaBlume ex Martius)和朴树(CeltissinensisPers.)等,土壤以冲击土和赤红壤为主,具有良好的透水性[19,22]。
1.2 研究方法
1.2.1 样地设置 基于2021年7月对鼎湖山国家级自然保护区的充分踏查,于2021年8月进行群落学调查。在以水翁蒲桃和蒲桃为主要优势种的河岸常绿阔叶林中,选取林分结构完整、林相整洁且外界干扰较少的群落设置样地。沿河岸每隔100 m设置1个面积20 m×20 m的乔木样方,共布设30个乔木样方,在乔木样方内以对角线四角和中心为基准共设置5个面积5 m×5 m灌木小样方和5个面积1 m×1 m的草本小样方,分别记录样地内所有乔木的种名、高度、胸径、数量和生长状况以及所有灌木和草本的种名、高度、盖度、胸径、数量和频度等。使用围径尺(精度1 mm)测量胸径,使用测高仪(精度0.5 m)测量高度,采用方格法[23]评估盖度,采用观察法描述样地基本概况,使用GPS定位仪测定样地位置信息。
1.2.2 功能性状的选取 根据TRY Plant Trait Database(https:∥www.try-db.org/TryWeb/Home.php)和中国植物图像库(http:∥ppbc.iplant.cn/),查询每个群丛中所有植物种类的性状类型,并对性状分类整理。共选取13个功能性状用于计算功能多样性指数,分别为生长型(包括乔木、灌木、草本和藤本)、物候型(包括常绿和落叶)、生活史(包括一年生、二年生和多年生)、子叶类型(包括单子叶、双子叶和蕨类)、叶质地(包括草质、革质、纸质、膜质和肉质)、叶面特征(包括有毛和无毛)、是否固氮、花期、种子传播方式(包括动物传播、水传播、自体传播和风传播)、性系统(包括两性、单性和孢子)、植株高度、胸径和盖度。
1.3 数据处理与分析
参考曾伟等[24]的重要值(IV)计算方法,乔木层重要值的计算公式为:IV=(相对密度+相对频度+相对优势度)/3;灌木层和草本层重要值的计算公式为:IV=(相对密度+相对频度+相对盖度)/3。
参考董世魁等[3]和余轩等[6]的物种多样性计算方法计算Margalef丰富度指数(R)、Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(Jsw)。
采用TWINSPAN(双向指示种分析)进行群丛分类。以本研究30个样地中所有植物的重要值作为分类的特征值,为消除异常值,排除个体数少于2的单一物种,整理得到重要值数据矩阵,将矩阵导入WinTWINS软件进行群落数量分类[25]。群丛划分参考《中国植被》[26]的群丛命名方法,同层片不同优势种或建群种用“+”连接,不同层片间用“-”连接。
根据所选择的13个功能性状,以物种为列、功能性状为行制作物种×功能性状数据矩阵,对定性性状赋值后,选用功能丰富度指数(functional richness index,FRic)、功能均匀度指数(functional evenness index,FEve)、功能离散度指数(functional dispersion index,FDis)和Rao二次熵指数(Rao’s quadratic entropy index,RaoQ)4个指标来描述功能多样性[27],并采用R 4.1.3软件的FD包计算功能多样性。
参考郑元润[28]改进的M. Godron稳定性测定方法测定群落稳定性。分别以群落物种总数的倒数累积百分比和物种出现的相对频度累积百分比分别设立横坐标和纵坐标进行曲线方程模拟。另外做1条与平滑曲线相交的直线方程y=100-x,交点越接近稳定点(20,80),群落就越稳定;相反,群落稳定性就越差[29]。采用群落交点坐标与稳定点坐标之间的欧氏距离来表示群落稳定性的大小。
所有数据均利用EXCEL 2017软件进行分类整理,图表制作利用GraphPad Prism 9完成,统计分析利用SPSS 26.0软件进行,如数据通过正态性和方差齐性检验,显著性差异采用LSD多重比较法进行分析,反之则采用Kruskal-Wallis法进行显著检验。群落稳定性、物种多样性和功能多样性间的相关性选用Pearson非参数相关性分析。
2 结果和分析
2.1 植被群落结构与物种多样性特征
调查结果显示:广东鼎湖山河岸林水翁蒲桃+蒲桃群落共有植物134种,隶属于55科98属,各植被层重要值排名前10的种类见表1,各植被层的物种多样性特征见表2。
由表1可见:从植被类型看,鼎湖山河岸林水翁蒲桃+蒲桃群落优势种多为热带亚热带地区广泛分布的植物。如乔木层中的水翁蒲桃、蒲桃、朴树、鱼尾葵、白楸〔Mallotuspaniculatus(Lam.) Muell. Arg.〕和银柴〔Aporosadioica(Roxburgh) Muller Argoviensis〕等,这些植物通常喜光,且对环境的适应性强。灌木层分布最多的优势种为苎麻〔Boehmerianivea(Linn.) Gaudich.〕,该植物根系深厚,适应力强,生长范围广,也是热带亚热带地区最常见的灌木;受乔木层植物遮光及河岸林高湿条件的影响,灌木层其他植物更多的为耐阴湿物种,如假鹰爪(DesmoschinensisLour.)、紫玉盘(UvariamacrophyllaRoxburgh)和厚叶算盘子〔Glochidionhirsutum(Roxb.) Voigt〕等。草本层多分布喜水湿植物,如菖蒲(AcoruscalamusLinn.)、芋〔Colocasiaesculenta(Linn.) Schott.〕、竹叶草〔Oplismenuscompositus(Linn.) Beauv.〕、海芋〔Alocasiaodora(Roxburgh) K. Koch〕和聚花草(FloscopascandensLour.)等。
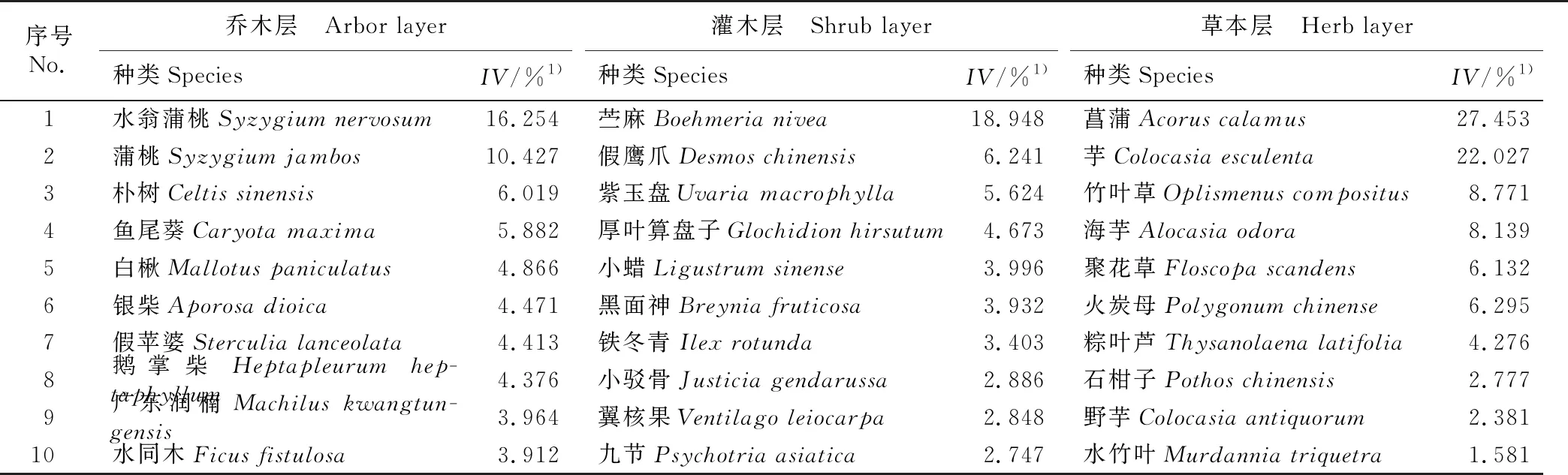
表1 广东鼎湖山河岸林水翁蒲桃+蒲桃群落各植被层重要值排名前10的种类
由表2可见:鼎湖山河岸林水翁蒲桃+蒲桃群落各植被层的Margalef丰富度指数(R)、Simpson多样性指数(D)和Shannon-Wiener多样性指数(H)由高到低依次为草本层、乔木层、灌木层,Pielou均匀度指数(Jsw)由高到低依次为灌木层、草本层、乔木层。
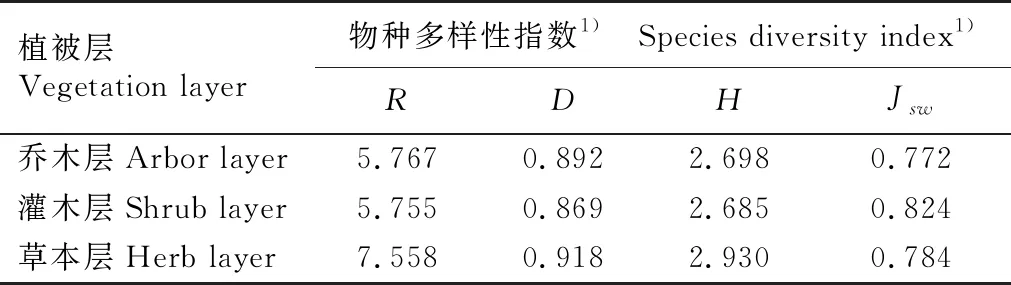
表2 广东鼎湖山河岸林水翁蒲桃+蒲桃群落各植被层的物种多样性指数
2.2 群丛类型划分结果
对广东鼎湖山河岸林水翁蒲桃+蒲桃群落30个样地所有植物的重要值矩阵进行TWINSPAN等级分类,结合研究区实际情况和群落生态意义,采用第5级划分结果,将研究区植物群落划分为8个群丛类型(图1)。由于许多群丛中没有明显的灌木层优势种或草本层优势种,因此将灌木层和草本层(以下简称为灌草层)的优势种合并统计。
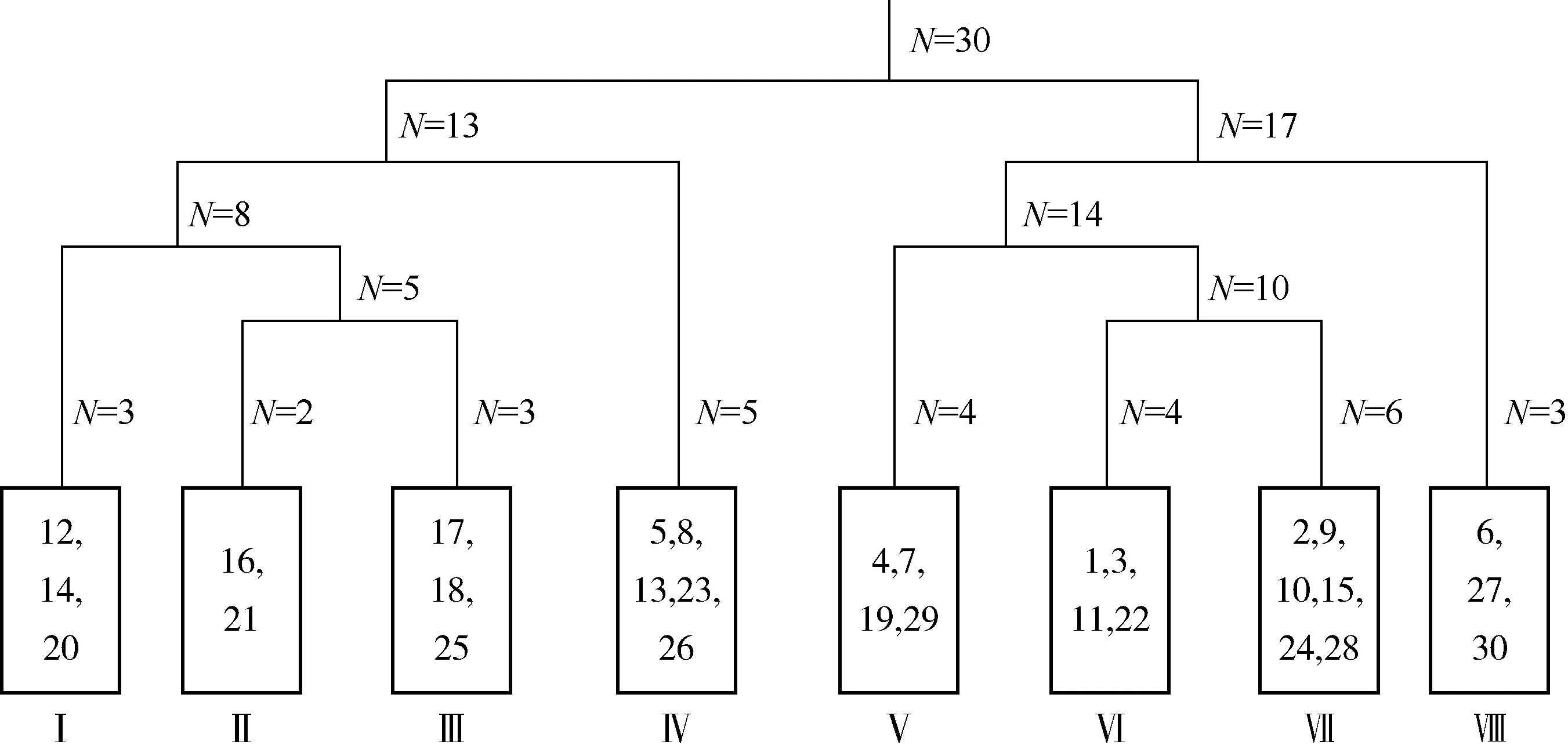
N: 样地数量Number of plots. 每个矩形框代表1个群丛,矩形框中数字代表样地序号Each rectangle box represents an association, and the numerals in the rectangular box represent the Nos. of plots. Ⅰ: 水翁蒲桃+蒲桃-聚花草群丛Ass. Syzygium nervosum+Syzygium jambos-Floscopa scandens; Ⅱ: 蒲桃+鹅掌柴-翼核果-微甘菊群丛Ass. Syzygium jambos+Heptapleurum heptaphyllum-Ventilago leiocarpa-Mikania micrantha; Ⅲ: 水翁蒲桃+水同木-小蜡群丛Ass. Syzygium nervosum+Ficus fistulosa-Ligustrum sinense; Ⅳ: 蒲桃+银柴-九节-荩草群丛Ass. Syzygium jambos+Aporosa dioica-Psychotria asiatica-Arthraxon hispidus; Ⅴ: 水翁蒲桃+广东润楠-聚花草群丛Ass. Syzygium nervosum+Machilus kwangtungensis-Floscopa scandens; Ⅵ: 水翁蒲桃+对叶榕-苎麻-海芋群丛Ass. Syzygium nervosum+Ficus hispida-Boehm erianivea-Alocasia odora; Ⅶ: 水翁蒲桃-苎麻-菖蒲群丛Ass. Syzygium nervosum-Boehm erianivea-Acorus calamus; Ⅷ: 水翁蒲桃+鱼尾葵-野芋群丛Ass. Syzygium nervosum+Caryota maxima-Colocasia antiquorum.
群丛Ⅰ:水翁蒲桃+蒲桃-聚花草群丛,该群丛包括12、14和20号样地,乔木层优势种为水翁蒲桃和蒲桃,伴生种为鹅掌柴〔Heptapleurumheptaphyllum(Linn.) Y. F. Deng〕和广东润楠(MachiluskwangtungensisYang);灌草层优势种为聚花草,伴生种为二色波罗蜜(ArtocarpusstyracifoliusPierre)、金毛狗〔Cibotiumbarometz(Linn.) J. Sm.〕和小驳骨(JusticiagendarussaN. L. Burman)等。
群丛Ⅱ:蒲桃+鹅掌柴-翼核果(VentilagoleiocarpaBenth.)-微甘菊(MikaniamicranthaH. B. K.)群丛,该群丛包括16和21号样地,乔木层优势种为蒲桃和鹅掌柴,伴生种为水翁蒲桃和白楸〔Mallotuspaniculatus(Lam.) Muell. Arg.〕;灌草层优势种为翼核果和微甘菊,伴生种为紫玉盘、华南羽节紫萁〔Plenasiumvachellii(Hook.) C. Presl〕和知风草〔Eragrostisferruginea(Thunb.) Beauv.〕。
群丛Ⅲ:水翁蒲桃+水同木(FicusfistulosaReinw. ex Bl.)-小蜡(LigustrumsinenseLour.)群丛,该群丛包括17、18和25号样地,乔木层优势种为水翁蒲桃和水同木,伴生种为鹅掌柴和广东润楠;灌草层优势种为小蜡,伴生种为铁冬青(IlexrotundaThunb.)、仙茅(CurculigoorchioidesGaertn.)和华南羽节紫萁。
群丛Ⅳ:蒲桃+银柴-九节(PsychotriaasiaticaWall.)-荩草〔Arthraxonhispidus(Thunb.) Makino〕群丛,该群丛包括5、8、13、23和26号样地,乔木层优势种为蒲桃和银柴,伴生种为华润楠〔Machiluschinensis(Champ. ex Benth.) Hemsl.〕和广东润楠等;灌草层优势种为九节和荩草,伴生种为栝楼(TrichosantheskirilowiiMaxim.)、贴生石韦〔Pyrrosiaadnascens(Sw.) Ching〕、菖蒲、栀子(GardeniajasminoidesEllis)和黑面神〔Breyniafruticosa(Linn.) Hook. f.〕等。
群丛Ⅴ:水翁蒲桃+广东润楠-聚花草群丛,该群丛包括4、7、19和29号样地,乔木层优势种为水翁蒲桃和广东润楠,伴生种为蒲桃和鱼尾葵;灌草层优势种为聚花草,伴生种为铁冬青、荩草、竹叶草、碎米莎草(CyperusiriaLinn.)、小蜡、海芋、海金沙〔Lygodiumjaponicum(Thunb.) Sw.〕和铁线蕨(Adiantumcapillus-venerisLinn.)。
群丛Ⅵ:水翁蒲桃+对叶榕(FicushispidaLinn. f.)-苎麻-海芋群丛,该群丛包括1、3、11和22号样地,乔木层优势种为水翁蒲桃和对叶榕,伴生种为阴香〔Cinnamomumburmanni(Nees et T. Nees) Blume〕等;灌草层优势种为苎麻和海芋,伴生种为白背叶〔Mallotusapelta(Lour.) Muell. Arg.〕、山葛〔Puerariamontana(Loureiro) Merrill〕和竹叶草。
群丛Ⅶ:水翁蒲桃-苎麻-菖蒲群丛,该群丛包括2、9、10、15、24和28号样地,乔木层优势种为水翁蒲桃,伴生种为对叶榕;灌草层优势种为苎麻和菖蒲,伴生种为芋和海芋等。
群丛Ⅷ:水翁蒲桃+鱼尾葵-野芋群丛,该群丛包括6、27和30号样地,乔木层优势种为水翁蒲桃和鱼尾葵,伴生种为蒲桃和鹅掌柴;灌草层优势种为野芋,伴生种为水茄(SolanumtorvumSwartz)、杖藤(CalamusrhabdocladusBurret)、海金沙、山蒟(PiperhanceiMaxim.)、娃儿藤〔Tylophoraovata(Lindl.) Hook. ex Steud.〕和铁线蕨。
2.3 群落稳定性分析结果
广东鼎湖山河岸林水翁蒲桃+蒲桃群落各群丛的稳定性分析结果见表3。由表3可见:群丛Ⅰ(水翁蒲桃+蒲桃-聚花草群丛)、Ⅱ(蒲桃+鹅掌柴-翼核果-微甘菊群丛)、Ⅲ(水翁蒲桃+水同木-小蜡群丛)、Ⅳ(蒲桃+银柴-九节-荩草群丛)、Ⅴ(水翁蒲桃+广东润楠-聚花草群丛)、Ⅵ(水翁蒲桃+对叶榕-苎麻-海芋群丛)、Ⅶ(水翁蒲桃-苎麻-菖蒲群丛)和Ⅷ(水翁蒲桃+鱼尾葵-野芋群丛)的稳定性拟合方程的R2值分别为0.999、0.995、0.983、0.972、0.998、0.947、0.976和0.992,说明该拟合曲线具有很好的拟合度,可以使用该拟合方程模拟物种总数的倒数累计百分比和物种出现的相对频度累计百分比的回归关系。上述8个群丛的稳定性拟合方程分别为y=0.000 2x3-0.037 6x2+3.915 0x+8.425 0、y=0.000 2x3-0.043 6x2+3.049 0x-16.830 0、y=-0.008 5x2+1.531 0x+28.950 0、y=0.000 1x3-0.027 4x2+2.149 0x+29.630 0、y=0.000 1x3-0.025 5x2+2.113 0x+28.440 0、y=0.000 2x3-0.040 1x2+2.727 0x+28.770 0、y=-0.008 4x2+1.463 0x+34.810 0和y=0.000 2x3-0.043 3x2+3.194 0x+10.240 0。除了群丛Ⅵ外,鼎湖山河岸林水翁蒲桃+蒲桃群落中其他群丛的稳定性较低,群丛稳定性由高到低依次为群丛Ⅵ、群丛Ⅱ、群丛Ⅳ、群丛Ⅴ、群丛Ⅷ、群丛Ⅶ、群丛Ⅰ、群丛Ⅲ。
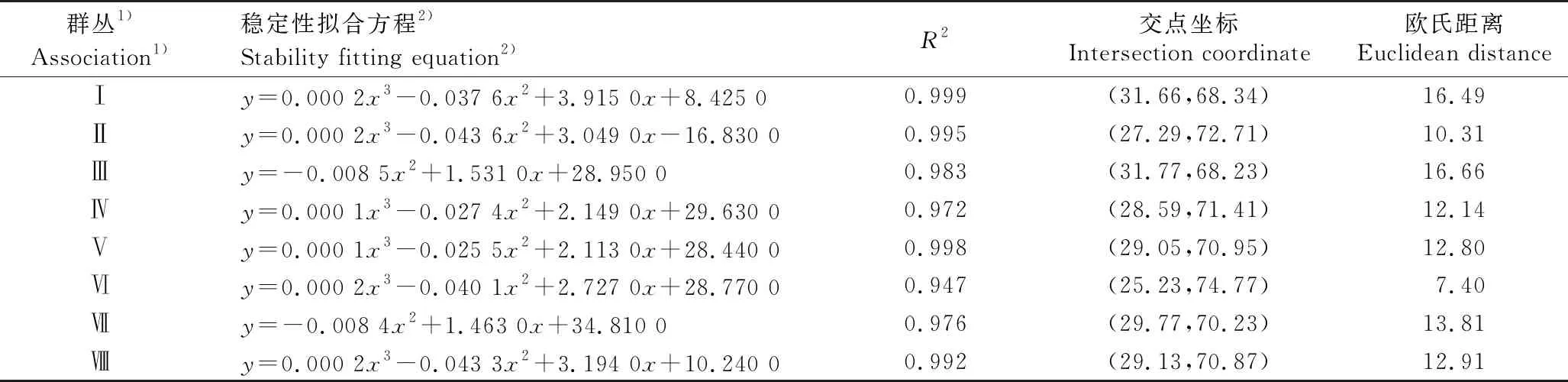
表3 广东鼎湖山河岸林水翁蒲桃+蒲桃群落各群丛的稳定性分析结果
2.4 群落多样性特征及其与群落稳定性之间的关系
广东鼎湖山河岸林水翁蒲桃+蒲桃群落各群丛物种多样性指数和功能多样性指数见表4。由表4可见:鼎湖山河岸林水翁蒲桃+蒲桃群落各群丛物种多样性指数中,除Pielou均匀度指数(Jsw)的差异较小,各群丛的Margalef丰富度指数(R)、Simpson多样性指数(D)和Shannon-Wiener多样性指数(H)均存在一定差异,如群丛Ⅱ(蒲桃+鹅掌柴-翼核果-微甘菊群丛)的R值显著(P<0.05)高于群丛Ⅰ(水翁蒲桃+蒲桃-聚花草群丛)、Ⅲ(水翁蒲桃+水同木-小蜡群丛)、Ⅴ(水翁蒲桃+广东润楠-聚花草群丛)和Ⅷ(水翁蒲桃+鱼尾葵-野芋群丛),群丛Ⅱ的D值和H值显著高于群丛Ⅰ。从功能多样性指数看,除群丛Ⅲ的功能丰富度指数(FRic)显著低于其他群丛外,其他群丛间的FRic值无显著差异,且均较高,如群丛Ⅳ(蒲桃+银柴-九节-荩草群丛)和Ⅵ(水翁蒲桃+对叶榕-苎麻-海芋群丛);各群丛间的功能均匀度指数(FEve)、功能离散度指数(FDis)和Rao二次熵指数(RaoQ)均无显著差异。
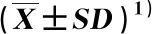
表4 广东鼎湖山河岸林水翁蒲桃+蒲桃群落各群丛物种多样性指数和功能多样性指数
广东鼎湖山河岸林水翁蒲桃+蒲桃群落稳定性、物种多样性指数和功能多样性指数间的Pearson相关系数见表5。由表5可见:R值和D值分别与H值间呈显著(P<0.05)和极显著(P<0.01)正相关;群落稳定性与R值、D值、H值和Jsw值间无显著相关性,但与FRic值和RaoQ值间均呈显著正相关;R值和D值与FRic值间呈显著正相关,而Jsw值与FRic值间呈显著负相关;D值和H值与FDis值间分别呈极显著和显著正相关;D值与RaoQ值间呈显著正相关;FRic值和FDis值与RaoQ值间分别呈显著和极显著正相关。
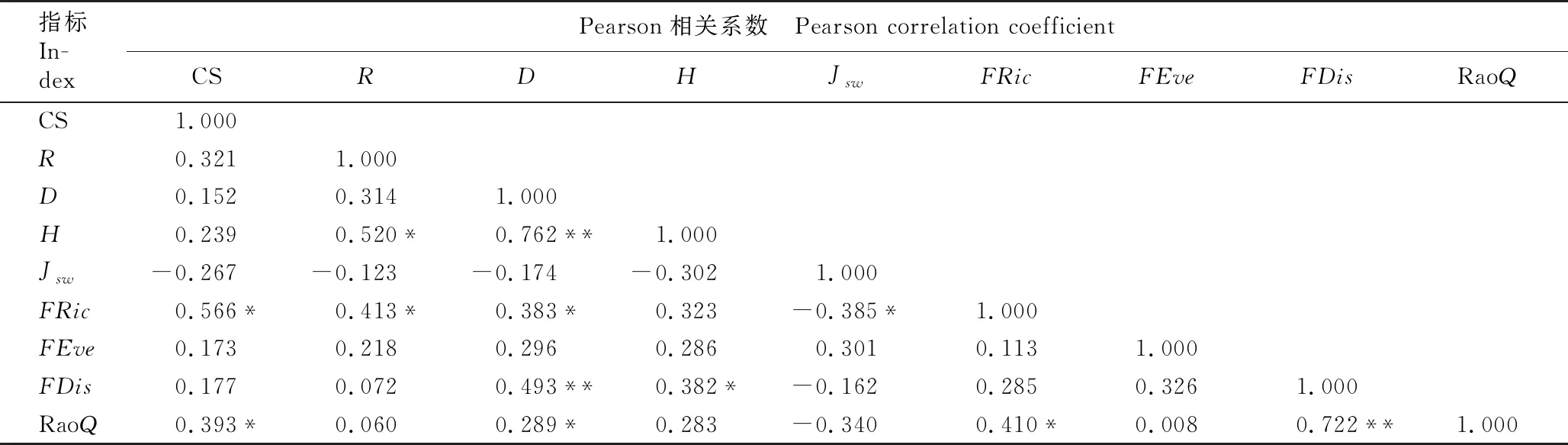
表5 广东鼎湖山河岸林水翁蒲桃+蒲桃群落稳定性、物种多样性指数和功能多样性指数间的Pearson相关系数1)
3 讨论和结论
3.1 群丛分类与群落稳定性的关系
群落稳定性是生态系统稳定性的一个层面,可以定义为群落受干扰时抵抗外界干扰的能力和系统受扰动后维持其原始结构或状态的能力,包括现状稳定、时间过程稳定和恢复力稳定等,由于概念差异和方法的不同,关于稳定性的维持机制仍很复杂[30]。通常,群落的稳定性与自我演替有关,趋于顶极演替的群落,往往具有更高的群落稳定性,但演替过程中也可能受种内和种间差异以及环境或人为干扰等因子的综合影响[17]。
本研究中,广东鼎湖山河岸林水翁蒲桃+蒲桃群落可划分为8个群丛,不同群丛的稳定性存在差异,这可能是由于每个群丛的物种组成和结构不同,导致物种的分布或功能群的类型不同,进而造成各群丛间的稳定性差异。除群丛Ⅵ(水翁蒲桃+对叶榕-苎麻-海芋群丛)外,其他群丛偏离稳定点(20,80)较远,稳定性均较低。这说明鼎湖山河岸林水翁蒲桃+蒲桃群落总体上尚未发育到成熟阶段,物种组成和群落结构尚处于不稳定的演替阶段,仍需要长时间演替才能发育为顶极群落[17]。这一方面可能是由于南方热带气候带来的极端天气(如暴雨或台风过后的径流冲刷)对鼎湖山河岸林的影响,导致靠近河岸的灌草丛死亡,同时加速了老弱病残树的死亡,使得部分新物种开始繁衍,但在快速的生长演替过程中,物种之间处于动态发育过程,各植被层或物种之间的平衡尚未建立,整体的群落结构不够稳定,因此群落的稳定性较弱[31]。另一方面可能是由于在群落的动态发育过程中,物种之间的竞争较激烈,使得竞争双方或一方的生长发育受到限制,影响整体群落的天然更新、多样性和结构等,导致群落总体的稳定性降低[32]。从各群丛结构上看,群落结构主要为乔木-草本结构和乔木-灌木-草本结构。其中以群丛Ⅵ、Ⅱ(蒲桃+鹅掌柴-翼核果-微甘菊群丛)和Ⅳ(蒲桃+银柴-九节-荩草群丛)的植物配置组合稳定性较高,均为乔木-灌木-草本结构;其次为群丛Ⅴ(水翁蒲桃+广东润楠-聚花草群丛)和Ⅷ(水翁蒲桃+鱼尾葵-野芋群丛),二者的稳定性接近,均为乔木-草本结构;其他群丛的稳定性由高到低依次为群丛Ⅶ(水翁蒲桃-苎麻-菖蒲群丛)、群丛Ⅰ(水翁蒲桃+蒲桃-聚花草群丛)、群丛Ⅲ(水翁蒲桃+水同木-小蜡群丛)。上述研究结果表明:乔木-灌木-草本结构的群落稳定性相比于乔木-草本结构的群落稳定性偏高,这可能与建群种或不同层片的优势种有关。优势种对提升群落稳定性具有促进作用,优势种可以决定种间相互作用的强度,进而影响稳定性波动[33-35]。例如:群丛Ⅵ和Ⅱ乔木层中的水翁蒲桃和蒲桃喜光性较强,且植物长势高大,对叶榕和鹅掌柴植株相对较小且耐阴,由于生物量的不同或对环境需求的不同,这些植物间可能存在偏利共生或互利共生形成的种间正联结,从而使群丛趋于完善和稳定[32]。此外,乔木-草本结构的群丛稳定性虽相对较低,但伴生种越多的群丛,稳定性明显偏高,如群丛Ⅴ的伴生种明显多于群丛Ⅰ。伴生种虽处于非优势地位,但其与优势种彼此伴生,在群落中与其他物种有着广泛联系,能够构成种间结合,形成不同的群落层片,对稳定性起到一定巩固作用。因此,对于结构合理和种群稳定的复层群落除了考虑物种多样性,更应考虑结构和不同层片数量的合理搭配,但由于稳定性的测定是以群落整体物种数和多度为计算基础,而稳定性又受到外界环境干扰,因此减少外界环境的各种干扰同样是维持群落稳定性的关键。
3.2 物种和功能多样性与群落稳定性的关系
从物种多样性或功能多样性的角度研究群落稳定性是森林生态系统研究的重要内容[3,36]。从物种多样性本身出发,物种多样性包括物种丰富度或特定区域内物种的数量以及物种均匀度或群落中物种生物量的分布2个方面[37]。而功能多样性指特定群落中植物功能性状值的大小、范围与分布状况,一般认为,物种多样性越复杂,功能多样性就越高[14]。本研究中,广东鼎湖山河岸林水翁蒲桃+蒲桃群落草本层的物种多样性高于乔木层,这是由于草本层的伴生种明显多于乔木层,且草本层多为耐受性植物,种子多且繁殖能力强,对环境的适应程度高[38],导致其物种多样性更高。此外,各群丛的物种多样性指数〔Margalef丰富度指数(R)、Simpson多样性指数(D)、Shannon-Wiener多样性指数(H)和Pielou均匀度指数(Jsw)〕均存在一定差异,一方面可能受群落内部物种组成或结构的相互影响,如物种类型或物种数的不同所形成的竞争差异影响,另一方面可能受热带性台风或暴雨的影响,径流冲刷了部分河岸,使沿岸物种及其分布受到影响。
相关性结果显示:广东鼎湖山河岸林水翁蒲桃+蒲桃群落的R值与功能丰富度指数(FRic)间呈显著(P<0.05)正相关,表明物种功能随物种数的增加而丰富。由于群落中的每个物种对功能的贡献都是独特的,物种多样性下降或物种丧失会引起功能性状丢失,导致功能多样性下降,这与多数研究结果[16,30]一致。多样性-稳定性假说认为较高的群落多样性有助于增强群落或生态系统稳定性,群落的稳定性一定程度上依靠物种数量,但群落物种或物种功能群的影响未被考虑到[8,39]。本研究中,物种多样性指数与群落稳定性间并无显著相关关系,这与郑世群等[4]、史浩伯等[10]和薛晨阳等[40]对多样性-稳定性的研究结果一致,表明群落稳定性没有受到物种数量及物种分布情况的影响。这种情况一方面可能是由于多样性增加群落稳定性的主要决定因子在于物种或物种功能群的能力,而不是多样性本身[12],而且由于功能多样性具有保险效应(功能冗余),即维持生态系统正常功能所需的最低下限,那么当某一生态系统的物种数目达到饱和后,即使物种再丰富,群落稳定性也不受物种多样性的影响,因此能够使群落稳定性保持在一定水平[1,16,41];另一方面可能是由于某些群落受到外界因子的干扰,使得物种应对干扰的能力减弱,而且物种间存在对外界响应的不同步性,即使物种功能群丰富,也难以提升群落的稳定性[10,42-43]。此外,水翁蒲桃+蒲桃群落稳定性与FRic值和Rao二次熵指数(RaoQ)间呈显著正相关,表明功能丰富度较高的群落,稳定性也会相对较高。这可能是由于功能多样性的互补效应,即通过种间功能的差异互补性增加了对外界资源的利用,因此在一定程度对群落稳定性具有促进作用[7]。另外,水翁蒲桃+蒲桃群落稳定性与功能多样性指数的相关性总体上高于物种多样性指数,说明功能多样性与群落稳定性间的关系强于物种多样性与群落稳定性间的关系。通常,RaoQ值表示物种及物种性状间的差异,当RaoQ值较高时,植物性状空间的集中程度越低,性状属性分布越分散且相对独立,因此稳定性也会降低[16,44]。但本研究中RaoQ值与群落稳定性呈显著正相关,可能是由于某些RaoQ值高的群丛物种分布均匀,且功能多样性相对高,加强了种间互补程度,使得群丛对外界资源利用差异变大,更有利于物种间合理利用资源进而促进稳定性。
综上可知,广东鼎湖山河岸林水翁蒲桃+蒲桃群落物种多样性与群落稳定性间无显著相关,但功能多样性与群落稳定性间相关性较强,群落的稳定性很大程度上取决于植物群落内部结构和物种功能对干扰的抵抗能力和自我修复能力。但造成这一现象的原因不仅与物种本身或其特征属性有关,还可能受外界干扰强度、种群变异性、群落生产力、生境异质性、竞争和扩散限制等影响[45-46]。因此,对于稳定性的衡量或多样性-稳定性的关系,仍然无法用单一因子解释,稳定性变化更有可能是多样性同其他因子综合作用的结果。
猜你喜欢
辽河(2024年1期)2024-03-04 03:01:57
中国水土保持科学(2023年6期)2024-01-05 05:53:12
水土保持研究(2022年1期)2022-12-27 07:35:24
花火彩版A(2021年10期)2021-01-03 14:11:10
广西植物(2019年2期)2019-09-10 07:22:44
天然产物研究与开发(2018年11期)2018-11-30 02:06:38
广东第二课堂·初中(2017年2期)2017-03-15 19:57:26
湖南大学学报(社会科学版)(2016年5期)2016-01-03 07:20:39
湖南大学学报(社会科学版)(2016年5期)2016-01-03 07:20:39
商业经济研究(2014年17期)2014-06-20 07:13:51
