
氮磷添加对呼伦贝尔草地凋落物分解的影响
2023-02-28 08:36:18杜鹏冲潘昱臻侯双利王智慧王洪义
草业学报 2023年2期
杜鹏冲,潘昱臻,侯双利,王智慧,王洪义*
(1.黑龙江八一农垦大学园艺园林学院,黑龙江 大庆 163319;2.中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统研究站,辽宁 沈阳 110016)
凋落物是草地生态系统能量流动和物质循环的重要组成部分,对维持草地生态系统服务功能具有重要作用[1]。陆地生态系统中约50%的净初级生产力以凋落物的形式返回土壤,成为土壤有机质的主要来源[2]。凋落物分解作为养分和碳(carbon,C)循环的关键环节,推动着土壤有机质矿化分解和营养元素的循环转化,是土壤和植物之间相互作用的重要纽带,对全球C循环具有重要意义[3-4]。凋落物分解是一系列物理和化学过程作用的结果,包括淋洗作用、机械破碎、土壤动物及微生物分解等[5-6]。凋落物分解受到内部因素和外部因素共同影响,其中内部因素包括纤维素、木质素、氮(nitrogen,N)、磷(phosphorus,P)含量及其比值等[7],外部因素包括气候、土壤理化性质及土壤微生物生物量等[8-9]。
养分是草地生态系统净初级生产力的主要限制因素之一。养分添加可以增加土壤肥力,促进退化草地恢复,以及提高净初级生产力[10]。畜牧业发达国家通常将施肥作为草原管理的重要措施[11]。氮(N)和磷(P)作为植物生长所必需的大量营养元素,是植物生长发育、群落动态、生态系统结构与功能的重要调控者,是北方草地生态系统的主要限制因子[1,12-13]。N、P养分添加对凋落物分解的影响存在较多争议[14]:有些研究认为养分增加能够促进凋落物的分解[15],尤其N素添加,较具有普遍性[3,16],而P素添加开展较少,多数情况下没有N添加作用明显;而有些研究发现养分添加对凋落物分解无显著作用[17]或有抑制作用[18]。其中植物种类及试验位点生境差异是产生不同影响结果的重要原因。关于养分添加对凋落物分解的影响研究主要有两种类型试验,一种是天然草地的凋落物放入养分添加处理的样地做模拟分解试验;另一种是养分添加处理的凋落物在本样地做原位分解试验。两种情况还不完全一样,第一种是只考虑环境因子变化的作用;第二种是考虑了植物和环境,但是无法区分二者的相对作用。因此,为厘清内在和外在因素对凋落物分解的影响程度,有必要同时开展植物自身养分和土壤养分差异对凋落物分解影响的研究。
为解决以上问题,本研究依托中国科学院沈阳应用生态研究所额尔古纳站氮磷添加控制实验平台,以羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)和披针叶黄华(Thermopsis lanceolata)3种不同养分利用方式物种为研究对象,设置了样方内分解和样方外分解两个对比试验。研究N、P添加对呼伦贝尔草地凋落物分解的影响以及分析植物内在养分因素和土壤养分因素对凋落物分解的影响程度。
1 材料与方法
1.1 研究区概况
研究地点位于内蒙古自治区呼伦贝尔市额尔古纳市境内,依托中国科学院沈阳应用生态研究所额尔古纳森林草原过渡带生态系统研究站(50°10′46″N,119°22′56″E)。该地区属于温带大陆性季风气候,冬季寒冷,夏季高温;年平均温度-2.5℃;年均降水量360 mm[19];土壤类型为黑钙土;地带性植被类型为草甸草原,植被组成以禾本科、菊科、豆科、蔷薇科植物为主,羊草和贝加尔针茅(Stipa baicalensis)为优势种,常见种有糙隐子草、披针叶黄华、寸草苔(Carex duriuscula)、达乌里芯芭(Cymbaria dahurica)和阿尔泰狗娃花(Heteropappus altaicus)等[19]。
1.2 试验设计
本试验在额尔古纳站氮磷添加控制实验平台开展,试验处理分别为对照(CK)、加氮(硝酸铵,NH4NO3,10 gN·m-2·yr-1)、加磷(磷酸二氢钾,KH2PO4,10 g P·m-2·yr-1)、氮磷共同添加(10 g N·m-2·yr-1、10 g P·m-2·yr-1),每个处理5次重复,一共20个样方。从2014年开始,在每年5月下旬植物返青时进行施肥,氮、磷肥料均为干施形式,分别在下雨前或傍晚前人工撒施,以利于肥料快速溶解。
1.3 样品采集、处理及实验测定
2015年9月生长季节结束后,在自然草地和氮磷添加控制实验平台分别收集3种植物枯萎的地上植株部分,样品放置分为样方内和样方外两部分:1)样方内处理:从自然草地选取羊草、糙隐子草和披针叶黄华3种植物的凋落物放于氮磷添加控制实验平台样方内进行分解试验;2)样方外处理:从上述氮磷添加平台对应的4个处理中选取3种相同植物凋落物,放于自然草地(样方外)进行分解试验。将收集的每种凋落物风干后剪成长度为10 cm的条段状,单独放入孔径为1 mm,大小为15 cm×20 cm的尼龙网袋中,其中羊草8 g,糙隐子草5 g,披针叶黄华5 g,每种植物凋落物放置4个网袋。凋落物袋放置于地表平铺,四角固定,并保证与地面充分接触。
野外凋落物分解网袋分别于2016年9月和2017年9月两次收回,每次每个物种取2袋,取回的样品剔除杂质,将凋落物放入信封,在烘箱中65℃烘48 h,称重,粉碎后用于元素分析。采用重铬酸钾氧化-比色法测定有机碳含量;采用浓H2SO4-H2O2消煮,然后用全自动间断化学分析仪(Cleverchem 200+,De Chem-Tech Gmb H,德国)测定全N和全P含量[20]。
1.4 数据分析
凋落物分解速率用k表示,采用负指数衰减模型Xt/X0=e-kt,其中Xt代表凋落物在分解一段时间(t)后的残留量,X0表示凋落物未分解时的初始重量,k为凋落物分解系数,t为凋落物的分解时间,以年表示。元素残留率用R表示,R=[(Ct×Xt)/(C0×X0)]×100%,Ct表示t时刻凋落物元素含量(mg·g-1),Xt表示t时刻凋落物的干重(g),C0表示初始元素含量(mg·g-1),X0为初始干重(g)。凋落物元素释放率用NR表示,NR=100-R,其中R为元素残留率。用WPS 2018进行数据整理,用SPSS 24进行方差分析,采用单因素方差分析(one-way ANOVA)比较不同处理间差异显著性(显著性水平为P<0.05,Duncan检验);用Graphpad-prism 7软件进行制图。
2 结果与分析
2.1 凋落物初始化学成分分析
对羊草、糙隐子草以及披针叶黄华凋落物初始化学成分分析表明(表1),3种植物初始化学元素组成存在一定的差异。凋落物中的C元素含量为羊草>披针叶黄华>糙隐子草。3种植物的C/N为羊草>糙隐子草>披针叶黄华;4种处理的C/N为P>CK>NP>N。披针叶黄华作为豆科植物的代表,其特有的固氮能力,导致其初始N元素含量远远高于羊草与糙隐子草。
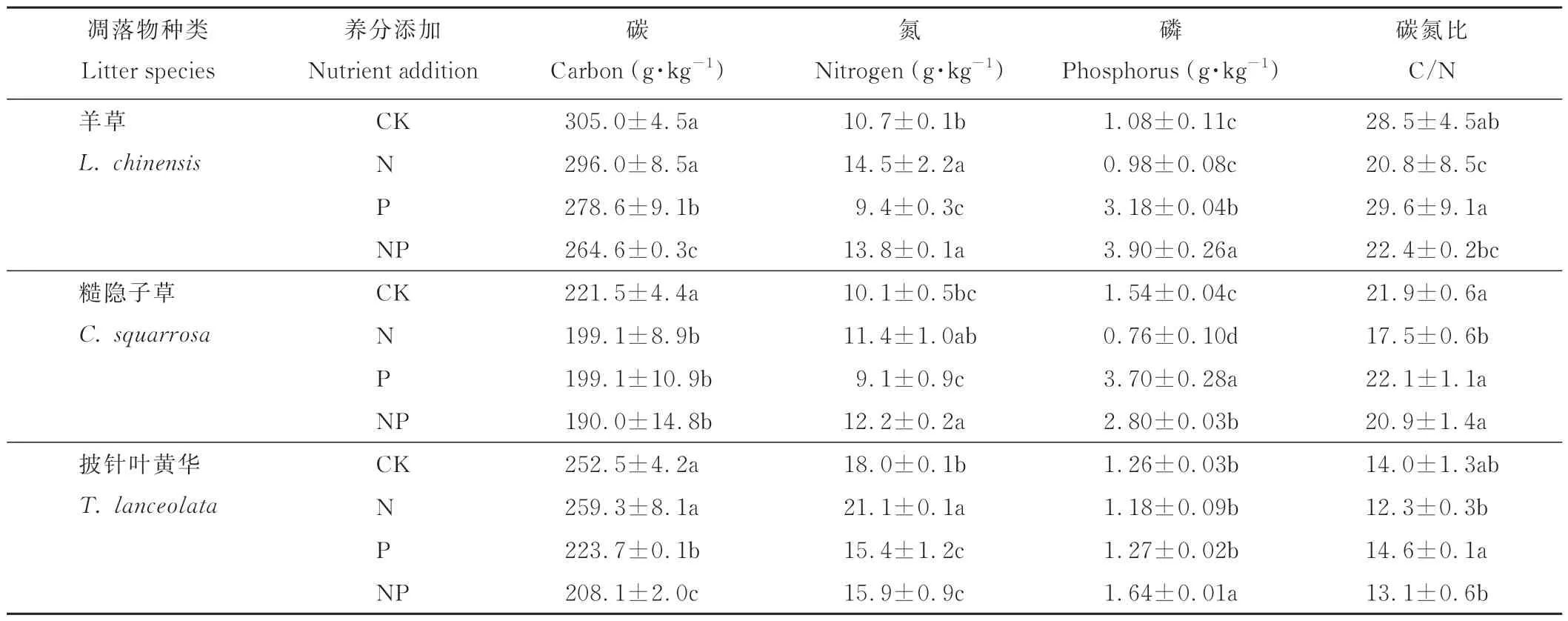
表1 各凋落物初始化学元素组成Table 1 The initial chemical elements of each litter
2.2 氮磷添加对凋落物重量变化的影响
在凋落物分解过程中,凋落物重量变化可衡量凋落物分解速度快慢。研究表明:4种处理下,各凋落物干重残余量均呈持续下降趋势,且前期分解速度快于后期(图1)。羊草与糙隐子草经过一年的分解,样方内与样方外的凋落物残余量无明显差异;分解两年后,样方内凋落物残余量低于样方外,其中羊草约降低9.3%,糙隐子草约降低8.4%。披针叶黄华第一年分解,样方外的凋落物残余量低于样方内,分解两年后,样方内与样方外的凋落物残余量无明显差异。样方内养分添加下,凋落物残余量显著低于CK处理,样方外各处理之间无明显规律。
2.3 凋落物分解速率模型
由表2可知,各植物对N、P、NP添加的响应不同,3种植物的半分解时间及分解95%时间都有所减少(羊草N素添加处理除外),皆表现出前期分解较快,后期分解较慢的趋势。3种凋落物平均分解速率为披针叶黄华>糙隐子草>羊草(图1)。样方内,羊草的分解系数为0.303~0.403,凋落物在NP添加下的分解周期最短,根据Olson分解模型分解95%时间排序为CK>N>P>NP;糙隐子草的分解系数为0.412~0.487,凋落物在NP添加下的分解周期最短,根据Olson分解模型分解95%时间排序为CK>N>P>NP;披针叶黄华的分解系数为0.427~0.533,在NP添加下的分解周期最短,根据Olson分解模型分解95%时间排序为CK>N>P>NP。3种植物皆表现出CK处理分解周期最长,NP添加处理分解周期最短。
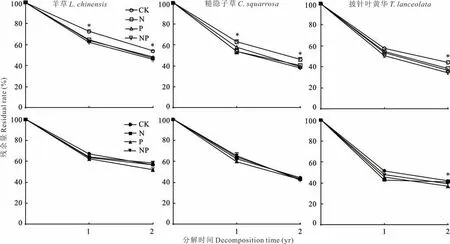
图1 养分添加下3种植物凋落物残余量变化Fig.1 Changes of litter residual rate of three plants under nutrient addition
样方外,羊草的分解系数为0.288~0.321,P添加处理分解周期最短,根据Olson分解模型分解95%时间排序为N>NP>CK>P。糙隐子草分解系数为0.405~0.432,NP处理分解周期最短,根据Olson分解模型分解95%时间排序为CK>N>P>NP。披针叶黄华分解系数为0.433~0.499,P添加处理分解周期最短,根据Olson分解模型分解95%时间排序为CK>N>NP>P(表2)。

表2 3种植物凋落物残留率随时间的指数方程Table 2 Exponential equation of litter residual rate of three plants with time
总体来说,外因方面,养分添加促进了凋落物分解;内因方面,养分添加对凋落物分解的影响因物种而异。
2.4 氮磷添加对凋落物C、N、P释放的影响
有机碳的变化可以从侧面反映凋落物的分解情况。图2表明,样方内,3种植物有机碳残留率与凋落物残留率变化趋势相同,皆呈下降趋势,养分添加处理与CK呈显著差异。样方外,3种凋落物有机碳残留量呈持续下降趋势,分解初期,NP添加处理有机碳残余量高于其他处理。羊草与糙隐子草分解2年后,样方内有机碳残余量低于样方外有机碳残余量,而披针叶黄华样方内外有机碳残余量基本一致。

图2 氮磷添加对有机碳残余量的影响Fig.2 Effect of nitrogen and phosphorus addition on organic carbon residual rate
由图3可知,2年分解过程中,样方内养分添加处理氮元素释放率高于样方外。3种植物均表现为N和NP添加处理氮元素释放率高于CK处理。羊草CK处理氮元素释放不同于其他处理,表现为富集-释放。

图3 氮磷添加对氮素释放率的影响Fig.3 Effect of nitrogen and phosphorus addition on nitrogen release rate
由图4可知,3种植物样方内外均表现为P元素持续释放,且前期释放快,后期释放慢。羊草与糙隐子草P元素释放表现为P、NP添加处理显著高于CK和N添加处理;而披针叶黄华表现为P、NP添加处理显著低于CK和N添加处理。

图4 氮磷添加对磷素释放率的影响Fig.4 Effect of nitrogen and phosphorus addition on phosphorus release rate
3 讨论
3.1 养分添加在外因和内因方面对凋落物分解速率的影响
影响凋落物分解的主要因素有外在因素和内在因素,厘清两者的影响程度,有助于今后更好地调控凋落物分解[21]。本研究中样方内分解试验表明,作为外在因素,养分添加可促进3种凋落物的分解,该结果与李文亚等[22]在草地开展的N、P添加对贝加尔针茅、羊草和冷蒿(Artemisia frigida)影响的研究结果一致;在林地开展的N、P添加对松林凋落物叶分解的研究结果也得出相同结论[23]。养分添加促进凋落物分解,主要是因为土壤养分增加改变土壤微生物的群落结构组成,增加微生物活性[24]和土壤酶活性[25],从而促进了凋落物分解。
一般研究认为,植物自身初始养分含量会影响凋落物分解,养分增加能促进以C、N、P为底物的凋落物分解酶的活性,进而提高了凋落物分解速率[26-27]。本研究中样方外分解试验表明,作为内因的养分添加对3种凋落物分解的影响不同。糙隐子草和披针叶黄华初始N、P含量较高的处理凋落物的分解较快,而羊草初始含N量较高的处理凋落物分解较慢。3种植物的不同响应方式与文海燕等[28]在黄土高原典型草原开展的N添加试验结果相似,其研究表明只有长芒草(Stipa bungeana)凋落物初始N含量与其分解速率显著正相关,而阿尔泰狗娃花的分解速率与初始养分没有显著相关性。物种间差异化响应与物种自身属性有关[29],如羊草具有较高难分解的木质素和纤维素[30],而糙隐子草木质素含量较低[31],披针叶黄华是含氮量高的豆科植物,这些均能影响凋落物分解速率。另外,C/N在很多生态系统中被证明是影响凋落物分解的主要因素[25,32],一般C/N较低的凋落物分解较快。本研究中糙隐子草和披针叶黄华初始C/N较低,平均分别为20.6和13.5,而羊草初始C/N较高,平均为25.3,因此,在分解环境相同的情况下,物种间的初始C/N可能也是影响其凋落物分解的重要因素[32-33]。
3.2 N、P以及NP添加对凋落物分解的影响
根据Olson分解模型,N、P和NP添加对凋落物半分解及分解95%时间影响不同,表现为促进或抑制作用。样方内分解条件下,3种植物均表现为CK处理分解时间最长,NP添加处理分解时间最短,N、P单独添加也促进了凋落物分解。其原因是呼伦贝尔草地主要受N限制,其次是P限制[34],养分添加提高了植物生产力,增加了土壤微生物、土壤酶、土壤动物的活性[9],尤其是NP同时添加,草地的主要养分限制被解除,物质和能量间流动更加畅通,能最大限度提高分解者的活性,进而提高了凋落物的分解速率。
与样方内不同,样方外3种凋落物对N、P和NP添加响应不同。羊草在含N量较高的N和NP添加处理下,凋落物分解95%时间较CK长,表现为抑制作用,该结果与Berg[35]的研究结果相似,其研究发现,N添加尤其在后期可明显降低凋落物分解速率,其原因是凋落物分解过程中较高的N素会与纤维素、单宁、木质素分解产生的次生代谢物质发生聚合反应[36],形成不易分解的物质,进而降低凋落物的分解速率;除此之外,初始N含量较高还会抑制木质素分解酶和胞外酶活性,也会影响木质素含量较高的凋落物分解[37]。与羊草不同,糙隐子草和披针叶黄华在初始N含量较高的N和NP添加处理中,分解95%时间均小于CK处理。3种植物均表现为P添加处理分解时间最短,其原因可能是P添加可提高磷酸酶活性,促进水溶性碳水化合物分解[38],进而提高了凋落物分解速率;另外,P添加处理能够提高3种凋落物中P素含量,可以为微生物分解者提供更多的P养分,因此可能缓解了凋落物分解过程中P素的限制,进而有利于凋落物的分解。
3.3 养分添加对凋落物元素释放的影响
有机碳是凋落物的重要组成物质,2年的分解过程中,样方内外均表现持续释放模式,与凋落物残留率变化趋势基本一致,该结果与李文亚等[22]在呼伦贝尔草地的研究结果一致,C作为植物主要的结构物质,早期分解主要是淋溶过程[39],随着分解进行,C/N降低,凋落物中耐分解的有机化合物逐渐分解,表现为持续的C释放过程[40-41]。本研究样方内凋落物N元素快于样方外N元素释放,其原因主要是养分添加作为外因更有利于凋落物分解,凋落物分解过程中N素作为底物,随分解时间逐渐释放[42]。N和NP添加处理,凋落物N素释放率较高,一方面是N素添加促进凋落物分解,另一方面是植物自身N素含量高,也能促进N素释放[43]。羊草的CK处理样方内外均表现为富集-释放,这种释放模式主要是凋落物初始N素含量低,微生物占领凋落物过程中将土壤中N素转移到凋落物上[13],后期随着微生物活性增加,凋落物分解逐渐加快,表现为先富集后释放。羊草和糙隐子草P元素的释放与N元素释放相似,P和NP添加处理下凋落物P素含量更高且释放速度更快。然而,P素添加没有加速披针叶黄华磷的释放,其原因可能是披针叶黄华为豆科植物,自身固氮较多,导致披针叶黄华凋落物含有较高的N含量和N∶P,因此,相比于N素,微生物在分解过程中更难以从其凋落物本身获取P素。
4 结论
2年的分解试验显示,样方内凋落物分解速率较样方外快,说明养分添加在外因方面作用大于内因方面。外因方面,养分添加促进3种凋落物分解;内因方面,因物种不同,其中对糙隐子草和披针叶黄华有促进作用,而对羊草则表现为抑制作用。从凋落物半分解及分解95%时间角度衡量,3种植物分解速率为披针叶黄华>糙隐子草>羊草。C作为植物主要的结构物质,其释放与凋落物残留率变化趋势基本一致。N和NP添加处理,均能促进3种凋落物N素释放,表现为样方内处理释放速率大于样方外处理。P和NP添加处理均能促进羊草和糙隐子草凋落物P素释放,但不能提高披针叶黄华P素释放。凋落物的元素释放是物质循环的重要过程,该过程因物种、分解阶段、环境条件不同而异,其释放机理还有待进一步研究。本研究揭示了养分添加下,外因作用是促进凋落物分解和养分释放的主导因素,后续研究应更多关注于外界因素变化对凋落物分解过程的影响。
猜你喜欢
小学阅读指南·教研版(2022年3期)2022-04-28 22:20:30
现代畜牧科技(2021年7期)2021-07-28 06:40:30
现代畜牧科技(2021年4期)2021-07-21 06:13:04
林业科学(2020年10期)2020-11-30 00:36:56
作文周刊·小学四年级版(2019年8期)2019-04-23 01:04:16
国际木业(2016年6期)2016-02-28 10:27:03
国际木业(2016年5期)2016-02-28 10:26:34
现代畜牧科技(2016年9期)2016-02-21 12:25:04
湖南农业科学(2014年1期)2014-02-27 14:25:26
当代畜禽养殖业(2010年4期)2010-08-15 00:49:57
