
木薯MeHsfB3a基因的克隆及其抗细菌性枯萎病功能鉴定
2023-02-27 03:08李琳琳王超群李春霞王红刚陈银华张肖飞耿梦婷
热带作物学报 2023年1期
李琳琳,王超群,李春霞,骆 凯,王红刚,陈银华,张肖飞,耿梦婷*
1.海南大学热带作物学院,海南海口 570228;2.国际热带农业中心,哥伦比亚卡利 AA 6713
植物在生长发育过程中面临生物胁迫和非生物胁迫,为了应对胁迫以维持正常生长,植物进化出了许多抵御逆境的机制[1]。其中热激转录因子(heat stress transcription factors, Hsfs)是该防御体系中响应逆境信号的重要元件[2]。研究表明Hsfs参与了高温、干旱、盐胁迫和氧化损伤等逆境的调控,在植物抵御非生物胁迫过程中有重要作用[3-4]。在拟南芥中过量表达大白菜BrHsf16基因,可明显提高转基因拟南芥植株在热胁迫下的存活率[5]。沉默辣椒CaHsfa1d基因,导致植株耐热性降低[6]。过量表达热激转录因子GmHsFA1的大豆植株在受到干旱逆境胁迫下,脯氨酸、可溶性糖含量明显增高,植株的抗干旱性增强[7]。拟南芥AtHsfA7b可调控下游抗逆相关靶基因表达响应盐胁迫,通过降低细胞失水率、调节渗透势来提高植株耐盐性[8]。用H2O2处理水稻种子,发现水稻中多个OsHsfs表达量增加,在抵抗氧化胁迫等方面发挥重要作用[9]。同时,Hsfs在作物响应病害胁迫过程中也起着重要作用。水稻白叶枯病是由黄单胞杆菌(Xanthomonas oryzaepv.oryzae,Xoo)引起的病害,分别对水稻抗病品种‘SH5’和感病品种‘8411’接种黄单胞杆菌,6 h后OsHsfA2a、OsHsfA2b、OsHsfB2a、OsHsfB2b、OsHsfB2c、OsHsfC1b和OsHsfC2c表达量显著上调,OsHsfA2c和OsHsfB4c表达量显著下调,这说明Hsfs在抵御病害侵染时发挥了作用[10]。过表达AtHsfA1b的拟南芥转基因植株在接种病原菌Pto DC3000进行抗性实验证明,过表达植株的细菌数目显著低于野生型,AtHsfA1b正调控拟南芥对Pto DC3000的抗病性[11]。
木薯(Manihot esculentaCrantz)是大戟科木薯属多年生灌木,是世界三大薯类作物之一[12]。木薯块根肉质,富含淀粉,以之为基础的食品和生物能源等产业是热区农业经济的重要组成部分[13]。木薯细菌性枯萎病(cassava bacterial blight, CBB)是由菜豆黄单胞菌属木薯细菌性枯萎病致病种(Xanthomonas phoseolipv.manihotis,Xpm)引起的病害,是影响我国木薯生产的重要病害之一。前人研究报道Xpm病原菌侵染木薯后,MeWRKY79和MeHsf20基因被诱导表达,MeWRKY79和MeHsf20转录因子可分别结合褪黑素合成基因MeASMT2启动子上的W-box和HSE元件来激活MeASMT2表达,调控褪黑素的合成积累,增强木薯对Xpm病原菌的抗性[14]。木薯 MeHsf3可提高水杨酸信号途径基因MeEDS1和MePR4的转录水平,参与木薯抵御Xpm病原菌的侵染[15]。
本研究基于前期转录组数据发现,我国木薯主栽品种‘SC8’在受到XpmHN11病原菌侵染时,热击蛋白转录因子MeHsfB3a的表达量显著上调。为了进一步验证MeHsfB3a在抵御木薯细菌性枯萎病中发挥的作用,本研究从‘SC8’木薯cDNA中克隆了MeHsfB3a全长序列,进行了生物信息学分析,利用qRT-PCR技术分析了该基因在木薯不同器官中的表达模式,并验证了MeHsfB3a的表达受XpmHN11病原菌诱导;采用病毒诱导基因沉默(virus-induced gene silencing, VIGS)技术沉默MeHsfB3a,验证了MeHsfB3a参与木薯对木薯细菌性枯萎病抗性抗病功能,为木薯抗病育种提供新的抗性基因。
1 材料与方法
1.1 材料
植物材料:本研究采用的木薯品种为‘华南8号’(‘SC8’),采自于海南大学木薯种质资源圃。烟草本生烟由本实验室提供。
质粒和菌株:VIGS沉默载体 pCsCMV-NC质粒由中国热带农业科学院热带生物技术研究所周鹏老师课题组赠与。大肠杆菌菌株DH5α、根癌农杆菌菌株GV3101、木薯细菌性枯萎病菌(Xanthomonas phoseolipv.manihotisHN11,XpmHN11)由本实验室分离获得。
试剂:LA Taq®高保真酶购于TaKaRa公司,限制性内切酶购于NEB公司,TRIzol Reagent购自 Invitrogen公司,RevertAidTM First Strand cDNA Synthesis Kit购自 TaKaRa公司,Nimble cloning试剂盒购自海南壹田生物科技公司,质粒小量快速提取试剂盒、PCR产物纯化回收试剂盒等均购自北京艾德莱生物科技公司。
引物:本实验的引物均由生工生物科技(上海)有限责任公司合成,引物序列如见表1。
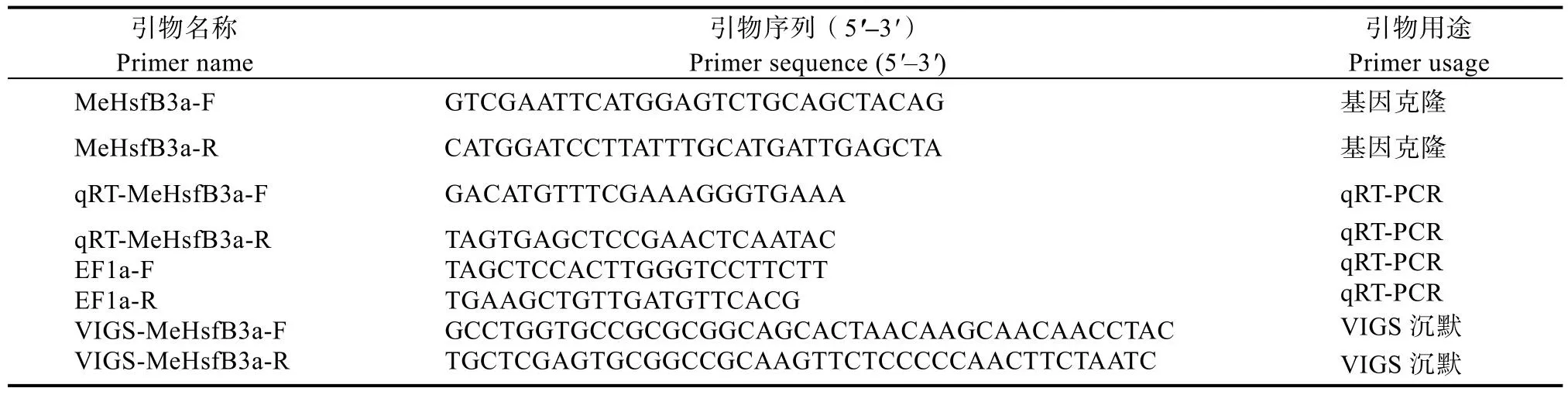
表1 引物序列Tab.1 Primer sequence
1.2 方法
1.2.1MeHsfB3a基因的克隆 采用TRIzol法提取‘SC8’木薯总RNA,用微量分光光度计检测提取的总RNA浓度和纯度,通过1%琼脂糖凝胶电泳检测提取的总RNA质量。采用RevertAidTM First Strand cDNA Synthesis Kit试剂盒反转录为cDNA,用于基因扩增。根据木薯基因组数据库phytozome中获得的MeHsfB3a(Manes.09G02-0300.1)序列信息,采用Premier 5.0软件设计扩增引物MeHsfB3a-F、MeHsfB3a-R。以cDNA为模板进行PCR扩增,克隆木薯MeHsfB3a基因编码区序列。反应体系为:TaKaRa LATaq0.2 μL,10×LA PCR Buffer 2 μL,dNTP Mixture 4 μL,上下游引物各 0.5 μL,模板 2 μL,加 ddH2O 补至20 μL。反应程序为:94℃预变性 3 min;94℃ 30 s,56℃ 30 s,72℃ 1 min循环 35次;72℃延伸10 min。用PCR产物纯化回收试剂盒回收目的片段,连接pEASY-Blunt载体上,转化进大肠杆菌DH5α感受态,挑取阳性单克隆进行菌落PCR检测,送生工生物科技(上海)有限责任公司进行测序后用质粒小量快速提取试剂盒提取pEASY-Blunt-MeHsfB3a质粒。
1.2.2 MeHsfB3a生物信息学分析 将测序得到的片段序列与预测的MeHsfB3a基因编码区序列进行比对,确定扩增的序列的正确性。MeHsfB3a蛋白的理化性质如氨基酸数、分子式、分子量、等电点、不稳定系数、脂肪指数和平均亲水系数由在线网站ExPASy(https://web.expasy.org/protparam/)进行预测。运用 GSDS(Gene Structure Display Server)在线网站分析MeHsfB3a外显子-内含子的结构。采用 Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)在线软件对MeHsfB3a蛋白进行亚细胞定位预测。
1.2.3MeHsfB3a基因的表达分析 设计MeHsfB3a进行qRT-PCR实验的引物:qRT-MeHsf-B3a-F、qRT-MeHsfB3a-R,以木薯新叶、成熟叶、顶芽、叶柄、块根、韧皮部、须根为模板进行该基因表达的组织特异性分析,分别以接种了10 mmol/L MgCl2和XpmHN11病原菌不同时间点(0、3、6 h和1、3、6 d)的cDNA为模板进行qRT-PCR,分析MeHsfB3a是否响应病原菌侵染。qRT-PCR反应体系为:TB Green Premix ExTaqⅡ(2×) 10 μL,上下游引物各 1 μL,cDNA 2 μL,加ddH2O补至20 μL。反应程序为:95℃预变性30 s;40个循环包括95℃变性 5 s,58℃退火 30 s,72℃延伸30 s,最后72℃延伸5 min。每个样品设置3个重复,以EF1α为参照基因。采用2-ΔΔCT法计算基因表达值。
1.2.4MeHsfB3a的VIGS载体构建与侵染‘SC8’木薯 采用在线软件 SGN VIGS Tool(https://vigs.solgenomics.net/)设计诱导MeHsfB3a的沉默靶基因片段。以 1.2.1获得的pEASY-Blunt-MeHsfB3a质粒为模板,特异性引物VIGS-MeHsfB3a-F、VIGS-MeHsfB3a-R扩增靶标片段,采用 Nimble cloning试剂盒构建pCsCMV-MeHsfB3a重组质粒。将重组质粒转入农杆菌 GV3101,挑取阳性单克隆于含卡那霉素和利福平(浓度均为50 mg/L)的LB液体培养基中过夜培养,菌液浑浊后4000 r/min,5 min收集菌体,用10 mmol/L MgCl2清洗菌体2次,加重悬液:1 mol/L MES(吗啉乙磺酸)+1 mol/L MgCl2+200 mmol/L AS(乙酰丁香酮)重悬菌体并调整OD600=0.75,每个处理组设置生物学重复3株,每株选择4片发育健康的叶片,用1 mL注射器将悬浮菌液注射进木薯每片裂叶背部主叶脉两侧,每片叶注射8个孔,每个孔注射约10 μL。以含有CsCMV病毒全序列和指示基因ChlI沉默靶点的重组质粒pCsCMV-B+为正对照。ChlI是组成镁螯合酶的亚基之一,是催化叶绿素生物合成的第一步,沉默ChlI可导致植株出现白化退绿表型。仅含有CsCMV病毒全序列的质粒pCsCMV-A-为负对照。侵染后45 d,正对照叶片出现白化退绿表型,分别提取负对照组和实验组木薯新叶总RNA并反转录为cDNA检测目的基因表达量。
1.2.5XpmHN11病原菌侵染VIGS沉默植株的抗病功能鉴定 从-80℃冰箱中取出保存的XpmHN11,划线培养于 LPGA固体培养基上,28℃倒置培养 2 d;蘸取长出的单克隆菌落于500 μL的LPGA液体培养基中,28℃,200 r/min摇床培养20 h左右;吸取浑浊的菌液200 μL,并用涂布棒均匀涂到LPGA固体培养基上,28℃培养24 h左右,10 μmol/L MgCl2洗下菌体,放置在50 mL离心管中;4000 r/min离心5 min收集菌体,用 10 μmol/L MgCl2洗菌 2 次,再用 10 μmol/L MgCl2重悬,调OD600=0.1。使用1 mL注射器将XpmHN11菌液注射到MeHsfB3a沉默植株新叶的叶脉两侧,选择4片叶片,每片裂叶注射4个孔,每个孔注射约 10 μL,注射后做上标记。木薯叶片侵染XpmHN11后,分别取0、3、6 d的叶片样品,观察水渍状病斑的扩散情况并拍照记录,使用ImageJ软件计算病斑面积。
2 结果与分析
2.1 MeHsfB3a基因的克隆
以‘SC8’木薯 cDNA为模板,PCR克隆MeHsfB3a基因全长,获得了大小约为750 bp的目的片段(图1)。将该片段纯化、回收并连接中间载体 pEASY-Blunt,转化大肠杆菌获得阳性克隆进行测序分析。结果表明扩增得到的MeHsfB3a片段大小为729 bp,编码242个氨基酸。

图1 MeHsfB3a基因克隆Fig.1 Cloning of MeHsfB3a
2.2 MeHsfB3a的生物信息学分析
利用在线网站 ExPASy分析木薯 MeHsfB3a蛋白的生理生化性质。结果显示 MeHsfB3a蛋白理论分子式:C1224H1934N350O383S9,理论相对分子质量(Mw)为27.9 kDa,理论等电点pI为7.59。该蛋白富含:谷氨酸 Glu(10.0%)、亮氨酸 Leu(9.1%)、赖氨酸 Lys(8.7%)、丝氨酸 Ser(8.3%)、精氨酸(7.5%)、苏氨酸(6.6%)、天冬酰胺 Asn(5.8%)。亲水指数从-2.744到2.222(负值表示亲水性,正值表示疏水性),亲水性平均指数-0.880,表明该蛋白水溶性较好(图2)。理论不稳定系数为56.86,属于不稳定蛋白质。脂溶指数为65.98。利用GSDS在线网站分析MeHsfB3a基因的结构特点,结果显示,该基因含有2个外显子和 1个内含子。利用在线网站 Plant-mPLoc SEfvEf预测MeHsfB3a蛋白可能定位于细胞核。

图2 木薯MeHsfB3a蛋白亲水性分析Fig.2 Hydrophilicity analysis of cassava MeHsfB3a
2.3 MeHsfB3a基因的表达模式分析
采用qRT-PCR技术对MeHsfB3a基因在木薯新叶、成熟叶、顶芽、叶柄、块根、韧皮部、须根的表达量进行分析。结果发现MeHsfB3a基因在成熟叶的表达量最高,约为新叶的45倍,其次为须根、叶柄,在其他部位中的表达极低(图3A)。木薯‘SC8’叶片接种了XpmHN11病菌0、3、6 h和1、3、6 d后,以接种了10 mmol/L MgCl2的木薯叶片为对照,提取叶片RNA并反转录为cDNA进行MeHsfB3a基因表达量分析,结果发现接种1 d后该基因的表达量急剧升高,其中在6 d时,表达量提高了60倍。这说明MeHsfB3a基因的表达受XpmHN11病菌的诱导(图3B)。

图3 MeHsfB3a基因表达模式Fig.3 Expression pattern of MeHsfB3a
2.4 VIGS沉默验证MeHsfB3a参与木薯对细菌性枯萎病抗病过程
2.4.1 VIGS沉默效果分析 接种VIGS实验的正对照pCsCMV-B+载体,在侵染45 d后,木薯新叶出现了白化退绿现象,而负对照 pCsCMV-A-与pCsCMV-MeHsfB3a重组质粒的叶片无变化,这表明pCsCMV-NC系统已经对木薯内源的ChlI基因产生了沉默(图4)。提取实验组 pCsCMVMeHsfB3a和负对照 pCsCMV-A-的木薯新叶的总RNA并反转录为cDNA,通过qRT-PCR检测目的基因沉默效率。每组数据来自3个生物学重复(3株),结果如图(图5),与负对照植株相比,沉默植株新叶中的MeHsfB3a表达量下降了68.26%~82.44%,说明MeHsfB3a基因在新叶受到了有效沉默,可以对新叶进行后续抗性实验。

图4 VIGS试验后木薯叶片表型变化Fig.4 Phenotypic changes of cassava leaves after VIGS test

图5 VIGS沉默MeHsfB3a的效果检测Fig.5 Detection of target gene expression after VIGS
2.4.2 VIGS沉默植株抗病性检测 为了验证MeHsfB3a在木薯抗细菌性枯萎病过程中的功能,本研究将病原菌XpmHN11接种至MeHsfB3a沉默植株(pCsCMV-MeHsfB3a)和负对照植株(pCsCMV-A-)新叶叶片。在接种 3 d时,MeHsfB3a沉默植株和负对照植株叶片均出现水渍状病斑,但是MeHsfB3a沉默植株的病斑扩散面积较大。接种6 d时,MeHsfB3a沉默植株的病斑进一步扩大,而负对照植株病斑缓慢扩大(图6A)。
为了使病斑面积具体量化,采用ImageJ软件计算接种病原菌3 d和6 d后的病斑面积,并制作柱状图进行显著性分析。在接种3 d时,MeHsfB3a沉默植株的病斑面积是负对照植株的 2.22~2.38倍;在接种6 d时,MeHsfB3a沉默植株的病斑面积是负对照植株的1.86~2.28倍(图6B)。此结果表明,沉默了MeHsfB3a基因后,木薯对XpmHN11病原菌的敏感性增加。

图6 XpmHN11侵染后不同时间点植株的抗性分析Fig.6 Phenotypes ofXpmHN11 infected leaves at different time points
3 讨论
植物 Hsfs不仅通过特异地结合热激蛋白(heat shock protein, Hsp)基因启动子上的热击响应元件(heat shock element, HSE),调控Hsp基因的表达,增强植物的耐热性,还可以调控许多抗逆基因的表达,提高植物抵御生物及非生物胁迫的能力[16]。Hsfs由多个基因家族编码,其蛋白质结构高度保守,根据不同的保守结构域可分为HsfA、HsfB、HsfC三类[17]。HsfA亚家族在热、干旱、盐和氧化损伤反应中的功能已经有很多研究报道,然而对HsfB、HsfC家族的了解还较少。最近的研究表明,HsfB和HsfC家族在植物对生物和非生物胁迫的反应中也具有重要的作用。过量表达OsHsfB2b的转基因粳稻脯氨酸含量下降,导电率上升,植株抗旱性下降[18]。5℃低温处理水稻,水稻 HsfA族和 C族成员表达量上调,B族成员表达量下降[19]。低温条件下甘蓝BraHsfC039的表达量上升,小麦过量表达TaHsfC3后,在低温处理时可承受更长时间[20]。
本研究通过RT-PCR从感病品种‘SC8’基因组中扩增获得 HsfB家族基因:MeHsfB3a,该基因全长为729 bp,编码289个氨基酸,含有2个外显子和 1个内含子。亚细胞定位分析发现,MeHsfB3a预测定位于细胞核,符合转录因子的定位特征。通常HsfB族编码的蛋白缺少转录激活结构域,可能对靶基因的转录起抑制作用。拟南芥AtHsfB1和 AtHsfB2b定位于细胞核直接抑制AtHsfA2、AtHsfA7a、AtHsfB1和AtHsfB2b的表达[21]。Hsfs在植物的不同器官中表达模式具有差异,如玉米ZmHsf06在根中表达量较高[22],而ZmHsf12在新叶中表达量高[23]。表达分析显示MeHsfB3a在成熟中表达量最高,在其他器官中的表达量较低。有研究发现木薯MCOL1522品种在细菌性枯萎病菌株XamORST4侵染时MeHsfB3a基因表达量上调[24]。因此,推测MeHsfB3a可能在木薯抵御细菌性枯萎病过程中发挥作用,但该研究并未克隆MeHsfB3a基因序列并验证该基因的功能。本研究将在海南分离得到的菌株XpmHN11接种于‘SC8’木薯叶片发现MeHsfB3a的表达被病原菌侵染激活,最高达到 60倍,表明该基因参与‘SC8’木薯对细菌性枯萎病病原菌的响应过程。前人的研究发现,木薯的其他Hsf家族基因成员也同样受细菌性枯萎病病原菌侵染的诱导。MeHsf3在Xpm病原菌侵染木薯SC124叶片6 h后,转录水平上调了8倍[15]。
病毒诱导的基因沉默技术(VIGS)是一种方便有效的反向遗传学技术,可以通过构建重组病毒载体来沉默植物体内的目的基因。目前已经开发了多种植物病毒载体,可以对植物各个组织的基因进行沉默[25]。其中由烟草脆裂病毒(Tobacco rattle virus,TRV)介导的VIGS体系对宿主影响较小,因此应用最为广泛。WEI等[14]在 2017年利用 TRV介导的沉默体系对木薯MeHsf20和MeWRKY79进行了沉默,结果发现沉默了MeWRKY79和MeHsf20的植株中褪黑素合成相关基因MeASMT2的转录下调,导致褪黑素减少,植株抗病性降低。LI等[26]在2017年同样以TRV介导的沉默体系诱导沉默木薯 bZIP转录因子中的MebZIP3和MebZIP5,沉默效率分别达到了50%和 40%,接种木薯细菌性枯萎病病原菌后,沉默植株表现出疾病敏感表型、防御相关基因转录水平降低、胼胝质沉积减少,说明了MebZIP3和MebZIP5正调控木薯细菌性枯萎病。最近TUO等[27]成功利用木薯普通花叶病毒(CsCMV)开发了适用于木薯的新型VIGS系统pCsCMV-NC。本研究利用pCsCMV-NC介导的VIGS技术沉默木薯MeHsfB3a基因,实时荧光定量结果显示实验组MeHsfB3a基因表达量均降低了68.26%~82.44%,这说明MeHsfB3a基因已经受到了显著沉默。用XpmHN11病原菌接种MeHsfB3a基因沉默木薯植株,发现木薯对细菌性枯萎病的抗病能力显著降低。基于以上结果,本研究证明了热击蛋白转录因子基因MeHsfB3a受XpmHN11病原菌诱导表达,参与木薯抗细菌性枯萎病的过程。在未来的研究工作中,本实验室将利用转基因技术,过量表达MeHsfB3a进一步验证该基因的抗病功能,并解析 MeHsfB3a转录因子调控的下游抗病通路。
猜你喜欢
世界热带农业信息(2018年11期)2018-01-17
广东第二课堂·小学(2017年5期)2017-05-27
创新作文(小学版)(2017年29期)2017-04-04
中国果业信息(2016年2期)2016-01-29
世界热带农业信息(2014年8期)2014-09-23
音乐生活(2014年3期)2014-04-25
中国果业信息(2012年9期)2012-01-23
植物营养与肥料学报(2011年3期)2011-10-24
植物营养与肥料学报(2010年4期)2010-11-06
环球时报(2009-08-27)2009-08-27
