
湖州河川沙塘鳢群体线粒体DNA cyt b 基因序列的遗传多样性分析
2023-02-26 06:02:10卞玉玲刘士力刘一诺贾永义李飞迟美丽郑建波程顺顾志敏
水产学杂志 2023年1期
卞玉玲,刘士力,刘一诺,贾永义,李飞,迟美丽,郑建波,程顺,顾志敏
(1.上海海洋大学农业农村部淡水水产种质资源重点实验室,上海 201306;2.浙江省淡水水产研究所,农业农村部淡水渔业健康养殖重点实验室/浙江省淡水水产遗传育种重点实验室,浙江 湖州 313001;3.上海海洋大学上海水产养殖工程技术研究中心,上海 201306;4.上海海洋大学水产科学国家级实验教学示范中心,上海 201306)
河川沙塘鳢(Odontobutis potamophila)属于鲈形目(Perciformes)、虎鱼亚目(Gobioidei),分布于钱塘江、黄河和长江中、下游等流域[1]。因其具有肉鲜味美、氨基酸含量丰富的特点,深受消费者的喜爱。为了满足不断增加的消费需求,野生沙塘鳢被过度捕捞,从而导致其数量锐减,IUCN(世界自然保护联盟)红色名录将其评级为信息缺乏物种[2,3]。近年来,随着苗种繁育技术的突破,为沙塘鳢规模化养殖打下了坚实的基础[4]。目前,在我国已记载的沙塘鳢属鱼类共4 种,分别是中华沙塘鳢(O.sinensis)、河川沙塘鳢(O.potamophila)、鸭绿沙塘鳢(O.yaluensis)和海丰沙塘鳢(O.haifengensis)[5]。在水产行业中,线粒体DNA(Mitochondrial DNA,mtDNA)标记技术被广泛应用于遗传多样性、亲缘关系鉴定和种属分类的研究。沙塘鳢具有较高的营养与经济价值,通过线粒体cyt b 基因序列开展遗传结构研究,对于种群的保护与良种选育具有重要意义。由于mtDNA 具有母系遗传的特性,使其能够作为分析群体遗传进化的依据,又因进化速度快、拷贝数多、检测方便等优点,为研究不同地理群体的水产动物种内及种间的生存状态、遗传变异,以及对将来的物种保护和环境调节奠定基础。谢楠等[6]将扩增得到的河川沙塘鳢cyt b 基因序列与同科22 种鱼的cyt b 基因进行比对,为沙塘鳢属和塘鳢科鱼类的系统分类提供依据。王丹等[7]以cyt b 基因对分布于珠江及长江的9 个野生中华沙塘鳢种群进行遗传多样性分析,基因流方向大致为长江流域种群流向珠江。张烨等[8]采用线粒体基因对安徽的4 个河川沙塘鳢群体进行评价,发现沙塘鳢群体间的分化程度较高。这些研究为沙塘鳢的种质资源开发与利用提供科学的考量。线粒体DNA 结构简单,其序列变异速率远高于核基因组,其中细胞色素b(cyt b)基因是mtDNA 自身编码的为数不多的功能蛋白之一。Zardoya 等[9]研究发现,cyt b 的进化速率适中,替换、缺失和插入等突变能够稳定持续遗传,可适用于种间、种内的遗传分析。本实验将湖州2 个群体的线粒体细胞色素b(cyt b)基因全长进行扩增,并与GenBank 中的太湖、鄱阳湖和巢湖的数据进行比较分析,探究人工繁殖对河川沙塘鳢种质资源的影响,为沙塘鳢的良种选育、种质资源保护与开发提供理论依据。
1 材料与方法
1.1 研究材料
实验采用湖州野生群体和养殖群体各1 个,其中养殖群体为同塘养殖、同批繁殖的河川沙塘鳢群体,于2017 年6 月采自浙江省淡水水产研究所八里店综合试验基地;野生群体为同批捕捞,同期采自湖州钟管镇。每个群体取32 尾样本,剪适量尾鳍组织,使用无水乙醇保存,储存于4℃冰箱中。
1.2 方法
每个样本取20~30 mg 尾鳍,采用酚-氯仿法[4]提取DNA,使用NanoDrop 2 000 测定DNA 原液浓度,并用琼脂糖凝胶电泳检测完整性,将其稀释至25 ng/μL 左右,于-20℃保存备用。
1.3 cyt b 扩增程序及测序
cyt b 扩增和测序的引物为通用引物L14724 和H15915。L14724:5'-GACTTGAAAAACCACCGTTG-3';H15915:5'-CTCCGATCTCCGGATTACAAGAC-3',引物由生工生物工程(上海)股份有限公司合成。PCR 为25 μL 反应体系:上、下游引物(10 μmol/L)各1 μL,2×PCR Mix 12.5 μL,DNA 模板25 ng,双蒸水补足体系。反应程序:94℃预变性5 min;94℃变性30 s,58.5℃退火30 s,72℃延伸2 min,共35 个循环;72℃延伸10 min。用1%的琼脂糖凝胶电泳检测产物品质,扩增产物由生工生物工程(上海)股份有限公司用正反引物进行双向测序。测序后部分有杂峰的样本,克隆后再次进行测序。
1.4 数据分析
用于比较分析序列的样本来自太湖(GenBank登录号:KM924270-KM924288)、鄱阳湖(登录号:KM924241-KM924269)和巢湖(登录号:KM924174-KM924199)。通过BioEdit 7.0 软件对序列进行分析,应用Mega 6.0 软件计算碱基含量,并采用邻接法基于Kimura's 2-Parameter 模型建立系统进化树,自举检验值(Bootstrap)设置为1 000 次。运用DnaSP 5.0软件计算单倍型数、单倍型多样性指数(H)、变异位点、核苷酸多样性指数(Pi)等,并检验中性分子进化演变,计算Tajima's D 和Fu's Fs 参数,以此推测群体的历史演化。使用Arlequin 3.1 中的分子方差分析(AMOVA)法计算遗传分化指数(Fst)。
2 结果与分析
2.1 河川沙塘鳢5 个群体线粒体DNA cyt b 区序列碱基组成
将湖州两个群体的原始序列进行软件比对和人工校对,发现八里店群体有7 个样本线粒体DNA cyt b 存在不同拷贝现象,其中2 个样本有1 个碱基缺失,5 个样本存在杂合位点。5 个群体包含138 个样本,对线粒体cyt b 1 141 bp 的DNA 序列基因进行了分析。结果显示存在变异位点378 个,占分析位点的44.26%,含有241 个简约信息位点,平均转颠换比值(TS/TV)为0.76。4 种碱基在所得序列中平均含量为A(26.8%)、G(13.8%)、T(28.0%)和C(31.4%),其中G 的含量最低,A+T 含量为54.8%,明显高于C+G 含量。
2.2 河川沙塘鳢5 个群体单倍型的发生关系
除去插入缺失的序列,在138 个个体中共定义了75 种单倍型,将分析的湖州群体单倍型序列提交GenBank,登录序列号为:MF286741-MF286759。河川沙塘鳢群体的单倍型网络中介图如图1 所示。湖州群体单倍型编号为(H1-H13),其中八里店群体具有10 种单倍型,钟管群体具有9 种单倍型,这两个群体共享6 种单倍型,与其余3 个群体不共享任何单倍型;太湖群体单倍型数量为11 种(H14-H24);鄱阳湖群体单倍型数量为25 种(H25-H49);巢湖群体拥有的单倍型最多,为26 种(H50-H75)。河川沙塘鳢单倍型具有明显的分化,除八里店和钟管群体具有共享单倍型外,其余群体都没有共享单倍型,这可能是由于八里店为养殖群体,亲本来源于钟管群体的原因,因此,将其统归为湖州群体,湖州群体位于网络中介图的中心区域,明显分成2 个部分。鄱阳湖群体的单倍型具有3 个明显分开的区域,太湖群体单倍型位于其中。巢湖群体位于另一端,呈树枝状分布。根据河川沙塘鳢单倍型的网络中介图,推断cyt b 基因序列可以区分不同来源的群体。

图1 河川沙塘鳢Cyt b 基因单倍型的网络关系图Fig.1 Median-joining network for sequence of Cyt b gene haplotypes of O.potamophila
2.3 河川沙塘鳢5 个群体线粒体DNA cyt b 基因遗传结构分析

表2 河川沙塘鳢群体线粒体cyt b 基因序列分子方差分析Tab.2 Analysis of molecular variance(AMOVA)of part sequence of cyt b in O.potamophila
使用DnaSP 5.0 软件计算河川沙塘鳢5 个群体的遗传多样性参数(表1),结果显示:5 个群体的单倍型多样性(0.869~1.000)较高,核苷酸多样性(0.01431~0.03848)具有明显分化。鄱阳湖和巢湖群体的单倍型多样性接近或等于1,核苷酸多样性约为太湖、八里店和钟管群体的两倍。其中巢湖群体的Pi值最高,为0.03848,鄱阳湖为0.03087。太湖和湖州的两个群体明显要低一些,为0.01431~0.01536。表明河川沙塘鳢5 个群体具有不同的遗传多样性。
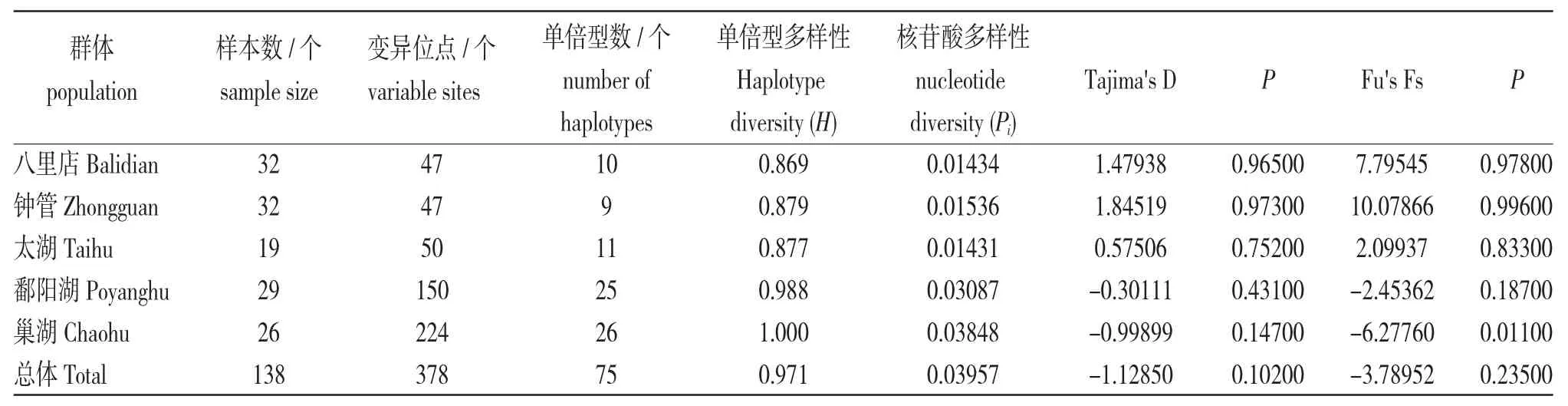
表1 河川沙塘鳢群体cyt b 基因序列的遗传多样性参数Tab.1 Genetic diversity parameters of cyt b gene fragments in O.potamophila
5 个河川沙塘鳢群体线粒体DNA cyt b 基因序列的遗传差异AMOVA 分析结果表明,群体间的遗传分化指数Fst=0.48445(P<0.01),整个遗传变异中群体间占48.44%,其余的遗传变异来自于群体内(51.56%),总体上,群体间具有较高程度的遗传分化。
2.4 河川沙塘鳢5 个群体遗传距离分析
通过Mega 6.0 软件计算各地理种群间遗传距离,所得结果见表3。由表3 可知,各群体间遗传距离为0.01493~0.05868,遗传距离差距较大。其中八里店与钟管群体的遗传距离最近,鄱阳湖与巢湖群体的遗传距离最远,构建的聚类图上也表现相同趋势(图2)。运用Arlequin3.1 计算各群体间的Fst在-0.01821~0.68483,其中八里店和钟管群体为负值,说明群体间基因交流频繁,太湖与鄱阳湖群体间的Fst小于0.25,其余群体间的Fst都大于0.25。除去钟管群体,八里店与其他群体间的Fst都在0.5 以上。八里店、钟管、太湖、鄱阳湖和巢湖群体内遗传距离分别为0.01468、0.01573、0.01449、0.03163 和0.03965。

表3 河川沙塘鳢5 个群体间的遗传距离(对角线下方)和遗传分化指数(对角线上方)Tab.3 Estimates of genetic distance(below diagonal)and genetic differentiation index Fst(above diagonal)among 5 populations of O.potamophila based on cyt b gene
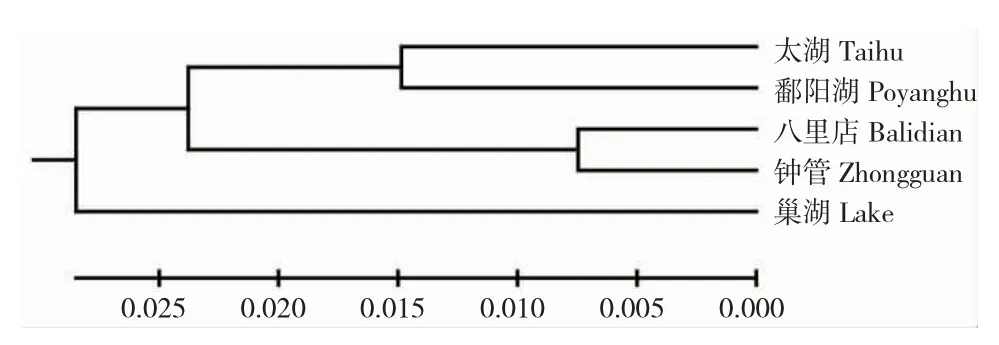
图2 基于线粒体cyt b 基因遗传距离构建的邻接树Fig.2 NJ trees of haplotypes of O.potamophila based on part sequences of the Cyt b gene
2.5 河川沙塘鳢5 个群体动态分析
对河川沙塘鳢群体进行核苷酸不配对分布分析和Tajima's D 中性检验,结果显示,河川沙塘鳢野生群体的单倍型检测中,核苷酸不配对分布(mismatch distribution)呈现多峰型(图3),虽然鄱阳湖群体和巢湖群体在中性检验中也呈现出Tajima's D 值为负值,但统计结果不显著(P>0.05)。八里店、钟管和太湖的Tajima's D 及Fu's Fs 值均为正值,且较高,表明这些群体可能没有经历过种群扩张。5 个群体Tajima's D 和Fu's Fs 检验均不显著(P>0.01)。

图3 河川沙塘鳢的核苷酸错配分布图Fig.3 The nucleotide mismatch distribution of O.potamophila
3 讨论
作为生物多样性的重要层次,遗传多样性是评价生物资源的重要依据,种群遗传多样性的度量和保护已成为对其生物资源进行评价的核心问题[10,11]。不了解重要种群的遗传多样性,就无法实现有针对性的科学保护和合理开发。种群内维持较高单倍型多样性的主要原因包括种群数量的大小、环境的不均一性及具有适应种群快速增长的特性。与淮河野生鲇(Silurus asotus)[12]、黄颡鱼(Pelteobagrus fulvidraco)[13]和乌鳢(Channa argu)[14,15]群体相比,河川沙塘鳢往往具有较高的单倍型多样性(0.869~1.000),但其核苷酸多样性(0.014 31~0.038 48)具有明显分化,可能是由于沙塘鳢生活习性等原因,短期内序列变异达不到有效积累[16,7]。八里店、钟管沙塘鳢群体表现出较高的单倍型多样性、较低的核苷酸多样性特征,符合鱼类第2 种单倍型与核苷酸多样性组合,高H 和低Pi类型[18]。鄱阳湖是我国第一大淡水湖,拥有较为发达的水系,因此,鄱阳湖沙塘鳢群体表现出丰富的遗传多样性,单倍型数量与核苷酸多样性均较高,整体水平表现为高单倍型多样性、高核苷酸多样性特征。太湖河川沙塘鳢群体的H 较低、Pi为最低,这可能与采用的样本量较少有关。
衡量种群多态性的重要指标包括种群间的遗传距离以及Fst,多态性程度与这两者存在相关性[19]。本文中,遗传距离为0.01493~0.05868,差距较大,依据种群间的遗传距离标准,由于八里店为养殖群体,亲本来源于钟管群体,二者亲缘关系较近,与其他群体亲缘关系较远。种群间的遗传距离分析与Fst的分析[20]结果相一致。基因流是指基因从一个种群转移到另一个种群,其大小影响群体间的相似性程度,基因流越大,相似性越高[21]。沙塘鳢的游泳能力与迁移趋向较弱,地理隔离较明显、遗传变异较高,可能会降低群体间的基因交流,生活史与生态习性影响种群之间的遗传变异状况[22]。刘士力等采用线粒体16S rRNA 基因片段进行扩增,对建德、钟管和八里店沙塘鳢群体进行遗传分析,AMOVA 显示群体间存在较高程度的遗传分化,群体间的Fst为0.39347,超过0.25。侯新远等[23]通过D-loop 基因分析25 种虾虎鱼类的遗传距离和系统发育树,获得不同虾虎鱼类的亲缘关系。郁建锋等利用12S rRNA和16S rRNA 基因,分析福建与太湖的河川沙塘鳢,发现大量的变异位点和简约位点,在分子水平上得出太湖沙塘鳢与其他地区沙塘鳢之间存在遗传分化差异[24],该线粒体基因的变异分化,推测与其生长习性相关。由于不同流域水产动物会形成地理隔离的状态,因而不同水系间会产生显著的种群分化[25]。
随着数据库中cyt b 基因序列数量的逐渐增多,cyt b 基因逐渐成为重要的分子标记之一[26]。目前,cyt b 基因被广泛应用于物种鉴定[27-29]、遗传结构分析[27-32]和系统发育研究[28,29]。对中国华南沿海的黄鳍棘鲷(Acanthopagrus latus)采用cyt b 基因进行分析时发现,海南至福建的8 个群体,共分为东、西两组,320 尾个体共有62 种单倍型,其中7 种单倍型为所有群体共享[30]。采用线粒体cyt b 基因序列对西江流域广西境内6 个江段139 尾卷口鱼(Ptychidio jordani)进行遗传多样性分析时共定义20 个单倍型,其中Hap3 为优势单倍型,存在于所有群体中,且占总个体数的39.57%[31]。在本实验中,除了湖州的八里店与钟管群体共享6 种单倍型外,其他群体间基本不共享单倍型。这可能是八里店养殖群体亲本来源于钟管野生群体的原因。不同种群间基因流动较少,这可能与河川沙塘鳢游泳力较弱相关。应用cyt b 基因可以明确地将湖州、太湖、鄱阳湖和巢湖群体之间区分开来。这与中国少鳞鳜(Coreoperca whiteheadi)类似,贵州省3 个中国少鳞鳜群体间不共享任何单倍型。中国少鳞鳜为溪涧性鱼类,生存环境限制了其迁移能力,地理种群间的交流机会少,变异的积累导致了遗传分化加大[32]。
综上所述,本研究通过对河川沙塘鳢八里店和钟管群体的cyt b 基因进行检测,结合其他3 个河川沙塘鳢群体(太湖、鄱阳湖和巢湖)共同分析其遗传多样性及其可能的适应机制,为湖州河川沙塘鳢的种质资源分类评估、种群关系研究及长期稳定保护提供了重要的参考依据。
猜你喜欢
垂钓(2024年1期)2024-03-08 04:09:35
流程工业(2022年12期)2022-12-30 06:36:48
水利水电快报(2022年7期)2022-07-18 01:02:40
海洋通报(2021年1期)2021-07-23 01:55:14
生物学通报(2021年4期)2021-03-16 05:41:26
现代出版(2019年6期)2020-01-14 02:20:36
文苑(2018年22期)2018-11-15 03:24:23
老友(2017年4期)2017-02-09 00:26:04
黄河之声(2016年8期)2016-10-21 02:26:58
癌变·畸变·突变(2014年1期)2014-03-01 04:39:36
