
“鱼-植物”共生模式下不同植物根际细菌多样性及群落结构研究
2023-02-26 06:02:22胡建勇李晓东翟旭亮时春明焦飞高攀
水产学杂志 2023年1期
胡建勇,李晓东,翟旭亮,时春明,焦飞,高攀
(1.新疆维吾尔自治区水产科学研究所农业农村部西北地区渔业资源环境科学观测实验站,新疆 乌鲁木齐 830000;2.重庆市水产技术推广总站,重庆 400020)
“鱼-植物”共生模式是通过植物吸收利用养殖水体中氮、磷的一种综合种养模式,因此,提高氮磷的利用效率对该产业的发展具有现实意义[1]。研究表明,植物对氮磷吸收与利用效率受种类等多方面因素的影响。水稻(Oryza sativa)对氮素的利用效率随施氮量的增加而下降[2];与凤眼莲(Eichhornia crassipes)和水芹菜(Oenanthe javanica)相比,水蕹菜(Ipomoea aquatica)对富营养化水质净化效果表现更加优秀[3];进一步研究发现,植物根系微生物中有益菌群能够提升植物营养元素的吸收与利用效率,促进植物生长发育,周冬梅等[4],柏文恋等[5]认为,根系微生物能够促进土壤中难溶态无机磷溶解和有机磷的矿化,促进磷的利用。而在“鱼-植物”共生模式应用的实践中,提高水体中氮磷利用效率的主要方法为多种植物组合模式,彭刚[6]利用“挺水植物荷花(Nelumbo)、芦苇(Phragmites australis)等+沉水植物植物苦草(Vallisneria natans)、伊乐藻(Elodea nuttallii)等+浮水植物水葫芦(Eichhornia crassipes)、浮萍(Lemna minor)等”的联合净化模式去除了水体中50%~70%的氮磷。基于上述研究进展,为解决新疆地区水产养殖面临的尾水生态治理的技术瓶颈,作者开展了以鱼菜共生为代表的“鱼-植物”综合种养模式的应用,取得了良好的效果。但提高共生植物对不同类型尾水氮、磷利用效率,需要强化不同植物的协同作用,重视其根际微生物的作用。为此,本文利用高通量测序技术分析了水稻、茨菰、水蕹菜、水芹菜、苏丹草等植物根系细菌群落结构,同时采集植物种植区水样进行水质分析,研究“鱼-植物”共生模式下不同植物根系细菌多样性及群落结构与水体中氮、磷含量的关系,以期为进一步丰富“鱼-植物”共生理论,为加快该技术的推广应用提供参考。
1 材料与方法
1.1 材料
以水稻(Oryza sativa)、茨菰(Sagittaria sagittifolia)、水蕹菜(Ipomoea aquatica)、水芹菜(Oenanthe javanica)和苏丹草(Sorghum sudanense)为研究对象;其中水稻品种为秋田小町,经育苗,大田生长;茨菰购自乌鲁木齐米东区种植户;水蕹菜购自乌鲁木齐新市区种植户;苏丹草采用种子直播法;水芹菜购自湖北省黄冈市。上述植物于5 月25 日定植于花盆,每种植物定植16 盆,随后置于植物浮床,种植于新疆水生野生动物救护中心循环水养殖池塘,其养殖模式为浮式平台养殖模式:构建养殖单元和生态净化单元,养殖单元为8 个直径3 m、高1.2 m 的圆形漂浮养殖池,配置水循环和集排污、增氧系统,分别放养易捕鲤(211.35±14.19)g/ 尾、松浦镜鲤(212.36±15.65)g/ 尾,放养密度均为10 kg/m3水体;生态净化单元放养500 g 以上的鲢100 尾/667m2、鳙50 尾/667m2,并利用本试验所用植物净化水质。8 月25 日剪取一定长度的植物根系经PBS缓冲液(pH7.4)漂洗后,置于20 mL PBS 缓冲液(pH=7.4)中,摇床震荡30 min 后经4℃12 000 r/min离心5 min 富集根系土壤备用;同时采集植物种植区水样,部分经4℃12 000 r/min 离心5 min 富集备用,其他用作水质分析。
1.2 方法
1.2.1 DNA 提取
根系及水样微生物DNA 提取使用OMEGA 公司E.Z.N.ATM Mag-Bi-nd Soil DNA 试剂盒并经凝胶检测后备用。
1.2.2 PCR 扩增及高通量测序
根系及水样微生物DNA 使用Life 公司Qubit2.0 DNA 试剂盒精确定量后,用上海生工的细菌16S rDNA 通用引物进行第一轮扩增。引物序列如下:341F:CCCTACACGACGCTCTTCCGATCTG(barcode)CCTACGGGNGGCWGCAG;805R:GACTGGAG TTCCTTGGCACCCGAGAATTCCA(barcode)GACTA CHVGGGTATCTAATCC。上述PCR 产物引入Illumina 桥式PCR 兼容引物进行第二轮扩增后,用0.6倍的磁珠(Agencourt AMPure XP)富集后经Life 公司Qubit2.0 DNA 试剂盒精确定量后委托上海生工使用Illumina MiseqTM平台测序。
1.2.3 水质测定
水样的采集、贮存、运输和处理具体参照GB/T12999-91,氨氮(NH4+-N)含量采用纳氏试剂比色法(GB7479-87)测定;亚硝酸盐氮(NO2--N)含量采用萘乙二胺分光光度法(GB7493-87)测定;硝酸盐氮(NO3--N)含量采用酚二磺酸分光光度法(GB7480-87)测定;总氮(TN)含量采用碱性过硫酸钾消解紫外分光光度法(GB11894-89)测定;总磷(TP)含量采用钼酸铵分光光度法(GB11893-89)测定。
1.2.4 数据分析
测序结果经去除接头、引物序列后拼接、过滤、去除嵌合体,最终得到可用于划分操作分类单元(Opera-tional taxonomic units,OTU)的序列数。水质试验结果用平均数±标准差表示,采用SPSS 17.0统计软件进行关联性分析。
2 结果与分析
2.1 不同植物根系细菌群落多样性
水稻(OS-1)、茨菰(ST-1)、水蕹菜(IA-1)、水芹菜(OJ-1)、苏丹草(SS-1)和水样(W-1)共得到有效OTU 数见表1。
由表1 可知,在“鱼-植物”共生模式下,不同植物根系细菌群落测序所得序列数差异极大,最高的茨菰是最低的水芹菜的1.7 倍。为了更好地反应研究的真实情况,本试验构建了各样品OTU 的韦恩图(图1)和树状结构(图2)。
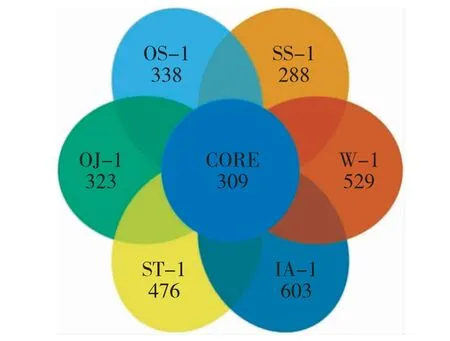
图1 不同样品OTU 数花瓣图Fig.1 Petal graph based on OTUs of different sample

表1 测序数据处理结果Tab.1 The results of sequencing data

图2 不同样品OTU 数聚类树图Fig.2 Bray-crutis tree of different samples OTUs
由图1 可知,不同植物在“鱼-植物”共生模式下根系细菌OTU 绝大部分为特有类型,彼此间差异极大,除水样外,水蕹菜最高,苏丹草最低。由图2可知,除水样外,茨菰和水稻聚于一个分支,说明它们的OTU 相似性较高,这可能与它们同属单子叶植物有关;而苏丹草与其他四种植物差异最大,这可能与其原本为陆生植物有密切关系。基于高通量的测序技术是研究根系微生物群落结构的主要手段,但对于该技术而言,保证测序结果的准确性即需要准确反映被测样本微生物群落的丰度和多样性。为此,本课题组构建了稀释曲线(图3)和Rank-abundance 曲线(图4)。
由图3 可知,随着扩增数的增加稀释曲线逐渐平缓,表明随着扩增数的增加产生新的OTU 的概率低,说明本研究的测序数据合理。而图4 中各条曲线凸凹不平,说明在“鱼-植物”共生模式下,各植物根系细菌群落组成均匀度低,不同种类菌群差异较大。本研究通过基于RDP classifier 分类方法,在科(family,图5)、属(genus,图6)水平上对以上5 种植物根系细菌菌群进行了分类分析,同样证实了不同植物在同一种植模式下根系细菌群落结构据巨大差异。
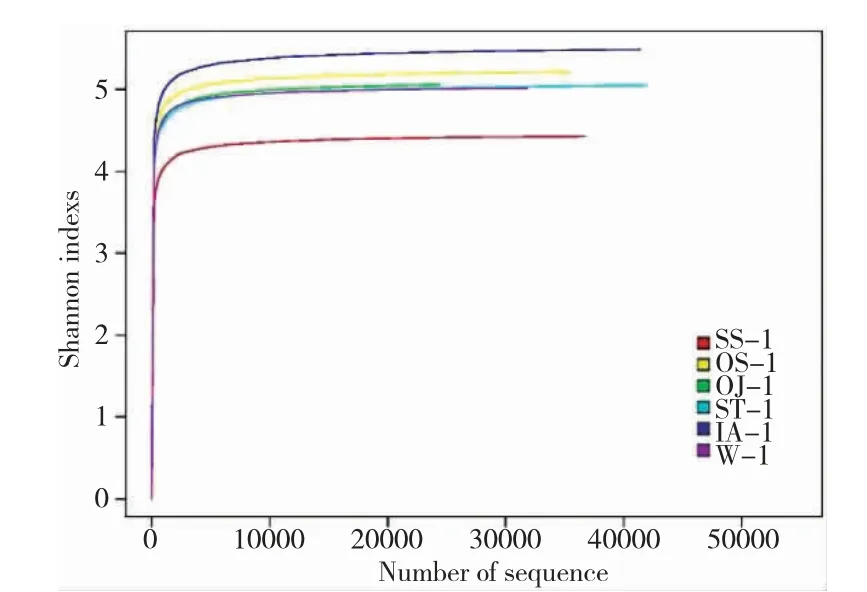
图3 稀释曲线Fig.3 Rarefaction curve of samples
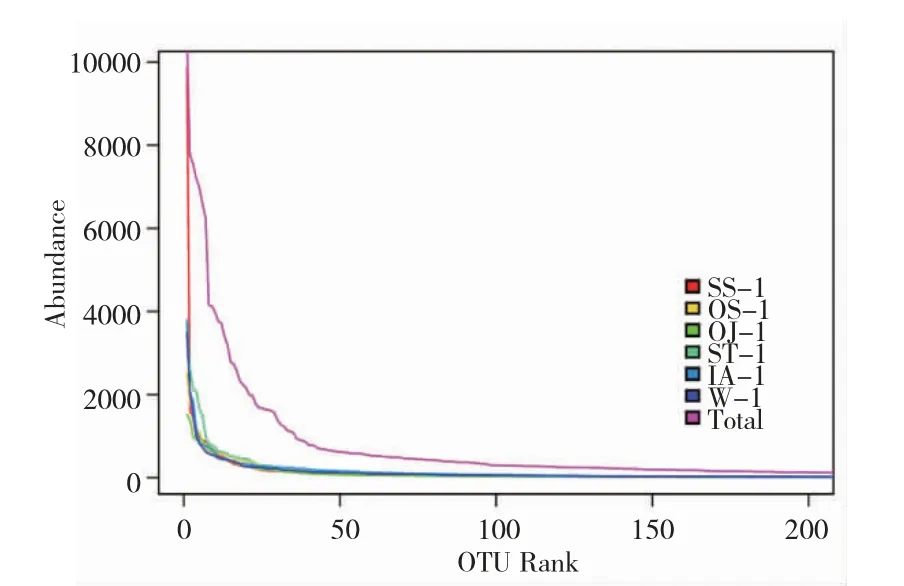
图4 Rank-abundance 曲线Fig.4 Rank-abundance curve of samples
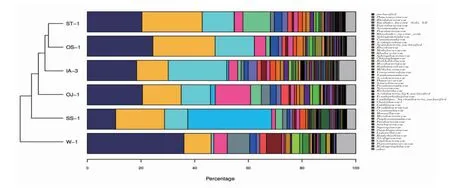
图5 科水平上不同处理样品物种丰度Fig.5 Abundance of species with different treatments on family level
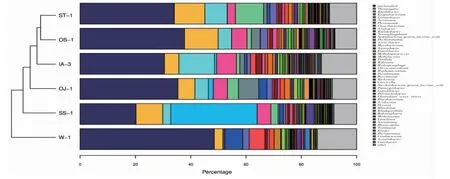
图6 属水平上不同处理样品物种丰度Fig.6 Abundance of species with different treatments on genus level
2.2 根系细菌群落结构与池塘水质关联性分析
在池塘“鱼-植物”共生模式下,水体是植物所需氮、磷及其他营养元素的唯一来源;水体中的氮、磷浓度能够影响植物的生长发育;而根系中的有益微生物菌群又有利于植物对各类营养元素的吸收。鉴于此,本研究对水质分析结果(表2)同上述五种植物根系细菌种群含量均高于0.1%的优势菌群在科属水平进行相关性分析。结果发现,以下各科(包括Planctomycetaceae、Enterobacteriaceae、Aeromonadaceae、Flavobacteriaceae、Sphingomonadaceae、Comamonadaceae、Acidimicrobiaceae、Rhodocyclaceae、Chitinophagaceae、Burkholderiaceae、Ectothiorhodospir aceae、Caldilineaceae、Desulfobacteraceae 和 Parachlamydiaceae)、各属(包括Rhodobacter、Ilumatobacter、Novosphingobium、Dechloromonas、Aquisphaera、Enterobacter、Pirellula、Ralstonia、Hydrogenophaga、Litorilinea、Arenimonas、Phreatobacter、Bosea 以及Planctomicrobium)与水体中的N、P 含量呈显著(P<0.05)或极显著相关性(P<0.01)(表3)。

表2 “鱼-植物”共生模式的水质测定结果Tab.2 The results of water quality in the“fish-plant”symbiosis system

表3 水质与植物根系细优势菌群在科、属水平上的相关性分析Tab.3 Correlation analysis of water quality and plant shizosphere microflora at family and genus levels
3 讨论
3.1 根系细菌与植物种类、生长环境的关系
植物根系对营养元素的吸收、利用与生长环境密切相关;同时,根系菌群在参与相关生理活动中起着重要作用。因此,研究植物生长环境与根系微生物群落结构关系将对提高植物对营养元素利用效率、增强抗逆性具有重要意义。例如,在盐碱环境中碱蓬(Suaeda glauca)根系中Echinicola 属、芦苇中Bacillus 属和Filobacillus 属菌群具有较高丰度[7];而在人工湿地中,相比梭鱼草(Pontederia cordata)和再力花(Thalia dealbata Fraser),美人蕉(Canna indica L.)根系具有较高的菌群多样性[8]。这说明同一生长环境下,植物根系微生物菌群结构与植物的种类密切相关,在荒漠环境下,半日花(Helianthemum songaricum)中优势属为Sphingomonas、Gemmatimonas、Aciditerrimonas、Pseudomonas、Adhaeribacte而四合木(Tetraena mongolica)中的Sphingomonas、Gemmatimonas、Aciditerrimonas、Steroidobacter、Streptophyta 为优势属[9];现有证据表明,植物根系分泌物类别和数量能够影响植物根系微生物群落结构[1,10]。本研究中5 种植物在“鱼-植物”共生模式下根系细菌OTU 结果看,绝大部分为特有类型,彼此间差异极大,进一步证实了上述结论。
3.2 根系细菌促进氮磷利用
植物对不同化合态的N、P 的吸收利用效率差异极大,对氮的利用主要为铵态氮和硝态氮,对磷的利用主要是处于溶解态的有机或无机磷[11,12]。在“鱼-植物”共生模式下,水体中的P 浓度含量较低,植物根系微生物通过生理生化途径提供可溶态磷以满足植物生长发育所需。现有证据表明,巨大芽孢杆菌(Bacillus megaterium)、假单胞菌属(Pseudomonas)、欧文氏菌属(Ewingella)等根系细菌能够参与相关生理活动,而且在根系中的真菌,尤其是丛枝菌根真菌在提升磷的利用效率中作用更为显著[13,14];不同植物对P 的利用效率有较大差异,相比灯心草(Juncus effusus L.)和蝴蝶花(Iris japonica Thunb.),石菖蒲(Acorus tatarinowii Schott)吸收同化磷素的效率更高[15]。这种差异可能与根系中与磷利用相关菌群数量有关,并已在水芹菜的研究中获得了证实[16]。与P 利用相比,植物对N 的利用表现出更为复杂的生理生化特性;就参与N 利用的根系微生物而言,涉及氨氧化细菌、亚硝酸盐氧化细菌和其他消化细菌、反硝化细菌、根瘤菌乃至部分真菌,但细菌在植物对N 的吸收利用中起着主要作用[1,16]。同样,不同植物中与N 利用相关的菌群存在着巨大差异,例如,芦苇根际比香蒲更适合亚硝酸细菌的生长[17]。在逆境条件下,植物根系细菌群落有较高的固氮和解磷等促生能力[9],而在“鱼-植物”这一种养模式下,水稻根系中存在与氮代谢如固氮、硝化、反硝化及除磷、降解纤维素、降解各类有机物、吸收金属离子、产生絮凝剂等作用的细菌[18],这与本研究所得到的结果一致,并且这些菌群在促进植物营养元素吸收中具有潜在的利用价值,相关研究已在种植业中开展了广泛研究并在土塘改良[19]、肥力提升[20]中取得了良好效果。
3.3 结论
“鱼-植物”共生模式下,不同种类植物根系细菌群落结构差异极大,这种差异与植物对营养元素的吸收及利用密切相关,且不同植物对各类营养元素的吸收及利用也存在巨大差异。鉴于此,为了提高共生植物对水产养殖中水或尾水的净化效率,建议在选择该技术时应采用多种植物搭配模式。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
小学生作文(低年级适用)(2020年10期)2020-11-10 09:12:12
中国建筑装饰装修(2020年6期)2020-07-10 09:41:16
福建基础教育研究(2019年2期)2019-09-10 07:22:44
福建基础教育研究(2019年2期)2019-05-28 08:39:49
今日农业(2019年14期)2019-01-04 08:57:40
水利规划与设计(2016年9期)2017-01-15 14:00:45
水生生物学报(2015年1期)2015-02-28 16:01:05
河南科技(2014年18期)2014-02-27 14:14:54
