
基于SNP 标记分析玉米品种新科910 的遗传基础和杂种优势模式
2023-02-26 11:54:52孙佩任帅张培风张玉红王蕊王文洁张瑞平李祥马朝阳李合顺王学军周联东
中国种业 2023年2期
孙佩 任帅 张培风 张玉红 王蕊 王文洁 张瑞平 李祥 马朝阳 李合顺 王学军 周联东
(河南省新乡市农业科学院,新乡 453000)
杂种优势是选育玉米新品种的重要理论基础,合理的玉米杂种优势类群划分以及杂种优势模式研究均对于提高玉米育种效率有重要意义[1-2]。20 世纪80 年代吴景锋[3]对我国主要玉米杂交种种质基础进行评述,指出国内种质遗传基础较为狭窄,应当尽早重视。黄益勤等[4]利用RFLP 标记进行杂种优势类群划分,通过聚类分析将45 份玉米自交系划分为6 个类群,分别为热带种质类群、Mo17 类群、FRB73 类群、地方类群Ⅰ、地方类群Ⅱ和自330 或OH43 类群。曹永国等[5]利用SSR 分子标记进行聚类分析将17 个玉米自交系划分为6 个类群。张世煌等[6]将我国玉米种质划分为6 个亚群或3 个杂种优势群,DOM(国内种质、旅大红骨和塘四平头)、Reid(BSSS 和PA)和非Reid(Lancaster 和PB)种质。
分子标记技术已成为作物育种中不可缺少的手段。从最初的RFLP(Restriction fragment length polymorphism,限制性内切酶片段长度多态性)等到SNP(Single nucleotide polymorphism,单核苷酸多态性)标记,为基因定位、分子育种和基因组选择等作物育种技术奠定了技术基础[7]。1996 年Lander[8]提出SNP为新一代分子标记。SNP 技术遗传稳定性更高、可靠性更强,易于实现高通量、大规模的基因型检测[9]。
目前,SNP(单核苷酸多态性)标记已被广泛应用于玉米育种研究[10-12],包括性状的基因定位、遗传多样性分析、遗传基础分析、品种鉴定等玉米育种方面的研究[13]。在玉米杂种优势利用方面,SNP 标记也已广泛应用于划分杂种优势类群,成为构建杂种优势模式的重要手段。史亚兴等[14]利用覆盖玉米全基因组的1059 个SNP 标记,通过聚类分析,将39 份自交系划分为5 个类群,划分结果与系谱来源基本一致。卢柏山等[15]利用1031 个SNP 标记进行群体划分,将39 份甜玉米自交系划分为5 个类群。高嵩等[2]采用3750 个SNP 标记将205 份玉米自交系材料分为瑞德、兰卡斯特、旅大红骨、塘四平头、PA、PB、热带血缘共7 个类群。赵久然等[16]采用SNP 芯片技术对344份具有广泛代表性和时效性的玉米自交系进行类群划分,可分为8 个类群,分别为旅大红骨、塘四平头、Iodent、兰卡斯特、P 群、改良瑞德、瑞德和X 群。
本研究结合SNP 标记芯片技术,通过对玉米品种新科910 的双亲进行基因型鉴定和分析,对新科910 的遗传基础和杂种优势模式进行深入研究,以期为新科910 双亲后续的改良和创新奠定基础,为黄淮海地区玉米新品种的选育提供理论指导。
1 材料与方法
1.1 试验材料供试材料为玉米品种新科910 的双亲自交系(母本K381、父本H865),新科910 于2014 年通过河北省审定,审定编号:冀审玉2014011号。以45 份国内常用玉米骨干自交系作为划分杂种优势类群的参照(表1)。

表1 45 份国内常用玉米骨干自交系基本信息
1.2 SNP 分子标记分析每份材料选择5 株植株的幼嫩叶片混合取样,DNA 提取采用改良CTAB 法,并进行纯化处理。SNP 分子标记检测在北京中玉金标记生物技术股份有限公司Affymetrix Axiom 平台完成,所用芯片为玉米育种专用的中玉芯1 号(Maize10K),该芯片含有9573 个SNP 标记,均匀分布在玉米10 条染色体上,相邻标记间的物理距离平均200kb 左右。
1.3 数据统计分析用Axiom Analysis Suite V5.2 软件的apt-genotype-axiom、ps-metrics、ps-classification和apt-format-result 模块进行SNP 基因分型分析,Power Marker V3.25 软件进行最小等位基因频率、缺失率、样品杂合率和基因多样性分析,Treebest V1.9.2软件的nj-tree模型构建进化树,Admixture V1.3.0 软件进行遗传结构分析,GCTA V1.26.0 软件进行主成分分析(PCA,Principal component analysis)。
2 结果与分析
2.1 SNP 数据质量控制利用中玉芯1 号芯片对47 份供试材料进行SNP 检测,通过对dish QC>0.82和标记检出率(call rate)>0.97 的样品进行SNP 位点质控,剩余7389 个标记,占77.2%。然后进行缺失率(miss)<0.1 和最小等位基因频率(maf)>0.05过滤,最终获得4167 个可用标记,占43.5%。
2.2 群体遗传相似度分析K381、H865 与45 份我国常用玉米骨干自交系间的遗传相似系数(表2)表明,K381 与丹9046、铁7922、U8112、C8605-2、掖478、郑58 改良瑞德类群种质间的遗传相似系数变幅为0.6058~0.7513,分别为0.6058、0.6119、0.6173、0.6439、0.7248、0.7513,说明它们之间遗传距离较近。H865 与吉单27 母本、吉444、黄野四、黄早四、LX9801、昌7-2 塘四平头类群种质间的遗传相似系数变幅为0.6356~0.8556,分别为0.6356、0.7006、0.7305、0.8019、0.8109、0.8556,说明它们之间遗传距离较近。

表2 K381、H865 与45 份我国常用玉米骨干自交系间的遗传相似系数
2.3 群体聚类分析基于筛选出的SNP 标记,利用Treebest 软件的nj-tree 模型构建进化树,以45 份国内常用玉米骨干自交系作为划分杂种优势类群的参照,所有材料被划分为6 个类群,如图1 所示。K381被划分到Ⅰ类群,Ⅰ类群中有郑32、B73、U8112、铁7922、C8605-2、丹9046、郑58、K381、掖478 和先玉335 母本,该类群多属于改良瑞德类群。H865被划分到Ⅱ类群,Ⅱ类群有黄早四、黄野四、吉444、昌7-2、H865、LX9801、吉单27 母本、掖81515 和苏湾1611,该类群主要是塘四平头类群种质。Ⅲ类群有德美亚3 号母本、KWS9384 母本、德美亚2 号母本、德美亚1 号母本、先玉335 父本、德美亚2 号父本和德美亚1 号父本,该类群多属于欧洲种质。Ⅳ类群有自330、莫群17、OH43、丹340、C052、吉853、综31 和综3,该类群属于自330 和旅大红骨类群。Ⅴ类群有Mo17ht、Mo17、吉002、合344、获唐黄、德美亚3 号父本和KWS9384 父本,该类群主要是兰卡斯特种质。Ⅵ类群有沈137、齐319、X178、沈3336、豫自87-1 和吉846,该类群主要是PB 种质。
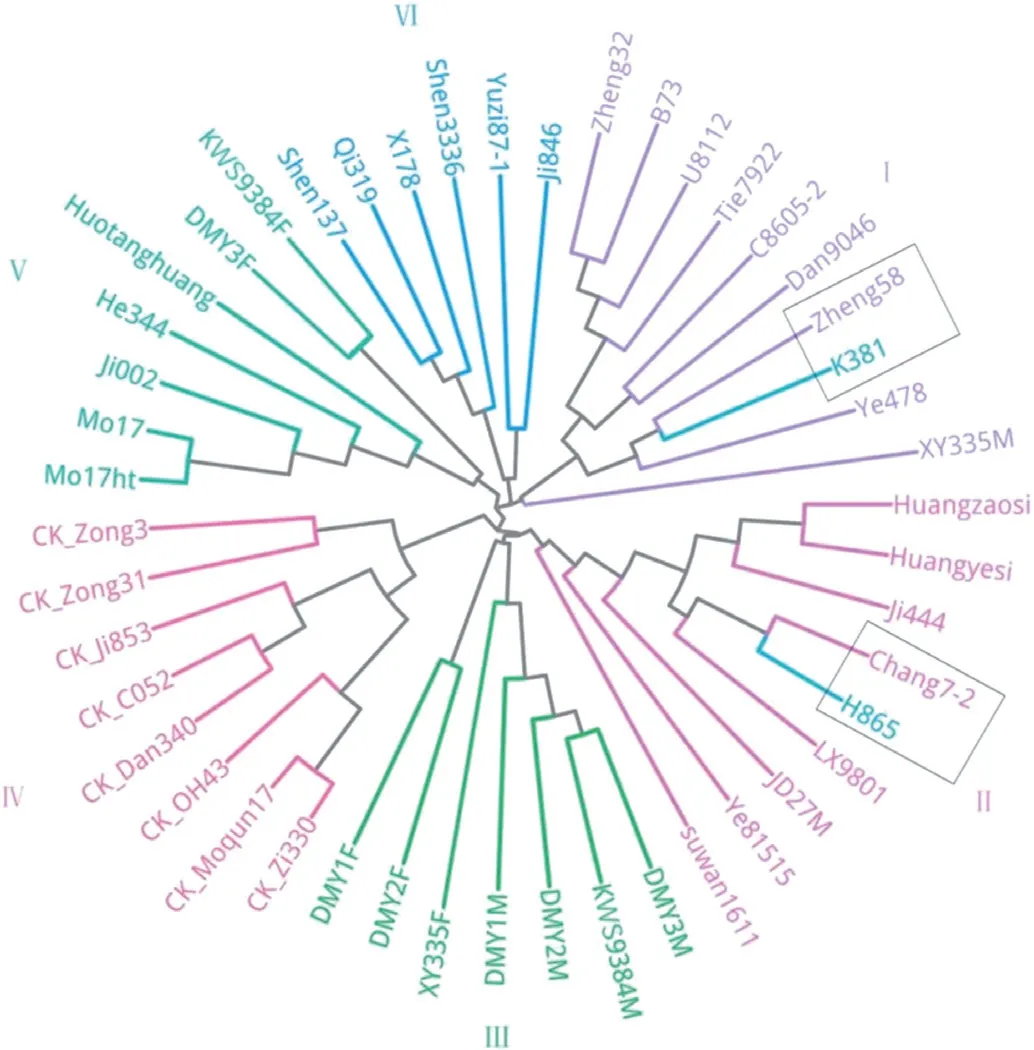
图1 以45 份我国常用玉米骨干自交系为参照构建的K381 和H865 系统进化树
2.4 主成分分析根据群体SNP 数据对供试材料进行主成分分析。前2 个主成分PC1 和PC2 贡献率分别为31.43%和26.59%,累积贡献率58.02%。PC1 和PC2 主成分的二维图(图2)显示。为了便于对比分析,图2 各自交系材料颜色与图1 一致。根据各自交系的集中程度,可清晰地鉴定出6 个类群,这与群体聚类分析结果基本一致。K381 与掖478、丹9046、C8605-2、先玉335 母本等改良瑞德类群材料距离较近,可归为一类,H865 与LX9801、昌7-2、吉444、黄野四、黄早四等塘四平头类群材料距离较近,可归为一类。
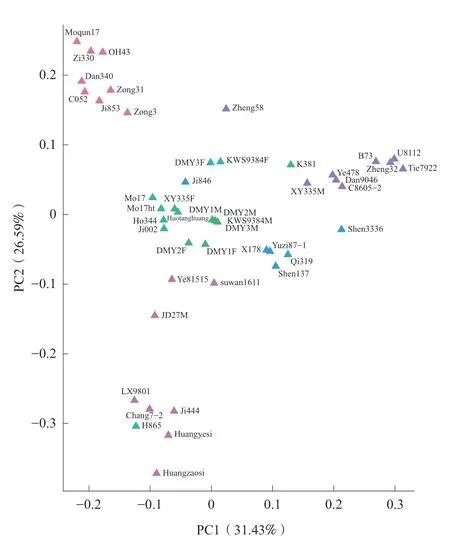
图2 二维主成分分析
2.5 群体结构分析基于筛选出的SNP 标记,利用Admixture 软件分析47 份玉米自交系材料的遗传结构,测试K=6 时最适合,47 份玉米自交系材料可分为6 个类群(图3),这与群体聚类分析和主成分分析结果基本一致。
Ⅰ类群中有德美亚2 号父本、KWS9384 父本、德美亚3 号父本、获唐黄、Mo17ht、吉002、合344 和Mo17,该类群主要是兰卡斯特种质。K381 被划分到Ⅱ类群,Ⅱ类群有先玉335 母本、K381、C8605-2、掖478、丹9046、B73、U8112、郑32 和铁7922,该类群主要是改良瑞德类群种质。H865 被划分到Ⅲ类群,Ⅲ类群有德美亚1 号父本、苏湾1611、掖81515、吉单27 母本、黄野四、LX9801、黄早四、H865、吉444 和昌7-2,该类群主要是塘四平头类群种质。Ⅳ类群有吉846、沈3336、豫自87-1、X178、沈137 和齐319,该类群主要是PB 群种质。Ⅴ类群有先玉335 父本、德美亚1 号母本、KWS9384 母本、德美亚3 号母本和德美亚2 号母本,该类群属于欧洲种质。Ⅵ类群有郑58、综3、综31、丹340、OH43、吉853、C052、自330 和莫群17,该类群属于自330 和旅大红骨。表3 为群体遗传结构分析所得的6 类群血缘在新科910 双亲中占有的比例。由图3和表3 可以看出,K381 遗传背景相对复杂,属于改良瑞德类群,H865 遗传背景比较单纯,属于塘四平头类群。

表3 K381 和H865 的群体结构分析
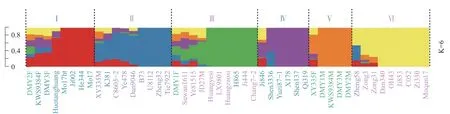
图3 47 份自交系的群体结构分析
3 结论与讨论
本研究以45 份国内常用玉米骨干自交系作为划分杂种优势类群的参照,分析玉米品种新科910的双亲自交系(母本K381、父本H865)的遗传基础。基于SNP 标记出现的频率计算材料间的遗传相似系数,分析材料间的遗传相似程度,为分群提供依据。同一血缘材料的遗传相似系数一般会在0.60或0.70 以上,0.50 左右或者以下的遗传相似系数一般代表材料间没有明显的血缘联系[17]。遗传相似度分析结果表明:K381 与掖478(0.7248)、郑58(0.7513)等改良瑞德类群材料之间的遗传相似系数较高,遗传距离较近。H865 与LX9801(0.8109)、昌7-2(0.8556)等塘四平头类群材料之间的相似系数较高,遗传距离较近。
玉米主要利用的杂种优势模式有瑞德×塘四平头、兰卡斯特×旅大红骨等[18-19]。本研究群体聚类分析结果表明,47 份玉米自交系材料被划分为6 个类群,利用SNP 标记对新科910 双亲K381 和H865 进行群体遗传相似度分析、聚类分析、主成分分析和遗传结构分析,K381 属于改良瑞德类群,父本H865 属于塘四平头类群,新科910 的杂种优势模式为改良瑞德×塘四平头,针对近年来黄淮海地区异常灾害天气的频繁发生[20],下一步将会引进新的种质材料,以此杂种优势模式为理论依据,对新科910 的双亲进行改良和创新,为抗生物胁迫与非生物胁迫玉米新品种的选育奠定基础。
猜你喜欢
Chinese Physics B(2022年11期)2022-11-21 09:28:30
蔬菜(2021年7期)2021-11-27 03:45:25
海峡科技与产业(2021年1期)2021-05-22 01:26:20
北京农学院学报(2019年1期)2019-02-22 05:39:46
丝绸(2018年1期)2018-01-25 01:00:28
河南农业科学(2017年4期)2017-04-12 09:55:45
西南农业学报(2016年5期)2016-05-17 05:42:20
西南农业学报(2016年6期)2016-04-16 05:12:47
山西农经(2016年5期)2016-02-28 14:24:37
茶叶通讯(2014年2期)2014-02-27 07:55:39
