
Hormetic effects of abiotic environmental stressors in woody plants in the context of climate change
2023-02-17 05:49ElenaErofeeva
Journal of Forestry Research 2023年1期
Elena A.Erofeeva
Abstract Woody plants contribute to the stability and productivity of terrestrial ecosystems and are significantly affected by climate change.According to the concept of environmental hormesis,any environmental stressors can cause hormesis,that is,stimulation in low doses and inhibition in high doses.Numerous studies have demonstrated plant hormesis under low doses of various abiotic stressors.However,the hormetic responses of woody plants to abiotic stressors from climate change are insufficiently studied.This review analyses data on the stimulating effects of low doses of climate stressors in experiments and in real ecosystems.Numerous laboratory and field experiments show that single and combined exposure to various climate stressors (temperature,humidity,and elevated carbon dioxide concentrations) can cause hormesis in various species and functional types of woody plants,which can be accompanied by hormetic trade-offs and preconditioning.In addition,there is evidence of climate hormesis in woody plants in ecosystem conditions.Field experiments in various ecosystems show that elevated temperatures and/or precipitation or elevated carbon dioxide concentrations causing hormesis in dominant tree species can stimulate ecosystem productivity.Moreover,climate hormesis of the growth and reproduction of dominant forest tree species contributes to the spread of forests,that is,climate-driven ecological succession.The main commonalities of climate hormesis in woody species include:(1) Low-dose climate stressors cause hormesis in woody plants when strong (limiting) stressors do not affect plants or these limiting stressors are mitigated by climate change.(2) Hormesis can occur with the direct impact of climatic stressors on trees and with the indirect impact of these stressors on plants through other parts of the ecosystem.(3) Climate stressor interactions (e.g.,synergism,antagonism) can affect hormesis.(4) Hormesis may disappear due to tree acclimatization with consequent changes in the range of tolerances to climate factors.This review highlights the need for targeted studies of climate hormesis in woody species and its role in the adaptation of forest ecosystems to climate change.
Keywords Abiotic stress·Ecological succession·Ecosystem·Forest·Hormesis
Introduction
In 2021,the Intergovernmental panel on climate change (IPCC 2021) reported that CO2concentrations were about one and a half times higher than before the industrial revolution,and the average temperature between 2011 and 2020 was 1.09 °C higher than during the pre-industrial period.Climate change,which is considered to be caused primarily by anthropogenic greenhouse gas emissions,poses a serious challenge to ecosystems due to the negative impact on ecosystem structure,species ranges and phenology (IPCC 2022;Xiao et al.2022).In terrestrial ecosystems,woody plants play an important role in ensuring the stability and productivity and are significantly affected by climate change (Buriánek et al.2013;Fatholahi et al.2017;Njana et al.2021).
Increasing experimental evidence has shown hormetic effects on plants of various stressors in low doses (Agathokleous et al.2022a;Erofeeva 2022c),including abiotic factors such as temperature,humidity that are directly related to climate change (Agathokleous et al.2019;Erofeeva 2021).This evidence suggests that climate change causes hormetic stimulation in woody plants.At the same time,despite extensive data on the climate effects on various trees (Friedrichs et al.2009;Gedalof and Berg 2010;Devi et al.2020;Kijowska-Oberc et al.2020;Li et al.2020;Heilman et al.2021),there have been no reviews on climate hormesis in woody plants based on data obtained both in experiments and in real ecosystems.
In the present review,we thus (1) analyze experimental evidence and patterns of hormetic responses of woody plants to low-dose abiotic stressors related with climate change;(2) examine the evidence showing climate hormesis of woody plants,obtained from natural ecosystems using dendrochronological analysis and with anthropogenic climate change in urbanized areas;(3) consider the impacts of climate hormesis in woody plants on ecosystems and consequently on ecological successions;and (4) analyze general patterns of climate hormesis in woody plants in conjunction with the effects of climate stressors.We also analyzed plant hormesis caused only by the most important climate stressors (temperature,humidity and CO2concentration),for which there is clear evidence of a relationship with climate change.
Hormesis in woody plants under experimental exposure to abiotic stressors related with climate change
Climate change affects woody plants mainly by disrupting the annual course of temperature and precipitation,which,in turn,depend on the atmospheric concentration of greenhouse gases,in particular CO2(IPCC 2021;IPCC 2022).Any low-dose stressor can cause hormesis,a positive,stimulating response,in living systems,including woody plants (Agathokleous 2018;Erofeeva 2022a,b),in contrast to the damaging effects of high doses of the stressor (Agathokleous and Calabrese 2022) (Fig.1).Hormesis drives biological modifications from cells to higher levels of biological organization and has emerged as a basic biological feature (Agathokleous and Calabrese 2022;Erofeeva 2022c).To date,there is significant evidence of biphasic hormetic responses of various indicators in plants with low-dose stressors having both a zero control level (e.g.,pollutants such as herbicides,heavy metals,micro-and nanoplastics) (Agathokleous et al.2021,2022b;Belz et al.2022) and an optimal control level (e.g.,abiotic factors such as temperature,light,humidity),the effects of which are described by Shelford’s tolerance law curve (Agathokleous et al.2019;Erofeeva 2021).In 2019,Agathokleous et al.demonstrated sufficient evidence of temperature hormesis in plants,including in woody species,and I analyzed the evidence for plant hormesis caused by various abiotic factors (light,temperature,soil humidity,CO2air concentration) (Erofeeva 2021).However,a detailed analysis of hormetic reactions of woody plants to experimental exposure to climate-related abiotic stressors has not been carried out.
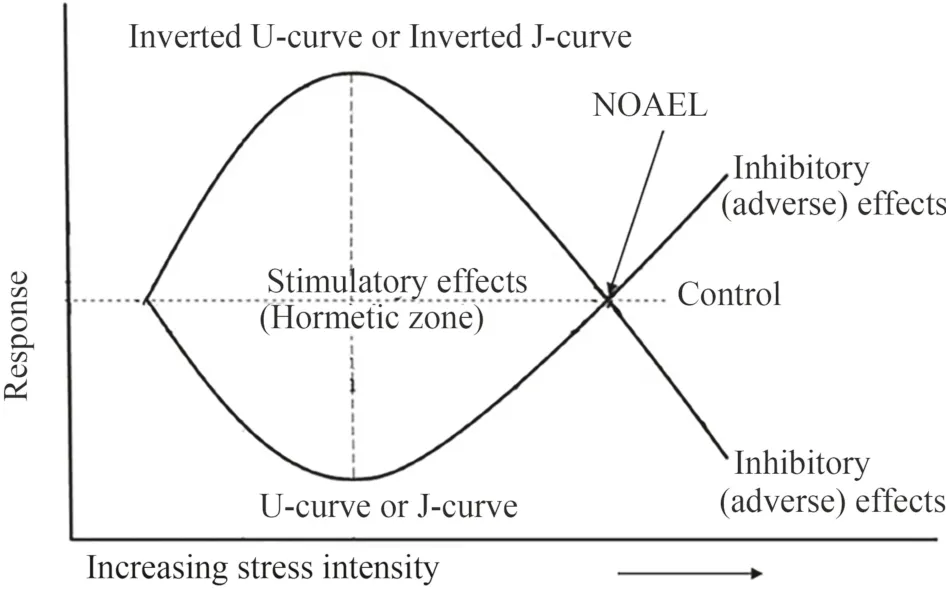
Fig.1 Two types of hormetic dose-response curves (modif ied from Erofeeva 2022b,c).The inverted U-curve (or inverted J-curve) demonstrates low-dose stimulation and high-dose inhibition and is the most frequently observed.The U-curve (or J-curve) demonstrates less damage at low doses and more damage at high doses.NOAEL,the no-observed adverse-effect level,is the maximal dose of a stressor that does not cause any damage.The doses in the hormetic zone have stimulatory effects;the maximal stimulating effect (MAX) is typically 130-160% of the control.These stimulatory effects improve the condition of the living system,including plants,compared to the control due to overcompensation (Calabrese 2008;Agathokleous and Calabrese 2020;Erofeeva 2022b,c)
Currently,a significant number of chamber and field experiments demonstrate hormetic stimulation in woody plants under exposure to various abiotic stressors related with climate change (Table 1);I use the term climate hormesis to describe these stimulations.Climate hormesis has been detected at higher CO2concentrations (Ainsworth and Long 2005;Centritto et al.2008;Allen and Vu 2009;Yang et al.2020),soil moisture (de Dato et al.2008;Motai et al.2017),and air temperatures (Apple et al.2000;Yuan et al.2018;Li et al.2020).Interestingly,in the vast majority of cases,climate hormesis has been found when abiotic stressors have exceeded the optimal (i.e.,control level).


Hormesis might occur not only with an excess,but also at lower levels of an abiotic factor (Calabrese and Agathokleous 2020;Erofeeva 2021).For example,de Dato et al.(2008) reported that moderate drought led to an increase in the aboveground standing woody biomass and N content in evergreen woody shrubs (Table 1).Hormesis caused by several low-dose stressors simultaneously is considered to be much more common in ecosystems compared to the hormetic effects of single stressors,but has been studied less (Agathokleous et al.2022b).Some studies have found climate hormesis in woody plants with a combination of various stressors (increased CO2+elevated air temperature) and synergism,that is,a significant increase in hormetic stimulation with a combination of these stressors compared to their isolated effects (Callaway et al.1994;Overdieck et al.2007;Usami et al.2008;Zhao et al.2012) (Table 1).Synergism of hormesis-inducing stressors has been reported in deciduous angiosperm trees such asPopulus cathayana(Zhao et al.2012),Quercus myrsinifolia(Usami et al.2008),andFagus sylvatica(Overdieck et al.2007) and in an evergreen conifer (Pinus ponderosa) (Callaway et al.1994).
Climate hormesis has also been reported for woody plants of various functional types (deciduous and evergreen angiosperm trees,evergreen conifers,evergreen angiosperm shrubs),in evolutionarily remote groups of woody plants,such as gymnosperms (e.g.,Pinus ponderosa,Picea sitchensis;Callaway et al.1994;Centritto et al.2008)and angiosperms (e.g.,Populus cathayana,Betula pendula) (Kuokkanen et al.2004;Zhao et al.2012),at various developmental stages (seedlings,vegetative stages,mature trees),and growing in different climate zones (temperate,boreal,subtropical,tropical) (Table 1).This universality of climate hormesis in woody species is apparently due to the general (nonspecif ci) mechanisms underlying the hormetic responses to climate abiotic stressors in various plant species throughout their lives.The nonspecific defense mechanisms (i.e.,universal for various plant species and their reactions to any stressors) known to have a significant role in plant hormesis (Agathokleous et al.2020;Erofeeva 2022a,b) include the synthesis of antioxidant defense (enzyme and non-enzyme antioxidants) and stress proteins,induced by low-dose stressors (Agathokleous et al.2020;Erofeeva 2022a,b).However,the role of nonspecific protective mechanisms in the hormetic responses of woody plants to climatic stressors is poorly understood.
Along with the nonspecific mechanisms,specific effects induced by feature(s) of the stressor at low doses are also essential for plant hormesis (Erofeeva 2022a),including climate hormesis in woody species.For example,increased atmospheric C O2causes the so-called CO2fertilization effect in trees,that is,increased rate of photosynthesis with limited leaf transpiration (Ueyama et al.2020;Smith et al.2015).In fact,CO2fertilization is a hormetic response,since a moderate increase in CO2content stimulates photosynthesis and growth (Ainsworth and Long 2005;Smith et al.2015;Ueyama et al.2020),and high concentrations of CO2suppress them (Reuveni and Bugbee 1997;Zheng et al.2019).Examples of hormetic stimulation by increased CO2are presented in Table 1.Elevated CO2concentrations specifically affect photosynthesis,which is essential for their effects.The partial pressure of CO2is an important limiting factor for photosynthesis since CO2reacts with ribulose-1,5-bisphosphate carboxylase-oxygenase (Rubisco) during photosynthesis to form carbohydrates (carbon fixation) (Huang et al.2007).
In addition,hormetic effects of climate stressors can be species-specific.For example,in an experimental warming study,annual biomass accumulation (g m-2a-1) over three years increased in the evergreen angiosperm shrubsCistus monspeliensisandPistacia lentiscus(growth stimulation) but decreased inCistus creticus(growth inhibition) compared to the control (de Dato et al.2008).In addition,functional types of trees can differ in the effect of climatic hormesis.For instance,in an analysis of 68 field and pot studies,elevated temperatures enhanced the growth of deciduous angiosperm species more than that of evergreen angiosperm trees (Way and Oren 2010).Moreover,evergreen tropical trees were more susceptible than temperate or boreal trees to growth declines induced by warming.The authors attribute these results to the different ecological strategies of trees from different biomes.The evergreen strategy is most common in sites with low nutrient or water availability.Therefore,evergreen species are less responsive to environmental changes because they have traits that limit nutrient loss;since there is a trade-offbetween nutrient retention and the ability to maximize productivity,evergreen species are constrained in their ability to increase growth when conditions (such as temperature or nutrient availability) become favorable.In addition,a meta-analysis of studies on CO2enrichment in free air showed that trees are more sensitive than other functional types of woody species such as shrubs to stimulating effects of CO2fertilization (Ainsworth and Long 2005).In addition,photosynthesis of some deciduous trees has also been found to be more sensitive to elevated CO2than that of conifers (Huang et al.2007).Thus,the ecological strategy of woody species affects the outcome of climate hormesis.
In woody plants,as well as other plant species,hormetic trade-offs have been found.Hormetic trade-offs include situations when the same dose of a low-dose stressor stimulates some endpoints and worsens others in the living system (trade-off1) or some indicators are stimulated as others are unchanged (trade-off2) (Erofeeva 2022a).For example,in some species of trees,elevated C O2concentrations increase the rate of photosynthesis,but decrease frost resistance (Barker et al.2005) (trade-off1),and in others,growth is not affected (DeLucia et al.1994;Lovelock et al.1999) (trade-off2).
Abiotic factors can also induce hormetic preconditioning in woody plants,whereby a low-dose stressor increases the resistance of living systems to subsequent more severe effects of this stressor or other strong stressors (Agathokleous et al.2020;Erofeeva 2022b,c).The organism can then better cope with environmental instability and new challenges (Agathokleous and Calabrese 2020;Erofeeva 2022b,c).For example,increased CO2with elevated temperature stimulate the growth of birch and enhance the resistance of stems to damage by hares due to a significant increase in papyriferic acid (PA) (Kuokkanen et al.2004),a toxic triterpene that causes indigestion and nausea in mammals.PA is secreted by glands on twigs of birches,inhibits the enzyme succinate dehydrogenase in gastrointestinal cells and decreases mitochondrial energy production in mammals (McLean et al.2009).Moreover,abiotic stressors can have specific effects on hormetic precoditioning.For example,elevated CO2increases the resistance of woody plants to drought because it reduces stomatal conductivity and transpiration (i.e.,increases water-use efficiency [WUE];Huang et al.2007;Gedalof and Berg 2010;Park et al.2020).WUE is the amount of carbon assimilated as biomass or seeds produced per unit of water used by plants (Park et al.2020).Thus,the specific impact of climatic stressors is important for hormetic preconditioning to strong stressors and thus for increasing plant productivity.
Hormesis in woody plants exposed to climate stressors in ecosystems
Most evidence for hormetic responses of woody plants to climate stressors has been obtained in experimental settings (Table 1).In real ecosystems,however,woody species are exposed to significantly more stressors simultaneously.In addition,climate change can indirectly impact plants through other parts of the ecosystem.Therefore,data on hormetic reactions of woody plants to climate changes obtained in real ecosystems are of critical importance.In addition,urban areas create their own climate patterns by altering physical properties such as evapotranspiration,albedo,emissivity,anthropogenic heat flux,wind speed,and air pollution (Vinayak et al.2022;Schneider et al.2022).Urban heat islands (UHIs),which have higher temperatures than surrounding non-urban areas,are the most evident example of local climate modification induced by urbanization (Vinayak et al.2022).In the inner city,the diurnal cycles of air temperature differ from those in the surrounding area,and typically,minimum air temperatures at night are much higher in the city (Schneider et al.2022).Urbanization-induced warming can reach up to 4 °C more than in non-urban,and urbaninduced climate changes are on the same order of magnitude as greenhouse gas-induced climate change (Vinayak et al.2022).
Climate change in urban areas can also stimulate effects in woody plants relative to the trees in surrounding areas,although the actual climate effects are masked by anthropogenic impacts and are difficult to isolate (Schneider et al.2022).For instance,based on tree ring analyses in 10 metropolises worldwide,Pretzsch et al.(2017) showed that urban trees tend to grow 14-25% faster than their counterparts in the rural surroundings.At the same time,the stimulation of tree growth depends on the climate zone.Stimulatory effects have been found for trees in cities of boreal (Tilia cordataMill.,Aesculus hippocastanumL.,Platanus×hispanicaMill.ex Münchh.) and subtropical (Robinia pseudoacaciaL.,Quercus roburL.) climate zones based on data collected over several decades before and/or since 1960 (Pretzsch et al.2017).This stimulation could be due to elevated air temperature by extending the growing season,a decrease in diurnal temperature range,and higher atmospheric CO2concentrations in urban areas compared to the rural areas (Pretzsch et al.2017).However,these effects were not found for cities in the Mediterranean climate zone.In addition,urban trees (Abies sachalinensis,Picea glauca) grow significantly slower than rural trees in the temperate climate zone,perhaps due to lower water availability compared to the subtropical and boreal zones,which receive sufficient rainfall.
Searle et al.(2012) reported an eightfold increase in the biomass of 2-year-old seedlings ofQuercus rubrain Central Park in New York City compared to seedlings in rural areas,and the root-to-shoot ratio was lower (urban ≈ 0.8,rural ≈ 1.5).The urban seedlings also had greater leaf growth,increased photosynthetic area (by 10 times) and nitrogen concentration in the leaves (by 23%) than seedlings in rural areas.A laboratory experiment conf irmed that these stimulating effects are most likely due to the increased air temperature in the city relative to rural areas (difference in average maximum temperatures of 2.4 °C and 4.6 °C in minimum temperatures) (Searle et al.2012).Growth of aPopulus deltoideswas similarly stimulated in urban (New York City) and rural areas,but biomass doubled in the urban trees compared to the rural (Gregg et al.2003).
In a study of the traits of 11 common woody plants (trees and shrubs) along an urban-rural gradient in Beijing,China,leaf size,specific leaf area,and leaf nitrogen and potassium became significantly lower moving from the urban to the rural,indicating that urbanization can enhance resource acquistion for growth and production (Su et al.2021).Soil nutrients and air temperature also decreased along the urban-rural gradient,while air relative humidity increased.A structural equation model showed that these alterations in physical factors may affect plant growth and production because of the improvement in resource acquisition capacity.
In addition,in forest and forest-tundra ecosystems,dendrochronological analyses of the radial growth of various tree species have revealed an increase in this indicator and its relationship with climate change in various regions.For example,Devi et al.(2020) analyzed data for 115 years and showed that elevated temperature and precipitation increased the radial growth of spruce (Picea obovataLedeb.) and larch (Larix sibiricaLedeb.) at the upper treeline ecotone in the mountains of the Polar Urals (continental subarctic climate).This finding was corroborated by positive correlations between growth and the two climate indicators.Studies in other subarctic regions showed a clear increase in the productivity of stands in the second half of the Twentieth century,but with a recent decrease in the rate of biomass accumulation due to the aging of stands (Bunn et al.2007).
In 2014,Pretzsch et al.(2014) reported that growth of the dominant tree species,Norway spruce [Picea abies(L.) Karst.] and European beech (Fagus sylvaticaL.),in the oldest experimental forest plots in Central Europe,was significantly faster (by 32-77%) than in 1960.Statistical analyses of the experimental plots and application of an ecophysiological model showed that the rise in temperature and extended growing seasons were the main contributors to the accelerated growth,especially in fertile areas.Similarly in 2014,Kauppi et al.(2014) reported a significant increase in annual growth of boreal forests in Finland since 1960 in response to climate warming.Their regression analysis showed a strong positive relationship between growing degree-days (GDD) (i.e.,the annual sum of daily temperatures above a predef ined threshold) and tree growth (m3ha-1a-1).
Caignard et al.(2017) found that tree reproduction has already been altered by climate change in temperate areas;reproductive effort (seed production) of temperate oaks has significantly increased with increasing spring temperature over the last decade.In contrast,no relationship was found between seed production and precipitation.Thus,climate change can stimulate not only growth,but also the efficiency of seed reproduction in populations of woody plants.
Because woody plants adapt to their climate,climate does not cause significant stress,exception in the ecotone zone (the transition zone between different ecosystems or biomes),where many environmental factors deviate significantly from the optimal for tree growth (Heilman et al 2021),causing stress on tree species.Consequently,climate stimulation of the tree traits described above are not caused by a shift in the climate factors to optimal values,but is due to climate hormesis.Thus,the facts discussed above demonstrate the ability of climate stressors to cause hormetic stimulation of growth and productivity of tree species in natural and urban ecosystems.However,more detailed and targeted studies of climate hormesis in real ecosystems are required,since the studies described above were not aimed at assessing the hormetic effects of climate and provide mainly indirect evidence of climate hormesis in woody plants.
The impacts of climate hormesis in woody plants on the state of ecosystems
Woody plants are the basis for some terrestrial ecosystems (e.g.,forest,forest tundra) since they are the main source of primary production,edificators (i.e.,species with habitatformation ability) and have an important role in the biogeochemical cycling of water,carbon and nutrients and flow of energy and are critical for the functioning,productivity and stability of ecosystems (Buriánek et al.2013;Fatholahi et al.2017).Therefore,the hormetic stimulation of woody plants caused by climate change can significantly affect ecosystems,but no studies have targeted this issue.Nevertheless,analyses of the literature have shown a lot of indirect data regarding this question in various types of ecosystems.
Several meta-analyses,which examined data from experimental climate changes in the field over several years,revealed stimulating effects on various dominant woody species and related ecosystem indicators.For instance,Wu et al.(2011) analyzed the results from 85 studies of various biomes,including those dominated by woody species (boreal forest,moist tropical forest,forest-steppe,among others) over 5 years and found that experimental warming and increased precipitation generally stimulated aboveground biomass (on average by 27% and 12%,respectively) of woody plants and ecosystem C fluxes.In addition,experimental warming significantly stimulated total net primary production (TNPP) and increased ecosystem photosynthesis and respiration (on average by 15%,20% and 27%,respectively).Experimentally increased precipitation has enhanced TNPP,net ecosystem exchange,ecosystem photosynthesis,and ecosystem respiration (on average by 28%,56%,40% and 30%,respectively),whereas reduced precipitation suppressed these indicators.The magnitude of responses of these ecosystem indicators to the experimental climate change systematically varied little with climate,indicating general sensitivities across ecosystems to the climate-change treatments.
In the 2001 meta-analysis of similar data from from 32 studies of forest ecosystems exposed to 2-9 years of experimental warming (0.3-6.0 °C),Rustad et al.reported that warming significantly increased aboveground plant productivity (annual production of biomass and litter per unit ground area) by 19%,soil respiration rates by 20%,and net N mineralization rates by 46%.Aboveground plant productivity increased more response to warming in colder ecosystems than warmer.Another meta-analysis,based on 36 experiments,revealed a high sensitivity of the biomass of Arctic deciduous and evergreen shrubs to experimental warming,expressed as an increase in biomass of 30%-50% compared to the control (Dormann and Woodin 2002).Increases in atmospheric CO2have also been found to enhance the productivity of various ecosystems in which woody species predominate.For instance,Smith et al.(2000),using free-air CO2enrichment (FACE) technology in an intact Mojave Desert ecosystem have reported that new shoot production of a dominant perennial shrub is doubled by a 50% increase in atmospheric CO2concentration in a high rainfall year.However,elevated CO2does not enhance production in a drought year.
These meta-analyses,based on extensive data,showed hormetic stimulation of ecosystem indicators as a result of exposure to low doses of climate stressors on woody plants in various climate zones and ecosystems.These facts indicate the universality of hormetic reactions in various types of ecosystems,similar to the manifestation of hormesis at the level of individuals and populations (Agathokleous 2018;Erofeeva 2022c).In addition,the meta-analyses revealed moderate stimulating effects at the ecosystem level (Fig.2) similar to the hormetic responses of individuals and populations (Agathokleous 2018;Erofeeva 2022c).The hormetic stimulation is usually 110-160% relative to the control (Calabrese 2008).

Fig.2 Induction of climate hormesis in woody plants and consequences.Climate hormesis can occur through the direct impact of climate stressors on woody species and the indirect effects of these stressors on plants via other parts of the ecosystem.Climate hormesis of dominant woody species can significantly stimulate ecosystem indicators (e.g.,ecosystem exchange,carbon flux) and increase eco-system stability and productivity.Climate stimulation of growth and reproduction of dominant forest trees can also contribute to forest expansion (i.e.,climate-driven ecological succession).Low-dose climate stressors can cause hormetic preconditioning to strong climate stressors contributing to the acclimatization of woody species under changing conditions
Hormetic stimulation of forest ecosystem productivity can significantly affect geochemical cycles,including reducing atmospheric carbon by increasing its absorption,which can mitigate climate change caused by anthropogenic CO2emissions.Thus,the study of climatic hormesis impacts on the stability and productivity of ecosystems is important for assessing and predicting the effects of climate change.
Climate hormesis in woody plants and ecological successions
Agathokleous (2018) suggested that abiotic hormesis in plants can lead to changes in the composition of communities and becomes the driving force of ecological succession.To date,more and more evidence indicates that climate hormesis in woody plants is essential for ecological succession processes.
Global warming is most pronounced at high latitudes,where temperatures over the last 30 years have increased by 0.6 °C per decade,twice as fast as the global average (Devi et al.2020).Therefore,such areas are most suitable for assessing the stimulating effects of warming in relation to woody plants,for which the ecotone of the area is often located in these latitudes.For example,Devi et al.(2020) reported that stimulation of radial growth and stand density in spruce (Picea obovata) and larch (Larix sibirica) due to the increase in air temperature and precipitation in the Polar Urals was accompanied by a 50-m upward elevational shift during the observation period (since 1900).Treeline advances in polar regions are mostly limited by temperature (Devi et al.2020).This study in the ecotone of spruce and pine habitats showed that warming stimulates tree growth and leads to the expansion of the species’ range.Thus,climate change can reduce the intensity of limiting stressors (e.g.,low temperatures),so that temperature turns from a strong stressor into a mild stressor.Growth,development,reproduction and competitiveness of spruce and larch are stimulated,and the tundra is gradually replaced by forest tundra.
In addition,Silva et al.(2016) found stimulating effects of several climate-related abiotic factors on tree growth and the spread of alpine forests in the ecotone area.Dendrochronological analyses at the upper limit of alpine forests in the Tibetan Plateau revealed a steady increase in tree growth since the early 1900s,which intensif ied during the 1930s and 1960s,reaching unprecedented levels since 1760.This growth acceleration has been observed in young and old trees and coincides with the establishment of trees outside the forest range,reflecting a connection between the physiological performance of dominant species and shifts in forest distribution.Measurements of stable isotopes (carbon,oxygen,and nitrogen) in tree rings indicate that tree growth was stimulated by the synergistic effect of rising atmospheric CO2and a warming-induced increase in water and nutrient availability as the permafrost thawed (Silva et al.2016).Thus,climate hormesis can contribute to ecological succession,including the expansion of forests (Fig.2).Detailed study of this issue is a task for the future.
The general patterns of hormesis in woody plants with combined effects of climate stressors
Climate change leads to changes in a set of abiotic factors (e.g.,temperature,precipitation,light,soil nitrogen) (Silva et al.2016;Devi et al.2020) and can affect plants in combination with anthropogenic stressors such as increased CO2in the atmospheric air,nitrogen deposition (Pretzsch et al.2017;Schneider et al.2022).Thus,climatic hormesis in woody plants is almost always a reaction not to one stress factor,but to a set of stressors directly or indirectly related to climate change,so the general patterns of hormetic reactions of plants need to be studied under the combined effects of climatic stressors.
To date,based on the analysis of the literature in the previous sections,the following patterns of climatic hormesis can be distinguished with combined effects of climate stressors:
(1) Limiting environmental factors (i.e.,any strong stressors) have a negative impact on organisms,even if other environmental factors are favorable (Odum 2004;Erofeeva 2021).This pattern is also evident for the hormetic effects of climate stressors.A significant number of studies have shown that climate stressors in low doses have stimulating effects on woody plants only in the absence of any strong stressors,that is,limiting environmental factors (Peñuelas et al.2007;de Dato et al 2008;Pretzsch et al.2017).For example,the increased temperature has stimulated the growth of trees in field experiments (de Dato et al 2008) and in natural ecosystems (Pretzsch et al.2017) only in the absence of restrictions on water supply.At the same time,if climate change decreases the intensity of the limiting abiotic factors to that of low-dose stressors,then a stimulatory effect occurs as in the case of woody species of high latitudes (Arctic and sub-Arctic climatic zones),where low temperature is a limiting factor (Rustad et al.2001;Devi et al.2020).However,if climate change creates strong abiotic stressors,then the effect will be inhibitory.For instance,experimental warming in the field led to decreased tree biomass and photosynthesis due to warming-induced moisture stress (Wu et al 2011).
(2) Environmental stressors can have both direct and indirect effects on organisms through changes in other environmental factors (Odum 2004).Consequently,the stimulating effects of low-dose climate stressors can be realized not only by direct impact on woody plants,but also indirectly due to changes in other parts of the ecosystem (Fig.2).For instance,climate warming can increase soil respiration,including in forest ecosystems (Rustad et al.2001),which may be a hormetic stimulation of soil microbial communities as shown for various low-dose stressors (Wang et al.2021;Agathokleous et al.2022c).In turn,the rate of mineralization of organic compounds in the soil and the amount of nitrogen available to plants increase,enhancing the growth and biomass of trees (Rustad et al.2001;Dormann and Woodin 2002;Melillo et al.2002 ;Wu et al.2011),especially in many mid-latitude forests where plant growth is nitrogen-limited (Melillo et al.2002).Thus,the eventual effect of climate change depends on which parts of the ecosystem are stimulated/inhibited with the woody species,potentially generating not only a positive,but also a negative impact on tree species.For example,climate warming might increase the number of tree pests (i.e.,a severe biotic stress),which would lessen any increase in tree biomass caused by the elevated temperature (Reyer et al.2017).However,these issues require further research.
(3) Interactions among environmental factors/stressors have long been known to impact different organisms (Odum 2004) through the additivity of multiple stressors (i.e.,the sum of their effects),synergism (the stressors together have greater effect than individual) or antagonism (lower combined effect than individual effects) (Piggott et al 2015).Examples of the synergism of elevated temperature and CO2during hormesis in woody species are presented in Table 1 and were discussed above.Thus,the interaction of stress factors is determined by the mechanisms underlying their physiological effects and of great importance for climate hormesis in woody plants.
(4) The stimulating effects of multiple climate stressors on woody species may change over time,mainly from acclimatization of the species.As a result,the optimum for the climate factor may shift,then new climate conditions will not cause a hormetic effect.Epigenetic memory also plays an essential role in the acclimatization of woody plants (Yakovlev et al.2016;Kijowska-Oberc et al.2020;Trontin et al.2021).
Epigenetic memory is the stable inheritance of changes in gene expression through mitosis and/or meiosis.The same tree genotype may produce different epigenomes during its lifespan to cope with different environmental cues.Alternatively,during its life,an individual tree may produce large numbers of offspring with different epigenomes.Consequently,each generation of seeds can receive different epigenetic “priming”,which is then subjected to natural selection.Thus,trees create seed banks that “store” these various epigenetic “primings”,thereby creating an “ecological epigenetic memory” that provides epigenetic responses to a wide range of environmental stressors (Maury et al.2018).
The role of hormesis in acclimatization,climate epigenetic memory,and the epigenetic mechanisms of climatic hormesis have yet to be studied in woody plants.However,data obtained for animals indicate the involvement of epigenetic mechanisms in transgenerational effects of hormesis including stimulatory effects of low-dose abiotic stressors (Agathokleous et al.2022d).Perhaps preconditioning by climate hormesis prepares trees for severe climate stressors and includes the formation of an epigenetic memory,which can become the basis for altering the development program in the present or subsequent generations and,consequently,change the tolerance range of the species and contribute to its acclimatization.
Conclusion
Numerous laboratory and field experiments have demonstrated hormetic responses in woody species to various abiotic climatic stressors such as temperature,humidity,and elevated CO2concentrations,which can have isolated and combined effects.Climate stressors can cause hormesis in various species and functional types of woody plants,and at different stages of development (seedlings,vegetative stages,mature trees) in terms of growth,productivity,biochemical and physiological indicators.In addition,hormetic trade-offs and preconditioning have been reported.
Climate hormesis is present in woody plants in various ecosystems.In urban ecosystems,anthropogenic climate change can stimulate the growth and productivity of woody plants over those growing outside the city in various climate zones (temperate,boreal,subtropical).In addition,based on dendrochronological analysis of the radial growth of dominant tree species over a long period (from several decades to 115 years),an increase in radial growth was related to climate changes (elevated air temperature and/or precipitation) in boreal,temperate forest and subarctic forest tundra ecosystems.
Hormetic stimulation of dominant woody species caused by climate change can signif ciantly affect the state of an ecosystem.Field experiments performed in various ecosystems (boreal forest,moist tropical forest,forest-steppe,Arctic and desert ecosystems) have shown that exposure of dominant trees to elevated temperature and/or precipitation or elevated CO2concentrations can stimulate ecosystem productivity.Other studies have found that climate stimulation of the growth and productivity of dominant forest trees contributes to climate-driven ecological succession.
Analysis of the patterns of climate hormesis in woody species has shown that hormetic stimulation by climatic stressors occurs when there are no limiting (strong) stressors or they are mitigated as a result of climate change (i.e.,limiting stressors become stressors in low doses and induce climate hormesis in woody plants).In addition,hormetic stimulation of woody species can occur as a result of direct exposure to low doses of climate stressors and through indirect effects of the stressors on other parts of ecosystems.Moreover,the interactions among climate stressors (synergism,antagonism,additivity) can alter the manifestation of hormesis in woody plants.Woody species can acclimate to a climate stressor and cease to be stimulated by the stress,and the optimal value and tolerance range to the stress may also change.
This review highlights the need for a targeted study of climate hormesis in woody species because hormetic stimulation of growth and productivity of dominant tree species can potentially affect the productivity of ecosystems,forests,forest succession,and geochemical cycles and mitigate the climatic effects of anthropogenic CO2emissions by increasing the absorption of atmospheric carbon.Long-term studies of climate hormesis for individual populations of tree species and at the ecosystem level in different climate zones should assess the role of climate hormesis in woody plants in the adaptation of ecosystems to climate change and the potential of using climate hormesis to manage the productivity of forest ecosystems and predict global and local consequences of climate change and mitigate the negative consequences of these changes.
 Journal of Forestry Research2023年1期
Journal of Forestry Research2023年1期
- Journal of Forestry Research的其它文章
- Epigenetic modification associated with climate regulates betulin biosynthesis in birch
- Growth trends and environmental drivers of major tree species of the northern hardwood forest of eastern North America
- Different xylogenesis responses to atmospheric water demand contribute to species coexistence in a mixed pine-oak forest
- Interannual dynamics of stemwood nonstructural carbohydrates in temperate forest trees surrounding drought
- Allocation patterns of nonstructural carbohydrates in response to CO 2 elevation and nitrogen deposition in Cunninghamia lanceolata saplings
- Different hydraulic strategies under drought stress between Fraxinus mandshurica and Larix gmelinii seedlings