
基于ISSR分子标记的紫云英种质资源遗传多样性及结构分析
2023-02-10 03:12史鹏飞刘春增李本银郑春风张成兰何春梅施海帆吕玉虎常单娜曹卫东
草地学报 2023年1期
张 梦,史鹏飞,刘春增,李本银,丁 丽,张 琳,郑春风,张成兰, 何春梅,施海帆,吕玉虎,常单娜,曹卫东*
(1.河南省农业科学院植物营养与资源环境研究所,河南 郑州 450002;2.信阳市农业科学院,河南 信阳 464000;3.河南省作物分子育种研究院,河南 郑州 450002;4.福建省农业科学院土壤肥料研究所,福建 福州 350013;5.南京农业大学草业学院,江苏 南京 210095;6.中国农业科学院农业资源与农业区划研究所,北京 100081)
紫云英(AstragalussinicusL.)是我国南方稻田重要的冬季绿肥,在培肥地力、降低化肥依赖、提升稻米产量和品质中发挥重要作用[1-2]。我国是紫云英的起源地,种质资源丰富,据推测紫云英起源中心在我国秦岭以南的中部山间河谷地带,并向东、向南扩展[3]。上世纪90年代以前,紫云英是我国农业生产主要有机肥源之一,种植分布广泛,形成了众多地方品种[4-5]。90年代以后,由于种植结构调整以及化学肥料的普及使用,紫云英种植应用范围急剧缩小,紫云英研究和选育工作停滞[2]。近年来,随着国家发展战略和社会大众对生态环境的重视,紫云英的产业化推广与应用获得了恢复性发展。但由于种质利用和研究中断多年,原有品种退化、损失、混杂严重,现有紫云英地方品种和种质资源遗传关系不明确,制约了对紫云英新品种的科学选育和管理认证[6]。
分子标记在作物遗传育种领域得到广泛应用,对于揭示供试材料遗传特征具有重要意义。陈坚等[7]利用简单重复序列(Simple sequence repeat,SSR)标记分析了9个紫云英品种遗传多样性,发现供试品种可聚为2个类群,且表现出一定的区域分布特征。与其他分子标记相比,简单序列重复区间扩增多态性分子(Inter-simple sequence repeat,ISSR)标记具有稳定、多态性高、简便及易操作等优点,被视为理想的遗传标记方法,已在多种动植物的亲缘关系[8]、种质鉴定[9]、遗传完整性[10]等研究方面得到应用。孙清信等针对紫云英优化了ISSR-PCR反应体系[11],张慧等[12]利用ISSR标记研究了 22个紫云英品种遗传多态性,发现供试品种平均多态率为59.2%,品种间的相似性系数界于0.63~0.95,22个紫云英品种可分为 4类。然而,由于长期缺乏有效管理和保护,加之研究中断造成部分品种和种质资源来源不明,育种资源遗传特征研究的不足制约了以紫云英为代表绿肥种业的进一步发展。
绿肥是基于绿肥作物通过固定光合产物和环境养分并在一定时期翻压还田形成的有机肥源,它的生产需要配合不同的生产制度和种植模式,在空间和时间“夹缝”中实现资源高效利用。因此,绿肥的产业化发展需要不断强化种质资源创新与利用、培育优质绿肥新品种,以适应生产方式、种植制度和应用模式的多样化需求,从根本上摆脱“依靠财政投入才能推广应用的传统有机肥料”之尴尬局面。因此,本文以绿肥作物紫云英为研究对象,利用国家绿肥种质资源库及各地不同时期的紫云英种质资源,通过分子标记分析供试资源的遗传多样性,明确不同紫云英种质资源的遗传关系及其遗传结构,评价育种材料在省份间交流和利用情况,以期为紫云英现有种质资源的充分利用、新品种选育以及现有品种的有效鉴别提供依据。
1 材料与方法
1.1 供试材料
参试紫云英种质资源共77份(表1),其中53份资源由中国农业科学农业资源与农业区划研究所(编号1~14,17和42~77)和福建省农业科学院土壤肥料研究所(编号15~16)提供;22份资源(编号18~39)为研究团队近5年在豫南地区收集获得;2份资源(编号40~41)为豫南地区主推紫云英品种。研究中根据来源地的不同将供试资源进行分类编号。

表1 77份紫云英种质资源
1.2 DNA提取
采用苗盘培养的方式,于苗期4片真叶完全展开时,对供试的77份资源进行采样。鉴于异花授粉植物群体内遗传多样性丰富,个体间可能存在较大差异,参考陈坚等[13]研究结果及供试资源数量,本研究采用多单株分别提取混合测定的取样策略:每份资源分别选择生长正常的紫云英幼苗25个单株,采集顶部真叶粉碎混匀后采用磁珠试剂盒(北京博友顺生物技术有限公司)分别提取紫云英叶片DNA,以1.5%琼脂糖凝胶电泳检测质量,并用Nanodrop分光光度计测定DNA浓度和质量。将检测合格的DNA样品稀释至40 μg·L-1,最后把同一资源的25份DNA稀释液等体积混合,置于—20℃下保存备用。
1.3 引物筛选及PCR扩增
参考孙清信等[11]和张慧等[12]的方法,以‘浙紫71-107’‘宁绿1号’‘闽紫3号’‘XYXH17-4’为模板,依据最优ISSR-PCR体系中的扩增条件,通过初步筛选及温度梯度复筛,从56条ISSR引物(北京六合华大基因科技有限公司)中筛选条带多态性高、重复性好、条带清晰的引物14条,对供试紫云英种质资源进行ISSR分析。其中,PCR扩增体系总体积为25 μL,DNA模板40 ng,10×Bufer 2.5 μL,primer 0.3 μmol·L-1,dNTP mixture 0.25 μmol·L-1,MgCl21.5 mmol·L-1,TaqDNA聚合酶1.25U(TaKaRa,宝生物工程(大连)有限公司);PCR扩增程序为94℃预变性5 min,然后进行35个循环:94℃变性45 s,56~52℃复性45 s,72℃延伸1.5 min;循环结束后72℃延伸10 min,4℃保存。PCR结束后,将PCR产物在2.0%的琼脂糖凝胶检测扩增产物(电压120 V,电泳50 min,电泳缓冲液浓度TAE),Marker为2 000 bp。电泳后凝胶成像仪(K8300北京科创锐新生物科技有限公司)进行成像拍照并保存DNA检测结果。
1.4 数据分析
PCR扩增产物电泳在凝胶的某个相同迁移率位置上有DNA条带记为1,无DNA条带记为0。使用Gel-pro analyzer软件估算出每个引物多态性条带,形成ISSR的表型数据矩阵,进行有关分析。
1.4.1个体间遗传特征分析 使用NTSYS软件计算相似系数(DICE系数),对77份紫云英地方种质资源进行UPGMA聚类分析。参考宋海斌等[14]和王琰琰等[15]方法,构建不同种质资源的指纹图谱:将14条引物依次编码为A~N,条带由小到大进行数字编号。根据扩增情况,每个紫云英种质资源形成了由字母和阿拉伯数字组成的带型信息码,如A1A5B2,表示该材料在引物A的第1和第5位点扩增出条带,在引物B第2位点扩增出条带。使用Excel中COUNTIF函数,逐次迭代比对信息码的条带信息,获得各引物可单独鉴别供试紫云英资源的数量和种类信息,并据此优化组合并缩减引物数量,获得核心引物(用最少的引物组合区分供试材料)。利用SPSS 22中神经网络模块的多层感知器进行特征选择,依据特征重要性逐次迭代核心引物位点信息,筛选能够区分供试资源的最小特征位点集合,最终构建出基于供试紫云英种质资源的指纹图谱编码。
1.4.2群体间遗传特征分析 采用GenAlEx 6.503加载宏[16]对不同省份(地区)的种质资源ISSR分子标记进行主坐标分析(PCoA),POPGENE 1.32软件[17]对不同来源地供试材料进行群体遗传相似性分析。由于本研究不同省份(地区)种质资源数量差异较大,因此基于主坐标分析和群体遗传相似性特征,将江苏-上海-安徽资源合并为苏沪皖资源库(SHW)、湖南-江西-广西-四川资源合并为湘赣桂川资源库(XGGC),对河南资源库(HE,24份资源)、福建资源库(FJ,13份资源)、浙江资源库(ZJ,11份资源)以及苏沪皖资源库(SHW,8份资源)、湘赣桂川资源库(XGGC,20份资源)进行分子遗传变异方差分析(AMOVA)。日本兵库种(AB-01)由于无法推测传播途径,未纳入本分析。
1.4.3省份材料遗传结构分析 不同于野外自然材料,供试绿肥种质资源在不同地区的数量确定,包括野生材料和人工杂交选择的育种材料。同时,由于材料来源较为复杂,野生材料不能明确是否为人工选育材料散落后的自然变异,部分育种材料亦不能考证亲本来源(与90年代后相关研究中断有关)。因此,鉴于供试材料涵盖了全国主要的紫云英生产省份以及主要栽培种,参考分子生态学方法利用Structure软件[18-19],对群体进行K=1~10的贝叶斯聚类,计算群体和个体Q矩阵,利用Structure Harvester[19]计算lnP(K)和ΔK,利用CLUMPP软件[20]对重复抽样数据进行分析,Distruct软件[21]对群体结构分析并进行数据可视化,以评价不同省份对紫云英种质资源的利用程度,推测我国主要紫云英栽培省份种质资源特征。标准地图由自然资源部地图技术审查中心标准地图服务网(http://bzdt.ch.mnr.gov.cn/)获得。
2 结果与分析
2.1 ISSR扩增结果及多态性
提取基因组总DNA的OD260/OD280在1.573~1.789之间,DNA浓度均大于40 μg·L-1,定量稀释混合后满足ISSR标记分析。筛选出14条扩增条带清晰、条带数较多、多态性良好的ISSR引物(图1)。在77个样品中共扩增出了150条清晰条带,137条为多态位点,平均多态性比率89.34%(表2)。各引物扩增的条带在4~16条之间,平均每条引物扩增的条带数为10.7条。观测等位基因数1.50~2.00,有效等位基因数1.13~1.70,基因多样性指数0.10~0.40,信息指数0.18~0.59,均以P809引物最高(表2)。引物分析显示,ISSR标记在77份供试紫云英种质资源中具有丰富的多态性,具备反映供试资源遗传特征的作用。

表2 ISSR引物信息及扩增结果
2.2 不同紫云英种质资源的个体遗传关系
77份紫云英种质资源的遗传相似性系数(Genetic similarity,GS)介于 0.567 7~0.868 6之间(图2)。其中,安徽‘当涂种’(AH-01)与‘弋江籽’(AH-02)之间遗传相似性系数最大(GS=0.868 6),说明两个资源亲缘关系最近;湖南‘澧县种’(HU-10)与河南‘DSQS17-4’(HE-14)之间相似系数最小(GS=0.567 7),说明两个资源亲缘关系最远。根据遗传相似性系数矩阵,对77份供试材料进行聚类分析,得到树状聚类图(图2)。在遗传相似系数为0.720 0时,77份资源可分为6个聚类:聚类Ⅰ包括日本‘兵库种’、广西‘萍宁3号’、四川‘川西种’以及全部供试江西资源和6份湖南资源;聚类Ⅱ包括全部供试安徽资源和17个河南资源;聚类Ⅲ包括12个福建资源、6个湖南资源、5个河南资源以及全部供试江苏、上海、浙江的资源;聚类Ⅳ为福建‘闽紫3号’;聚类Ⅴ为河南‘DSQS17-4’;聚类Ⅵ为河南‘DSHS17-6’,供试资源在聚类中的分布呈现出明显的区域聚集特征。

图2 基于ISSR的77份紫云英种质资源的聚类图
2.3 地区间种质资源的群体遗传分化
各地区紫云英种质群体间的遗传距离(Genetic distance,GD)存在差异(表3),地区间资源的平均遗传距离呈现出:国外>广西>四川>安徽>上海>江苏>河南>江西>福建>湖南。国外资源与湖南、江西、福建资源的遗传距离相对较小(GD<0.30),与安徽、四川资源的遗传距离相对较大(GD > 0.37)。各省份资源中,存在福建-浙江、福建-湖南、福建-河南、湖南-江西等4个遗传较为相似(GD< 0.05)的群体。主坐标分析进一步呈现出不同地区资源的相似性(图3)。总体上,福建紫云英资源位于轴坐标中心位置,河南、安徽、江苏、上海紫云英资源位于横轴负值区域,江西、湖南、四川以及国外资源位于横轴正值纵轴负值区域,浙江资源则位于横轴纵轴正值区域(图3,不同地区个别资源存在一定的离散),说明不同地区紫云英资源具有一定分化,而福建紫云英资源可能与其他省份存在遗传交流。由于不同省份(地区)种质资源数量差异较大,广西、四川、上海、安徽资源较少,因此基于主坐标分析和群体遗传相似性特征,将江苏-上海-安徽资源归为苏沪皖资源库(SHW)、湖南-江西-广西-四川资源归为湘赣桂川资源库(XGGC)。对河南资源库(HE,24份资源)、福建资源库(FJ,13份资源)、浙江资源库(ZJ,11份资源)以及上述两个资源库(SHW,8份资源;XGGC,20份资源)进行分子遗传变异方差分析(AMOVA),进一步验证区域资源的遗传分化情况。结果显示,群体分化系数为0.136 3,达到极显著水平(P< 0.001),其中种群间变异占比13.9%,种群内变异占比86.1%(表4),说明5个供试群体存在显著的遗传分化。
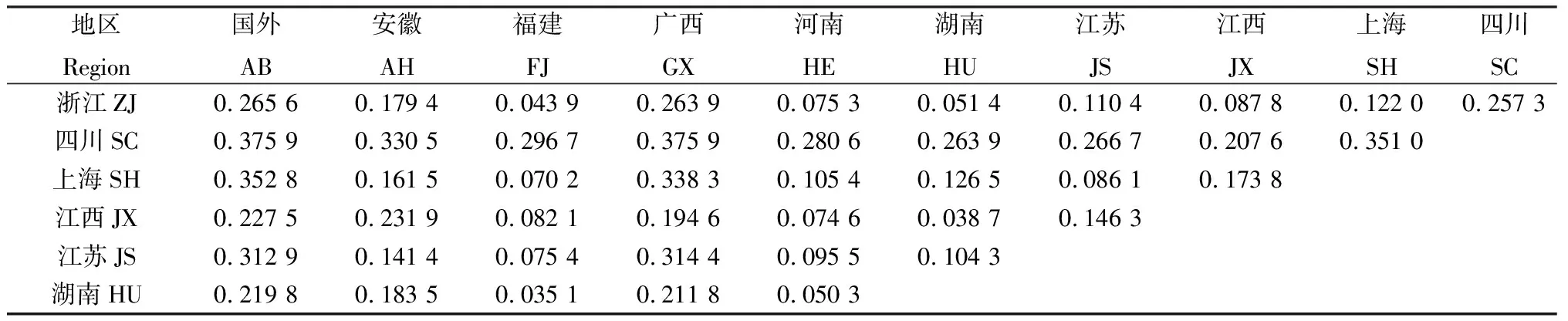
表3 不同地区紫云英种质资源群体的遗传距离

表4 紫云英种质资源群体的分子遗传变异方差分析
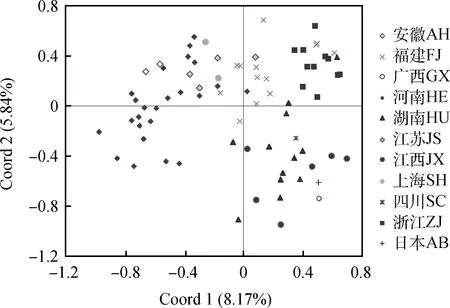
图3 不同地区紫云英种质资源的主坐标分析
2.4 不同地区种质资源的群体遗传结构
不同地区紫云英种质资源的群体遗传结构存在差异(图4)。当假设供试种质资源存在2个亚群体(K=2)时,10个地区中,河南、安徽、江苏、上海遗传结构相似,四川、广西、湖南、江西、浙江资源的遗传结构相似,福建则为亚群体混合型;当假设存在3个亚群体(K=3)时,河南、安徽遗传结构相似,江苏、上海相似,四川、广西、湖南、江西相似,浙江则自为一体,福建仍呈现亚群体混合型。基于数据对数概率变化率(ΔK)推测最佳亚群体数,当K=2时,ΔK=36.59,而K≥3时,ΔK<2.8,说明10个省份(群体)的供试种质资源最佳亚群体数应为2个。关联地理位置及推测的起源中心,供试紫云英呈现明显的传播特征,即以河南-安徽-江苏-上海-福建为一线(北方资源库),以四川-广西-湖南-江西-福建为一线(西部和南方资源库),紫云英群体遗传结构逐渐变化。需要说明的是,群体遗传结构分析与个体聚类结果存在差别(图2和图4),主要原因是两者分析方法和研究对象不同所致。

图4 不同省份紫云英资源的遗传结构
2.5 引物鉴别分析及指纹图谱构建
利用循环迭代的方法分析了14个引物对77份紫云英种质资源的鉴别能力,发现不同ISSR引物扩增条带信息鉴别供试资源的能力存在差异,表现为P809=P825>P886>P821>P810>P836>P868>P843>P811>P844>P846>P859>P857=P889,其中P809和P825均可单独鉴别67份资源,而P857和P889无法单独鉴别供试资源(如图5)。基于引物的鉴别能力,通过神经网络模型进行特征提取,筛选出P809和P886为核心引物,同时为尽可能降低指纹位数,通过迭代法筛选特征位点,构建出以P809第3,8,9,10,12,15扩增位点以及P886第1,3,5,9,10,11扩增位点为特征的指纹图谱(表5)。

图5 ISSR引物鉴别特征图谱
3 讨论
3.1 紫云英种质资源的遗传多样性与指纹图谱构建
本研究对11个地区收集的77份紫云英地方种质资源开展ISSR分子标记分析,14条引物共扩增出150个条带,137条为多态性条带,各引物的基因多样性和信息指数为0.10~0.40和0.18~0.59。陈坚等[7]对9个紫云英品种遗传多样性分析发现,供试资源遗传多样性指数为0.13~0.38,多态信息指数为 0.22~0.56,与本研究结果相似。此外,本研究供试种质资源之间遗传相似性系数为0.567 7~0.868 6,相似性最高的安徽‘当涂种’(AH-01)与‘弋江籽’(AH-02)之间亦检测出23个差异位点,占总多态性位点的16.8%,说明77份紫云英地方资源不存在异名同种现象,具备遗传特异性。目前,不同紫云英的分子标记鉴别方法并未形成,如何高效便捷鉴定不同紫云英资源,成为绿肥行业发展面临的重要问题。本研究中发现不同ISSR引物对77份紫云英资源的分子标记特征存在差异,虽然14个引物的多态性条带比率为50%~100%,但引物对不同资源的区分能力却存在差别,表现为P809=P825>P886>P821>P810>P836>P868>P843>P811>P844>P846>P859>P857=P889,其中P809和P825单独可区分67份资源,而P857和P889无法单独鉴别供试资源。因此,基于资源的遗传特异性以及ISSR不同引物对资源的区分鉴别能力,构建了以P809和P886条带为核心的紫云英ISSR标记指纹图谱。宋海斌等[11]和王琰琰等[15]利用SSR标记分别对102份甜瓜(CucumismeloL.)品种和220份雪茄烟(NicotianatabacumL.)种质资源进行了指纹图谱分析,提取核心引物18对和14对用作指纹分析,与本研究方法相似,但引物数量较多,原因可能是其供试资源数量高于本研究、使用的分子标记亦不同。指纹图谱应用具有一定局限性,如P809和P886分别可扩增16和14个条带,理论上最高可鉴别230=1 073 741 824份资源,77份资源鉴别仅需要7个有效条带(27=128>77),而实际情况却是14个供试引物均无法单独鉴别77份资源,说明引物的每个条带并非皆为区分种质资源的有效条带。因此,若紫云英种质资源数量增大,核心引物或者特征位点数量也将增多。目前,我国紫云英种质资源主要保存在中国农业科学院、福建省农业科学院、江西省农业科学院和湖南省农业科学院,有效种质资源的具体数量并未明确,对紫云英指纹图谱的开发和应用仍有待深入研究。
3.2 不同地区紫云英种质资源的遗传关系、利用情况及演化推断
本研究中,77份供试资源可分为6个聚类,聚类组成分布呈现出明显的区域聚集特征,同一地区种质资源倾向于聚集一个聚类(图2和图3),说明紫云英在传播过程中表现出明显的分化和地理分割性,这种特征与刘硕等[22]对中亚及我国普通杏(PrunusarmeniacaL.)和西伯利亚杏(PrunussibiricaL.)野生居群,张旭等[23]对内蒙古、宁夏、甘肃16个不同居群红砂(ReaumuriasoogoricaM.)的研究结果类似,却与邓亚博等[24]对河南省内狗牙根(CynodondactylonP.)以及钱茱希等[25]对洞庭湖地区南荻(TriarrhenalutarioripariaL.)种质资源研究结论(地理距离与种群的遗传变异分布无显著相关性)不同,推测研究的空间尺度可能影响上述结论的产生。另一方面,自然地理条件、气候因子、人为因素都是影响物种遗传分化的重要因素[26-27]。前人专著《中国绿肥》记录了秦岭、大别山各山区湿润地带有野生种分布的现象[4];《中国紫云英》则根据我国紫云英农家品种大多数冬性都较强、有一定的耐寒性及适应于较高的海拔等特性推测,它的起源地可能在秦岭以南的中国中部山间河谷地带,然后向南向东扩展[3]。本研究基于ISSR分子标记进行种群遗传结构分析发现,不同地区紫云英种质资源的遗传结构呈现明显的地域特征,即以河南-安徽-江苏-上海-福建为一线,供试群体遗传结构类似,可称为北方资源;以四川-广西-湖南-江西-福建为一线,资源的遗传结构类似,可称为西南方资源。若参照前人所推测的起源中心,即秦岭以南的中国中部山间河谷地带为紫云英的起源中心(图5),则上述群体遗传结构特征印证了前人关于紫云英起源与扩展的推断。对于福建为何呈现出亚群体混合型特征,可能的原因有两个:一是地理位置和地形原因造成福建成为紫云英传播链末端;二是福建紫云英相关研究(尤其是杂交育种)开展较早、较为系统收集了我国其他省份的种质资源,开展了广泛的杂交育种工作及其相关研究[3,7-9]。然而,本研究的结论建立在14条ISSR引物对77份(国内76份)不同地区紫云英的150个扩增位点信息的基础上,供试资源具有一定代表性,但缺少野生资源及相关育种材料亲本的重要信息,人为因素(如耕作制度变化、骨干亲本选择)或自然气候(如历史气温、降水变化等)如何影响紫云英传播和分化并未明确,不同紫云英种质资源形成的分子生态学原理以及各省对于紫云英种质资源在育种中的应用仍有待深入研究。针对绿肥产业发展,未来应加强起源地科考、人工诱变研究,不断收集和创制种质资源,配合加代扩繁、快速育种等技术,大量培育适应不同生产需要的绿肥新品种,形成推动绿肥产业发展的“源动力”,更好服务种植业的绿色低碳发展。
4 结论
77份供试紫云英种质资源遗传相似性系数为0.567 7~0.868 6,既具有丰富的遗传多样性,又表现出明显的分化和地理分割特征。根据引物条带多态性及其鉴别能力,构建了以P809第3,8,9,10,12,15扩增位点和P886第1,3,5,9,10,11扩增位点为特征的指纹图谱。以资源所在地区为居群进行群体遗传变异和结构分析发现,群体分化系数为0.139 3,达到极显著水平(P< 0.001),其中河南-安徽-江苏-上海资源可划为北方资源群,四川-广西-湖南-江西资源为西南方资源群,遗传结构特征在福建发生融合。
猜你喜欢
武术研究(2021年2期)2021-03-29
上海农业科技(2020年2期)2020-04-02
贵州林业科技(2019年2期)2019-08-26
山西地震(2019年1期)2019-03-20
西安交通大学学报(2019年3期)2019-03-08
中学时代(2018年6期)2018-11-20
海峡姐妹(2018年7期)2018-07-27
电子制作(2017年10期)2017-04-18
中国教育技术装备(2016年11期)2016-12-01
作文与考试·小学高年级版(2016年14期)2016-09-10
