
基于COⅠ基因序列的泥鳅、大鳞副泥鳅选育群体杂交子代遗传多样性分析
2023-02-08 10:09:06李福贵程林慧何坤张士林刘铮铮孙艺文阮记明吴华东
水产科技情报 2023年1期
李福贵 程林慧 何坤 张士林 刘铮铮 孙艺文 阮记明 吴华东
(江西农业大学动物科学技术学院,江西南昌 330045)
泥鳅(Misgurnusanguillicaudatus)和大鳞副泥鳅(Paramisgurnusdabryanus)分别隶属于鳅科(Cobitidae)、花鳅亚科(Cobitinae)的泥鳅属(Misgurnus)和副泥鳅属(Paramisgurnus),在我国及东南亚水域广泛分布,是名特优经济鱼类品种[1-2]。然而,由于目前缺少国家审定的鳅科鱼类新品种,再加上近亲繁育、过度捕捞、养殖水域水体污染等因素,泥鳅的种质资源正急剧衰退,因此,亟需加强泥鳅种质资源保护和种质创新利用。
遗传多样性是物种资源状况评价的重要指标,也是物种适应环境变化和进化的基础。通过遗传多样性分析,可以评价物种资源现状和种质创新应用潜力[3-4]。线粒体细胞色素氧化酶亚基Ⅰ基因COⅠ具有母系遗传、进化速率适中等特点,其序列常被用作物种鉴定的分子条形码,也被广泛应用于鱼类群体遗传学和系统学研究[5-12]。笔者团队前期开展了泥鳅、大鳞副泥鳅群体选育,获得了具有较明显生长优势的杂交子代,为新品种选育提供了优良群体[2]。本研究基于线粒体COⅠ基因序列,分析了经两代群体选育的泥鳅、大鳞副泥鳅自繁及其杂交后代的遗传多样性和遗传结构,以期对两个选育群体开展科学评估,并为后续泥鳅新种质资源创新利用提供基础资料和参考依据。
1 材料和方法
1.1 样品采集和基因组DNA提取
本研究中亲本培育、苗种繁育等试验均在江西省吉安市新干县赣源生态泥鳅养殖场(北纬27°47′12.96″,东经115°27′35.67″)内完成,遗传多样性分析试验在江西农业大学鄱阳湖水生动物遗传育种与生物技术实验室完成。2018年5月,选择经两代群体选育的二倍体(见图1)泥鳅(M)与大鳞副泥鳅(P)作为亲本,通过M(♀)×M(♂)与P(♀)×P(♂)群体自繁获得M、P两个自繁后代,通过M(♀)×P(♂)与P(♀)×M(♂)获得正交(MP)和反交(PM)两个杂交后代。

图1 亲本DNA含量测定
从4个后代群体中各随机抽取30尾泥鳅(共120尾),剪取鱼鳍,置于无水乙醇中,-20 ℃冷冻保存备用。参考标准酚-氯仿抽提法[13]提取并纯化基因组DNA,共获得120个DNA样品。用1.4%的琼脂糖凝胶电泳检测DNA质量,用微量核酸浓度测定仪测定其浓度,于-20 ℃冷冻保存。
1.2 PCR扩增及测序
参照张望等[14]报道的方法设计4个泥鳅群体COⅠ基因序列扩增引物,其中上游引物序列为:5’-CACGACGTTGTAAAACGACCAACYAATCAYAAAGATATYGGCAC-3’,下游引物序列为:5’-GGATAACAATTTCACACAGGACTTCYGGGTGRCCRAARAATCA-3’。引物由生工生物工程(上海)股份有限公司合成,于-20 ℃保存备用。
聚合酶链反应(PCR)采用10 μL体系,其中含染料的TIANGEN 2×TaqPCR Master Mix 5 μL,上、下游引物各0.4 μL,模板DNA 0.4 μL,DEPC水(无DNA/RNA酶)3.8 μL。PCR反应在BIO-RAD T100TMThermal Cycler型PCR检测仪上进行,反应体系为:94 ℃预变性3 min;94 ℃变性40 s,52 ℃退火1 min,72 ℃延伸1 min,重复35个循环;72 ℃再延伸10 min;4 ℃保存。PCR产物经1.4%琼脂糖凝胶电泳检测后,从每个群体中各随机挑取15个阳性扩增产物(共60个)送至生工生物工程(上海)股份有限公司进行双向测序。
1.3 数据统计与分析
线粒体COⅠ基因PCR产物测序完成后,用Clustal X软件进行序列比对分析,用Codon W软件分析密码子的使用偏好性,用DNASP 5计算遗传参数、MEGA 5.0计算群体遗传距离,采用邻接法(neighbor-joining,NJ)并基于Kimura双参法构建邻接关系进化树,利用Network 5010软件构建单倍型网络图。
2 结果
2.1 COⅠ基因序列的碱基组成和密码子偏好性
经比对分析,得到4个泥鳅种群COⅠ基因片段同源序列,其全长为682 bp,共检测到135个多态位点(占比19.79%),其中有128个简约信息位点,7个单碱基变异位点,变异均为转换或颠换,且转换多于颠换,未发现碱基的插入与缺失(见图2)。

图2 4个泥鳅种群COⅠ基因片段的变异位点比较
4个泥鳅种群的COⅠ基因片段碱基组成相似,且种间差异小。4个泥鳅种群的COⅠ基因片段碱基A、T、C、G含量平均值分别为22.85%、31.9%、26.6%、18.7%,A+T的含量(54.0%~56.2%)均高于C+G的含量(43.8%~46.4%),呈现出典型的碱基组成偏向性(见表1)。
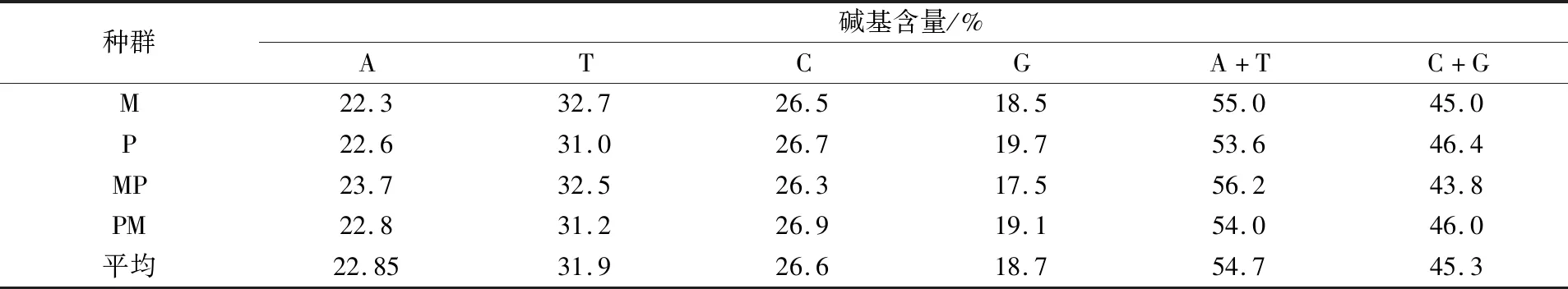
表1 4个泥鳅种群线粒体COⅠ基因序列碱基组成的比较分析
4个泥鳅种群COⅠ基因编码氨基酸时的密码子使用偏好性分析结果显示,M、P、MP、PM明显偏好使用的密码子分别有31、32、36、31个,其中Leu-CUU是M、MP、PM最主要的密码子,TER-UGA和Arg-CGA是P的主要密码子。M、P、MP、PM这4个泥鳅种群分别有7、12、6、6个密码子使用率为0,表明这些密码子在编码氨基酸时没有被使用(见表2)。

表2 4个泥鳅种群COⅠ基因密码子的使用偏好性

表2 (续)
除去使用率始终为1和非编码氨基酸的终止密码子,基于剩余密码子对基因的密码子使用偏好性进行聚类分析,结果见图3。由图3可见,MP与M首先聚为一支,再与PM、P聚类。这一结果表明MP的密码子使用偏好性与母本M更为相似,也一定程度上表明泥鳅和大鳞副泥鳅杂交子代偏母系遗传。

图3 基于COⅠ基因密码子偏好性的聚类图
2.2 遗传多样性和单倍型组成
4种泥鳅COⅠ基因的遗传多样性参数见表3。M、P、MP、PM 4种泥鳅的单倍型多态性(h)均为1.000±0.024,整体合计单倍型多样性(h)为0.997±0.004;M、P、MP、PM 4种泥鳅的核苷酸多态性(π)分别为0.020 66±0.006 48、0.025 80±0.002 31、0.029 61±0.005 62和0.022 30±0.001 94,整体合计核苷酸多态性(π)为0.092 82±0.001 77。综上,4种泥鳅均具有高单倍型多样性和高核苷酸多态性,多态性由高到低依次为MP>P>PM>M。

表3 4种泥鳅COⅠ基因的遗传多样性参数
4个泥鳅种群的单倍型分布情况见表4。60个泥鳅样本共检测到56个单倍型,包括3个种类共享单倍型和53个独有单倍型。其中,P群体中15个个体共检查到15个独有单倍型(Hap1~Hap15);M群体中13个个体为独有单倍型(Hap16~Hap25,Hap27~Hap29),2个个体共享1种单倍型(Hap26);MP群体中13个个体为独有单倍型(Hap30~Hap32,Hap34~Hap43),2个个体共享1种单倍型(Hap33);PM群体中12个个体为独有单倍型(Hap44~Hap46,Hap48~Hap56),3个个体共享1种单倍型(Hap47)(见表4)。利用Network 10.20.0软件对4个泥鳅种群间的单倍型进化关系进行分析,结果显示,MP的单倍型与M聚集在一个主支,PM的单倍型与P聚集在一个主支(见图4)。

表4 4个泥鳅种群的单倍型分布
2.3 遗传距离
M、P、MP、PM 4个泥鳅种群的种内遗传距离分别为0~0.020、0~0.024、0~0.027和0~0.023,种间遗传距离在0.024~0.180,其中MP与M遗传距离最近,为0.046;PM与P遗传距离最近,为0.024(见表5)。

表5 基于COⅠ基因不同单倍型的Kumara 2-parameter遗传距离
采用邻接法构建分子系统进化树(见图5),结果显示,4个泥鳅种群同种不同个体间大部分以较高的置信值分别聚类,相聚之后再与外群聚类;有3个MP个体与M聚类,2个M个体与MP聚类,7个PM个体与P聚类,6个P个体与PM聚类,这与单倍型中介网络图的结果相符合。

3 讨论
3.1 4个泥鳅种群的COⅠ基因序列特征
本研究发现,在泥鳅、大鳞副泥鳅选育群体自繁和杂交后代群体的线粒体COⅠ基因序列中,A+T含量均高于C+G含量,呈现出明显的AT碱基偏好,此结果与硬骨鱼类COⅠ基因的研究结果[15-19]一致。
在4种泥鳅COⅠ基因的682 bp同源序列中共检测到135个变异位点,占比为19.79%,变异位点中有128个简约信息位点。施文瑞等[9]在大鳞副泥鳅4个群体104个样本642 bp的COⅠ基因序列中,共检测到44个多态性位点,仅占总位点数的6.85%,简约信息位点只有35个,这可能与分析区域或物种间的差异有关[3,9]。本研究发现,在4个泥鳅种群所检测的变异位点中,转换比例远大于颠换,尽管转换不易引起密码子编码的氨基酸变化,但仍呈现了丰富的变异特征。
3.2 4个泥鳅种群的遗传多样性
线粒体COⅠ基因因具有较高的遗传多态性而被广泛用于大鳞副泥鳅[9]、银鱼(Hemisalanxprognathus)[10]、带鱼(Trichiuruslepturus)[11]、哈氏仿对虾(Parapenaeopsishardwickii)和细巧仿对虾(Parapenaeopsistenella)[12]等水产动物遗传多样性及遗传结构分析。根据Grant等[20]提出的遗传多样性标准,单倍型多样性与核苷酸多样性的临界值分别为0.5和0.005,数值越高表明遗传多样性越高。本研究中,M、P、MP、PM 4种泥鳅的单倍型多态性(h)均为1.000±0.024,核苷酸多态性(π)分别为0.020 66±0.006 48、0.025 80±0.002 31、0.029 61±0.005 62和0.022 30±0.001 94,均显著高于临界值,全部呈现出高单倍型和核苷酸遗传多样性,遗传多样性由高至低依次为MP>P>PM>M。物种内遗传多样性或变异丰富度是物种对环境适应性和进化潜力的象征[21],因此可以预测,本研究中的4个泥鳅种群具有较强的环境适应力和遗传进化潜力。
3.3 4个泥鳅种群的遗传进化
物种密码子的使用具有保守性的特点,相近物种的密码子使用模式相似[22]。本研究结果表明,MP与M的密码子使用偏好性相似,P与PM的密码子使用偏好性更相似,表明泥鳅和大鳞副泥鳅杂交子代COⅠ基因序列偏母系遗传。单倍型多样性进化分析和COⅠ基因系统进化分析结果也共同验证了这一结果。群体间亲缘关系与其遗传距离和遗传相似系数有直接关系[23-24]。研究表明,不同种鱼类的线粒体基因组DNA序列存在10%左右的遗传变异[23],且基于COⅠ基因序列的种内遗传距离大多在0.02以下,而种间遗传距离在0.2以上[24]。本研究中,4个泥鳅种群的种内最大遗传距离为0.020~0.023,种间遗传距离为0.024~0.180,表明存在一定的种内差异,但种间差异较小。这是否与泥鳅、大鳞副泥鳅两个物种间存在基因嵌套现象有关[25],还有待进一步研究验证。
3.4 泥鳅种质创新建议
笔者团队在前期研究中发现,MP的形态特征与泥鳅(M)母本相似并具有明显的超母本生长优势[2]、性腺发育障碍等特点。因此,团队将针对上述两个方面继续开展泥鳅新品种的选育研究,具体工作为:(1)继续分别开展泥鳅、大鳞副泥鳅的群体选育至F4~F5代,最终通过两个物种间经济杂交获得杂种优势更加突出的MP新品种;(2)以泥鳅、大鳞副泥鳅选育群体间正交获得MP并辅以染色体组操作,以期获得异源四倍体泥鳅新品系(种);(3)若异源四倍体泥鳅有正常和稳定的经济性状且为可孕育群体,可与二倍体泥鳅或大鳞副泥鳅进行轮回杂交,获得异源三倍体泥鳅新品系(种)。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
作文周刊·小学一年级版(2022年16期)2022-05-07 11:28:30
生物学通报(2020年11期)2020-10-22 01:20:20
小学生导刊(2018年22期)2018-08-21 01:47:50
中成药(2018年7期)2018-08-04 06:04:10
琴童(2018年2期)2018-03-06 16:45:32
金色少年(奇趣科普)(2017年8期)2017-09-07 06:38:51
西南农业学报(2016年6期)2016-04-16 05:12:47
法医学杂志(2015年4期)2016-01-06 12:36:36
生命科学研究(2014年1期)2014-04-29 00:44:03
