
藏北高寒草甸土壤线虫群落结构和多样性对增氮的响应
2023-02-07 02:22:36沙玉宝干珠扎布胡国铮王学霞严俊何世丞高清竹
草业学报 2023年1期
沙玉宝,干珠扎布*,胡国铮,王学霞,严俊,何世丞,高清竹
(1.中国农业科学院农业环境与可持续发展研究所,北京 100081;2.国家农业环境那曲观测实验站,西藏 那曲 852000;3.北京市农林科学院植物营养与资源研究所,北京 100097;4.那曲市草原站,西藏 那曲 852000)
近百年来工业、农业兴起产生了大量的氮氧化合物,加剧了大气氮沉降,从而带来了一系列的生态环境问题[1],过量的氮素累积会对草地生态系统造成负面影响。研究表明氮沉降会导致草地土壤酸化,改变植被类型和植物群落的组成,从而间接地影响土壤动物群落的组成及其多样性[2]。土壤线虫是土壤食物链和食物网的重要组成部分,生活环境相对稳定,在土壤生态系统能量流动和物质循环中发挥着重要作用[3]。
氮沉降可以通过改变植物群落组成和结构进而影响土壤线虫群落,梁思维[4]在半干旱草地的研究表明,植物丰富度与土壤线虫的多度、丰富度存在显著正相关,氮添加降低了草地土壤线虫的多样性和密度。此外,氮沉降还可以通过提高可利用氮含量从而提高草地土壤线虫多样性[5]。也有研究表明,氮沉降改变了土壤中原有氮素的利用路径,使得土壤线虫及其相关微生物的关系发生了变化,从而改变了土壤线虫群落结构和功能[6-7]。增氮对不同c-p(colonizer-persister)型的土壤线虫影响不同,例如c-p 1类型线虫主要是由食细菌类和食真菌类线虫组成,土壤氮含量的增加可以促进土壤中细菌和真菌等微生物数量增加,从而间接提高草地土壤线虫群落中食细菌线虫和食真菌线虫数量[8-9]。此外,氮沉降可以改变植物群落组成和结构,影响土壤微生物群落和食物链,进而改变土壤动物群落组成和营养级结构[10-11]。
作为“世界屋脊”的青藏高原是亚欧大陆最大的地貌单元,草地资源十分丰富,其中藏北高原作为青藏高原重要的组成部分,平均海拔4500 m,分布着我国典型的高寒草地生态系统。氮沉降对藏北高寒草地土壤线虫群落组成和结构具有重要影响[12]。研究表明,氮沉降通过影响草地生产力[13]和物种的丰富度[14]、植物地下生物量以及土壤中的动物群落总量[15],改变高寒草甸土壤线虫数量,导致线虫物种多样性降低、优势度减少[16]。该地区已有的研究中,关于氮沉降对植物生产力和土壤养分的研究比较多,有学者研究发现增温通过改变植物群落组成和土壤微生物数量显著降低了高寒草甸土壤线虫密度[17],也有研究发现放牧改变了土壤线虫通道指数并提高了土壤有机质转化效率[18],但是关于藏北地区土壤线虫群落结构和多样性对不同水平氮沉降的响应机制仍不明确。
本研究通过模拟不同梯度氮沉降,研究藏北高寒草甸土壤线虫群落结构和多样性对氮沉降的响应,旨在明确影响土壤线虫群落变化的关键因素,为青藏高原高寒草甸生态系统适应性管理提供数据基础。
1 材料与方法
1.1 试验区概况
研究区位于西藏自治区那曲市色尼区国家农业环境那曲观测实验站(31°44′N,92°02′E)。该区平均海拔超过4500 m,属于高原亚寒带半干旱季风性气候,雨热同季,年均气温为0.3℃,生长季平均气温为8.0℃;年均降水量为462 mm,生长季降水量为403 mm,主要集中在5-9月;年日照时数为2790 h。试验地位于地势平坦、群落结构均匀的高寒草甸,主要以莎草科高山嵩草(Kobresia pygmaea)为建群种,其他的莎草科植物还有线叶嵩草(Kobresia capillifolia)和矮生嵩草(Kobresia humilis);伴生种包括禾本科的紫花针茅(Stipa purpurea)、早熟禾(Poa pratensis),豆科的藏豆(Stracheya tibetica)、紫花棘豆(Oxytropis subfalcata)以及杂类草钉柱委陵菜(Potentilla saundersiana)、二裂委陵菜(Potentilla bifurca)和肉果草(Lancea tibetica)等,平均植被覆盖率大于80%,土壤类型为高寒草甸土,土壤质地为砂壤土,5 cm深处的土壤容重为1.01 g·cm-3,土壤pH为7.05,0~15 cm深 度 土 壤 中 的 有 机 碳、总 碳、总 氮 和 总 磷 含 量 分 别 为41.39 g·kg-1,49.84 g·kg-1,6.78 g·kg-1和1.43 g·kg-1[19]。
1.2 试验设计
增氮试验开始于2014年,处理包括:Nck(0 kg N·hm-2·a-1)、N7(7 kg N·hm-2·a-1)、N20(20 kg N·hm-2·a-1)、N40(40 kg N·hm-2·a-1),每个处理4个重复,共16个小区,大小为3 m×3 m,相邻小区之间有间隔2 m的缓冲带。本研究所用的数据为2016年8月30日采集。氮素添加剂量参照中国氮沉降分布的格局,西藏地区的干湿沉降率为7 kg N·hm-2·a-1[20],所以本试验模拟浓度约为该地区自然沉降量的1、3和6倍,最高梯度也是本地区2050年氮沉降的估计浓度40 kg N·hm-2·a-1[19],该施氮浓度下不会对草地生态系统造成负面影响[21]。用NH4NO3模拟氮沉降,于每年5-8月的第一天给所有氮处理小区施肥,共计4次,Nck、N7、N20、N404个处理下每个小区每次施加硝酸铵的量为:0、4.6、13、26 g,即每个小区每年施加硝酸铵的量为:0、18.4、52、104 g。每次将硝酸铵溶于5 L水后施于样地,之后再浇5 L的水将叶片表面的硝酸铵冲洗掉,对照组添加等量的水以避免外加的水造成影响。各处理下每个小区全年用水40 L,约等于当地年降水量的1%。
1.3 测定指标与方法
1.3.1 土壤氮含量与地下生物量测定 2016年8月中旬,用直径5 cm的土钻取15 cm深土壤,每个小区取两钻,并混匀为一份样品,用于线虫的统计、土壤氮含量和植物地下生物量的测定。采用KCl浸提-靛酚蓝比色法对土壤氨态氮和硝态氮含量进行测定:取新鲜土壤10 g放入锥形瓶,加入1 mol·L-1氯化钾溶液50 mL,放在振荡器上震荡1 h,静置10 min后过滤,浸提液中的氨态氮和硝态氮用连续流动分析仪(Auto Analyzer 3,Bran Luebbe,德国)测定;挑出的植物根系用清水清洗,在烘箱中105℃杀青30 min,以70℃烘干至恒重,测定干重。
1.3.2 土壤线虫的分离与鉴定 取100 mL土壤,室温条件下,用改进的贝尔曼浅盘分离法分别经24、36、48 h后收集浅盘水,过筛、冲洗、保存[17],当分离出的线虫数量小于150条时,鉴定所有的线虫种类,当线虫数量多于150条时,只鉴定150条,然后统计所鉴定出的线虫数量。参照《植物线虫分类学》[22]和《中国土壤动物检索图鉴》[23],根据线虫的头部形态学特征和取食生境将土壤线虫划分到属,个体数占总捕获量10%以上的标记为优势类群:+++,个体数占总捕获量1%~10%的标记为常见类群:++,个体数占总捕获量不足1%的标记为稀有类群:+。
根据划分原则[23],将土壤线虫划分为4个营养类群:食细菌线虫(bacterivores,Ba)、植物寄生类线虫(plantpara-sites,Pp)、食 真菌线虫(fungivores,Fu)、杂 食-捕 食类线虫(predators-omnivores,Om)。同时参考Bongers[24]的研究,根据线虫不同的生活史对策,将其划分为r-对策者向k-对策者过渡的5个c-p类群:c-p 1到c-p 5;其中较为典型的机会主义c-p 1类线虫世代时间短,产卵量多,代谢较快,对不利环境抵抗性强,在食物充足条件下种群增长迅速;而c-p 5类线虫世代时间长,产卵量少,对生存环境中的压力变化极为敏感。
1.3.3 地上生物量测定 禾本科和莎草科在本研究中统称为禾莎草,本研究中豆科和杂草将统称为杂类草。采用无损测量法测定植物地上生物量[25]。无损测量法是指用样地附近植物高度、盖度和生物量建立回归方程,用该方程估算小区地上植物生物量。在试验样地附近随机选择20个0.5 m×0.5 m的校正样方,测定校正样方内各物种的高度和盖度。随后将校正样方地上植物齐地面剪下,分功能群装入信封中,带回实验室,在烘箱中105℃杀青30 min,以70℃烘干至恒重,测定干重。随后计算校正样方内禾莎草和杂类草的平均高度和总盖度,用平均高度H、总盖度C和地上生物量B建立回归方程,根据试验小区内每个功能群植物的平均高度和总盖度,用该回归方程计算试验样地内不同功能群植物的生物量。回归方程如下:
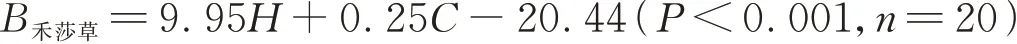

1.4 计算公式
参考Shannon-Wiener指数(H′)和Pielou均匀度指数(J)研究土壤线虫群落的多样性,公式如下:

式中:Pi为土壤线虫群落第i类群个体数占总个体数的比例;S为土壤线虫群落中所有线虫类群数。
采用自由线虫成熟度指数(maturity index,MI)、植物寄生线虫指数(plant parasite index,PPI)、线虫通道指数(nematode channel ratio,NCR)和瓦斯乐斯卡指数(wasilewska index,WI)表示高寒草甸土壤线虫群落功能结构特征。通常MI可以反映土壤生态系统稳定性及其受干扰程度,一般越稳定的土壤环境中成熟度指数越高;PPI与作物产量和土壤肥力密切相关,可以看出土壤线虫对植物的危害程度;NCR可以判断土壤有机质分解的途径,NCR若大于0.5,代表土壤中有机质分解以细菌为主,反之则以真菌为主;WI代表土壤中食细菌真菌线虫数量与植物寄生线虫数量的比值,WI小于1表明矿化途径主要由植物寄生线虫参与,反之表明矿化途径主要由食细菌真菌线虫参与[26]。公式如下:

式中:v(i)代表生态演替中对属于k-选择和r-选择的属分别赋予的c-p值,f(i)代表自由生活的线虫属在线虫种群中所占的比例,f′(i)代表植物寄生类线虫属在线虫种群中所占比例,NBF为食细菌线虫数量,NFu为食真菌线虫数量,NPp为植物寄生线虫数量。
1.5 数据分析
采用单因素方差(One-way ANOVA)分析增氮对土壤中线虫数量、植物生产力和土壤养分含量的影响;本研究数据利用IBM SPSS Statistics 22.0进行相关性分析,利用Microsoft Excel 2016和Origin 2021软件作图。
2 结果与分析
2.1 增氮对土壤线虫密度与群落组成的影响
在所有处理中,N7处理土壤线虫密度最小,为245条,N20处理下土壤线虫密度最大,为328条(图1)。较对照处理,N40和N20均显著增加了土壤线虫密度(P<0.05),分别增加了14.4%和25.4%。本试验中,16个小区共计分离得到土壤线虫4535条,53个属。其中,对照4个小区的线虫1046条,45属,增氮样地线虫3489条,38属。在Nck和N7样 地 中,拟 丽 突 属(Acrobeloides)为优势属,分别占个体总数的10.7%和10.8%。色矛属(Chromadorita)为N7样地的特有属,盘旋属(Rotylenchus)为N20样地的特有属,棱咽属(Prismatolaimus)、丝尾垫刃属(Filenchus)和似绕线属(Anaplectus)等在所有处理样地中均为常见属(表1)。
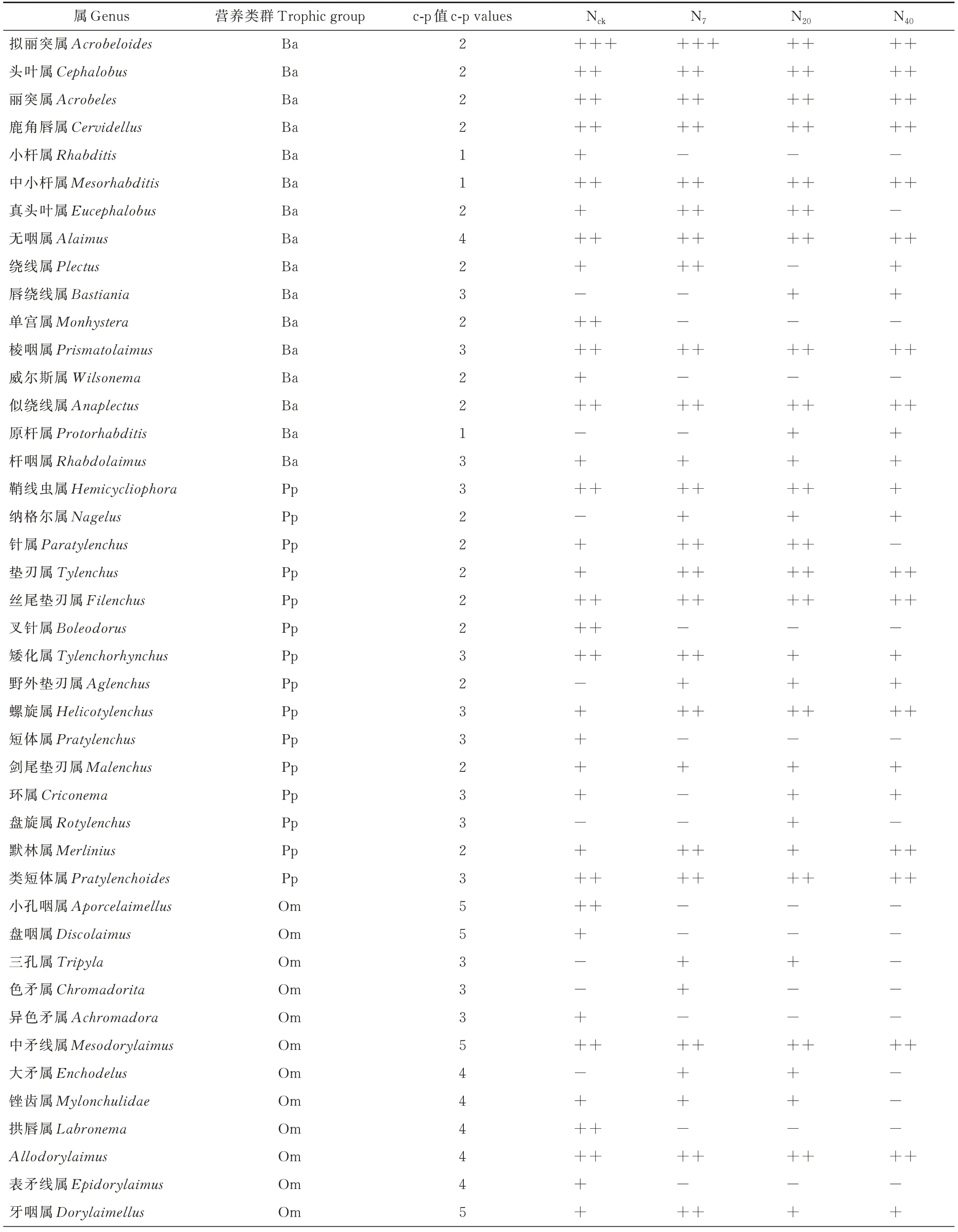
表1 增氮处理下高寒草甸土壤线虫群落组成、营养类群和c-p值Table 1 Compositions of nematode community and trophic groups and c-p values under nitrogen increase treatment in alpine meadow

图1 增氮处理下高寒草甸土壤线虫的密度Fig.1 The density of soil nematodes in alpine meadow under nitrogen increase treatment

续表Continued Table
食细菌线虫在所有样地中均为优势类,占个体总数的33.1%~42.8%;其次是植物寄生类线虫,占个体总数的25.2%~30.9%。在Nck和N7处理中,各类线虫所占比例表现为食细菌线虫>植物寄生类线虫>食真菌线虫>杂食-捕食类线虫;在N40、N20处理中,各类线虫所占比例表现为食细菌线虫>植物寄生类线虫>杂食-捕食类线虫>食真菌线虫,N40、N20处理中食真菌线虫所占比例显著低于Nck中食真菌线虫所占比例(P<0.05)(图2a)。对c-p分类而言,N20和N40中c-p 2类群相对丰度较Nck分别增加了57.1%和43.5%(P<0.05)。在所有处理样地中,各类别线虫占总数的比例均为c-p 2>c-p 4>c-p 3>c-p 1>c-p 5,其中c-p 1、c-p 3和c-p 5占总体比例在各个处理中均无变化(图2b)。
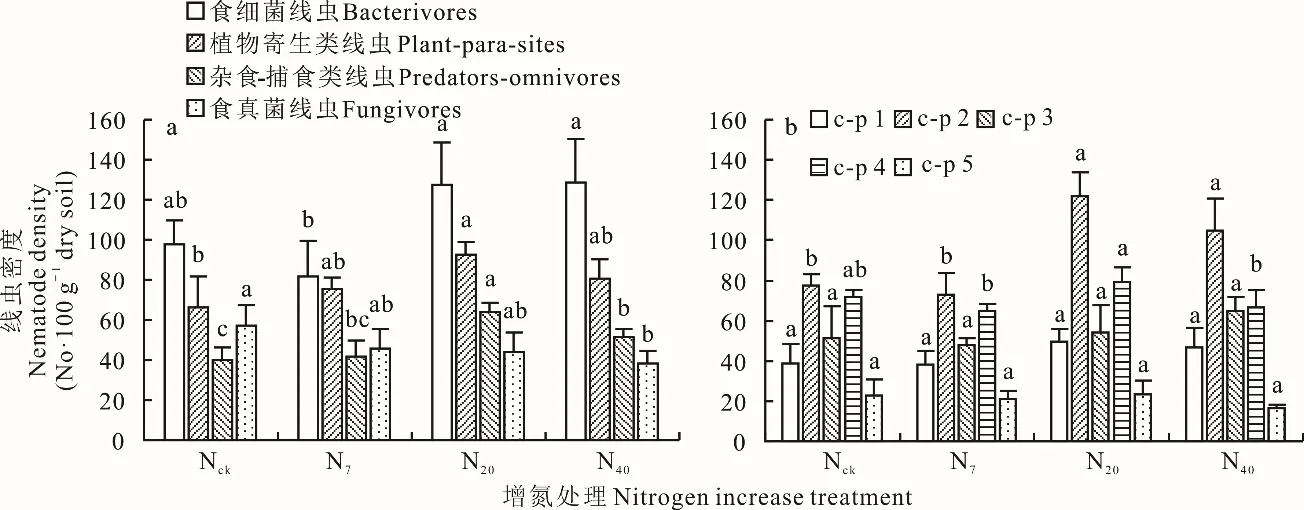
图2 增氮处理下高寒草甸不同类群土壤线虫密度Fig.2 The soil nematode density of different taxa in alpine meadows under nitrogen increase treatment
2.2 增氮对土壤线虫多样性的影响
在不同处理下,线虫多样性指数(H′)和均匀度指数(J)均表现为Nck>N40>N20>N7,增氮显著改变了线虫多样性和均匀度(P<0.05),且线虫多样性指数和均匀度指数在N7处理中下降最为明显,表明低水平的增氮处理降低了土壤线虫的分布均匀度和线虫群落结构的稳定性(表2)。
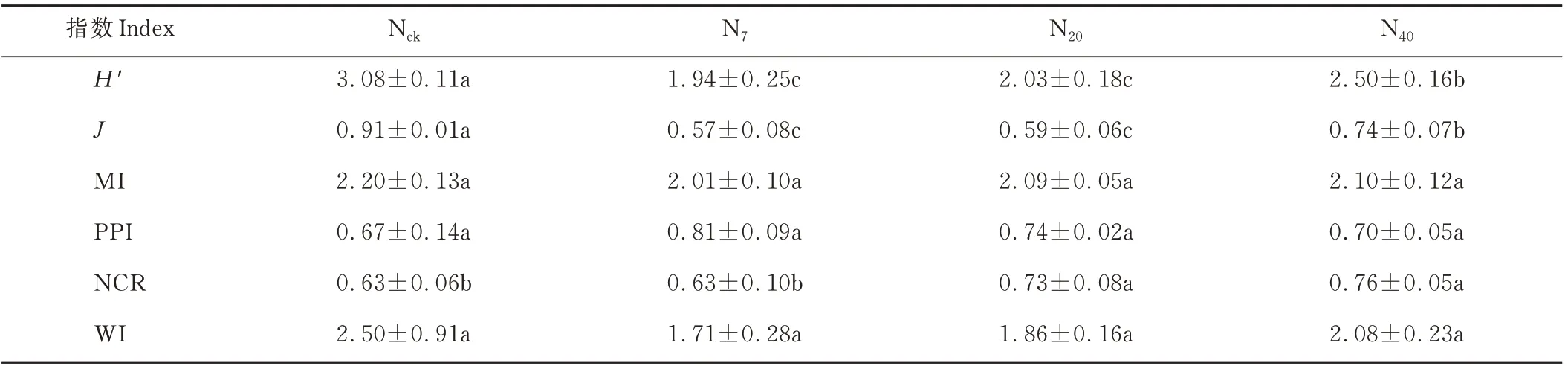
表2 增氮处理下高寒草甸土壤线虫生态指数Table 2 Nematode ecology indices under nitrogen increase treatment in alpine meadow
不同处理间植物寄生线虫指数(PPI)、自由线虫成熟度指数(MI)、瓦斯乐思卡指数(WI)无显著异,N20、N40显著增加了线虫通道指数(NCR)(P<0.05),在不同增氮条件下,线虫通道指数均大于0.5,瓦斯乐思卡指数均大于1。
2.3 增氮对植物群落特征和土壤氮含量的影响
N40和N20均显著增加了高寒草甸禾莎草和杂类草的高度(P<0.05),具体表现为:N40>N20>Nck(表3),N7处理对植物高度无影响,并且施氮对样地的植物地下生物量无显著影响。与Nck相比较,N20和N40条件下禾莎草盖度增幅分别达34.6%和69.9%,N7和N20条件下杂类草盖度显著增加(P<0.05),而N40处理对杂类草盖度无显著影响。与Nck相比较,N7、N20和N40条件下地上生物量增幅分别达59.0%、122.3%和139.7%。此外,N20、N40显著提高了土壤中氨态氮和硝态氮含量(P<0.05)。

表3 增氮处理下高寒草甸植物群落特征和土壤氮含量Table 3 Plant community characteristics and soil nitrogen content under nitrogen increase treatment in alpine meadow
2.4 土壤线虫群落及其影响因素分析
土壤线虫数量与地上生物量、禾莎草盖度、植物群落高度等存在相关性(图3),其中食细菌线虫数量与土壤氨态氮及杂类草高度呈极显著正相关(P<0.01),植物寄生类线虫数量和禾莎草高度呈显著正相关(P<0.05),而食真菌线虫数量与地上生物量及杂类草高度呈显著负相关(P<0.05);土壤线虫总量与食细菌线虫数量、植物寄生类线虫数量、杂食-捕食线虫数量以及植物地上生物量呈极显著正相关(P<0.01),与土壤中氨态氮、禾莎草盖度及植物群落高度呈显著正相关(P<0.05);此外,食细菌线虫数量和食真菌线虫数量显著负相关(P<0.05),增氮条件下两类土壤线虫存在竞争关系。在土壤含氮量和植物生产力方面,土壤中氨态氮、硝态氮含量和植物生物量无显著相关关系(P>0.05)。

图3 土壤线虫群落与植物群落及土壤氮含量相关性分析Fig.3 Correlation of soil nematode communities with plant community and soil nitrogen content
3 讨论
3.1 增氮对土壤线虫群落结构的影响
土壤线虫在高寒草地生态系统中数量众多,并且与植物、土壤微生物等联系紧密,共同维持着生态系统能量和物质动态平衡,是衡量生态系统健康的重要指标[27]。长期或短期增氮通过改变生物及非生物等因素,直接或间接地影响土壤线虫的生活环境[28]。以往的研究表明,植物寄生类线虫和食细菌类线虫是青藏高原高寒草甸土壤中主要的线虫类群,杂食-捕食类线虫和食真菌线虫所占的比例较低[29]。本研究证实了在对照条件下,以食细菌线虫和食真菌线虫为主的c-p 2类群线虫是高寒草甸线虫优势类群。在增氮条件下,体型较小的植物寄生类线虫垫刃属、针属和螺旋属等数量增加,成为高寒草甸土壤线虫中的常见属,这与在甘南草原进行的长期增氮研究结果一致[29]。在环境胁迫条件下,植物寄生类线虫在汲取植物营养的同时可以通过消化道分泌有毒有害物质,可能威胁植物的生长和土壤健康[30]。因此,需要进一步关注长期增氮条件下高寒草甸中植物寄生类线虫数量的变化及其对植物生产力的影响。
增氮可以改变高寒草甸土壤的理化性质,直接或间接地影响植物群落组成[31],进而影响土壤线虫的群落结构和密度[32]。有研究表明,长期增氮降低了青藏高原高寒草地植物群落多样性,显著增加了植物地上生物量[33]。葛怡情[34]在高寒草甸上的研究发现长期氮沉降对禾莎草植物生长有促进作用并且提高了土壤中氨态氮和硝态氮含量。相比较于长期的施氮试验,本研究施氮处理提高了植物地上生物量,高浓度施氮增加了禾莎草盖度,杂类草盖度在最高梯度处理下和对照组无差异,可能与氮沉降造成的杂类草光合作用降低有关[35]。其次,施氮条件下禾莎草群落盖度的增加对植物地上生物量的增加贡献很大,禾莎草群落盖度和食细菌线虫数量呈极显著正相关。闫俊[36]也认为增氮改变了青藏高原高寒草地线虫群落结构,增加了食细菌线虫数量,减少了食真菌线虫数量,这可能与增氮抑制了土壤中真菌菌丝的生长有关[37]。本研究明确了在增氮条件下,食细菌线虫数量的增加是藏北高寒草甸土壤线虫总量增加的主要来源。
3.2 增氮对土壤线虫多样性的影响
高寒草甸土壤动物多样性是保持草地生态系统稳定的重要条件[38]。相关研究表明,增氮显著增加了青藏高原高寒草甸土壤线虫多度,降低了土壤线虫物种丰富度[39]。胡靖[40]和王静[11]的研究均发现增氮增加了高寒草甸土壤线虫的数量但降低了线虫群落的多样性,这与增氮引起的草地植物群落的改变有关。同时,在我国内蒙古地区半干旱草原[4]和典型草原[41]上的研究表明,增氮导致土壤线虫数量下降并且减少了线虫的多样性。本研究结果表明,增氮处理中多样性指数(H′)和均匀度指数(J)显著低于对照,其原因可能是增氮改变了藏北高寒草地植被生产力和生态系统的食物链,使土壤中微生物群落和数量发生变化,进而影响了土壤线虫群落结构和多样性[42]。
土壤线虫生态指数可以在一定程度上综合反映环境变化对线虫群落结构和多样性带来的影响[26]。已有研究表明,增氮降低青藏高原东部草地土壤线虫自由线虫成熟度指数和瓦斯乐思卡指数[40]。在内蒙古退化草地上的研究结果表明增氮增加了自由线虫成熟度指数[43]。本研究发现,增氮对自由线虫成熟度指数、植物寄生线虫指数和瓦斯乐斯卡指数无显著影响,表明增氮没有改变植物寄生线虫对此样地土壤的适应性,地上植被生长良好,土壤线虫群落受增氮处理的影响较小。本研究中,线虫通道指数NCR>0.5,高水平增氮处理下(N20,N40)线虫通道指数NCR增加显著,说明本研究区域中土壤线虫类群整体以食细菌线虫为主,增氮促进了高寒草甸生态系统有机物通过细菌途径分解的过程。此外,各处理线虫WI>1,表明单位土壤中食微线虫数量比植物寄生类线虫多,矿化途径主要由食细菌真菌线虫参与[44]。
4 结论
高寒草甸土壤线虫的优势类群为食细菌线虫、植物寄生线虫和c-p 2类线虫,增氮显著降低线虫多样性和均匀度,增加高寒草甸土壤线虫密度和NCR,对PPI、MI、WI没有影响。因此,增氮尽管增加土壤线虫密度,但降低线虫多样性,并有可能改变有机物分解过程和土壤中的食物链关系。在后续研究中,应当结合植物残体和土壤微生物群落组成,分析长期增氮条件下植物和微生物等食物来源与土壤线虫群落结构之间的关系,有助于深入解析高寒草甸土壤线虫群落变化。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
今日农业(2022年14期)2022-09-15 01:44:26
青海草业(2022年2期)2022-07-23 09:34:58
今日农业(2020年19期)2020-12-14 14:16:52
活力(2019年21期)2019-04-01 12:17:10
中成药(2018年2期)2018-05-09 07:20:04
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
天然产物研究与开发(2016年6期)2016-06-05 10:29:30
中国科技信息(2015年2期)2015-11-16 08:18:32
