
鲫寄生7种指环虫的形态学和分子鉴定
2023-01-30 07:29:28程姜雯潘瑛子马幸荣王桂堂李文祥
水生生物学报 2023年2期
程姜雯 潘瑛子 马幸荣 王桂堂 李文祥
(1.西藏大学理学院, 拉萨 850000; 2.中国科学院水生生物研究所农业农村部水产养殖病害防控重点实验室, 武汉 430072;3.西藏自治区农牧科学院水产科学研究所, 拉萨 850032)
指环虫属(DactylogyrusDiesing, 1850)隶属于单殖吸虫纲(Monogenea)指环虫目(Dactylogyridae)指环虫科(Dactylogyridae), 是至今发现的物种多样性最高的一个属, 全世界已经发现900多种[1]。指环虫宿主特异性较强, 一般一种指环虫只寄生在一种宿主上[2]。绝大多数种类寄生于鱼类的鳃部, 刺激鳃分泌黏液, 影响呼吸, 甚至破坏鳃组织, 引起病毒、细菌与真菌的继发性感染, 导致鱼类的大量死亡, 造成严重的经济损失[3]。寄生于鲢(Hypophthalmichthys molitrix)的鲢指环虫(D.hypophthalmichthys)与小鞘指环虫(D.vaginulatus), 及寄生于草鱼(Ctenopharyngodon idellus)的鳃片指环虫(D.lamellatus)等都给我国的淡水鱼类养殖造成较大损失[4,5]。
鲫(Carassius auratus)是我国重要的养殖种类,在全国各地都有养殖, 随着养殖密度的提高, 指环虫病的发生日益严重和频繁, 成为水产养殖危害最严重的寄生虫病之一[6]。在我国, 报道的鲫鳃部寄生的指环虫有8种之多[7], Li等[8]在湖北省自然湖泊和养殖池塘的鲫中发现了7种指环虫, 分别为坏鳃指环虫(D.vastator)、中型指环虫(D.intermedius)、弧形指环虫(D.arcuatus)、弓茎指环虫(D.baueri)、美丽指环虫(D.formosus)、望外指环虫(D.inexpeatatus)和锚钩指环虫(D.anchoratus)。Tu等[9]利用形态特征和分子标记方法, 对华中地区养殖鲫(C.auratus)鳃上的美丽指环虫(D.formosus)进行了形态学重描述和系统发育分析。Ling等[10]比较了华中地区鲫的坏鳃指环虫和中型指环虫的形态特征差异, 及18S rDNA+内转录间隔区(ITS-1)的分子序列差异。
随着各国间的鱼类引种和水产品贸易, 鲫寄生的指环虫也被带入到世界各地。在日本的金鱼中发现了叉茎指环虫(D.dukeiti)、美丽指环虫、锚钩指环虫、弓茎指环虫、中型指环虫和坏鳃指环虫6种指环虫[11]; 在保加利亚发现了坏鳃指环虫、中型指环虫、弓茎指环虫、美丽指环虫和锚钩指环虫5种指环虫[12]; 在印度[13]、 伊拉克[14]、伊朗[15]、墨西哥[16]、巴西[17]、古巴[18]和澳大利亚[19]等国家的金鱼鳃上都报道了上述种指环虫, 而这些指环虫在中国都有报道。
一种鱼类寄生多种指环虫, 这种现象在拟鲤(Rutilus rutilus)和大头欧雅鱼(Squalius cephalus)中有报道过[20], 这也给指环虫的种类鉴定带来了困难。对于指环虫属的分类鉴定, 主要采用形态学结合分子鉴定[21]。由于鲫的有些指环虫种类的形态描述比较早, 且缺乏分子数据, 本研究通过对我国鲫寄生指环虫种类调查, 重新描述和测量形态学特征, 并提供分子分类数据, 为鲫寄生指环虫的种类鉴定提供参考依据。
1 材料与方法
1.1 指环虫样品收集
鲫采自湖北省武汉市梁子湖(30°11′05.460′′N,114°37′33.984′′E), 共50尾。将活鱼运回实验室, 暂养于水族箱中。将实验鱼麻醉(MS-222)后, 在解剖镜(Nikon C-DSS230)下, 用解剖针逐次分开鳃丝检查。在发现指环虫后, 用解剖针将其剥离, 放入清水中, 冲洗干净。将虫体活体直接在显微镜下观察、拍照, 并进行初步的形态鉴定, 一部分虫体用聚乙烯醇封片, 用作进一步的形态观察和测量;另一部分虫体用90%的酒精固定, 用作分子鉴定。
1.2 指环虫的形态学鉴定
在显微镜(OLYMPUS-CKX41)下观察虫体形态结构, 并利用电子目镜(Pro-MiceoScan 5630)进行拍照和形态描述。按照指环虫的测量参数和标准[7],利用软件(Scope Photo 3.0)对已经拍照的指环虫进行测量, 并对几丁质结构进行绘图。
1.3 指环虫的分子鉴定和系统发育分析
DNA提取、扩增与测序除望外指环虫与弓茎指环虫外, 每种指环虫选3只, 按照蛋白酶K、酚-氯仿抽提法提取指环虫基因组DNA并选用特异性引物[21]扩增部分18S rDNA、全部的ITS1 rDNA及部分5.8S rDNA等片段; 引物序列分别为上游引物S1 (5′-ATTCCGATAACGAACGAGACT-3′); 下游引物H7 (5′-GCTGCGTTCTTCATCGATACTCG-3′)或者IR8 (5′-GCTAGCTGCGTTCTTCATCGA-3′)。
PCR扩增体系为50 μL, 包括 20 ng左右的指环虫基因组DNA、1.25 U的ExTaq聚合酶(TaKaRa)、1 μL 20 mmol/L的上下游引物、4 μL 2.5 mmol/L dNTP溶液、 5 μL含20 mmol/L MgCl2的10×PCR缓冲液, 最后用灭菌的双蒸水定容到50 μL。扩增反应程序如下: 95℃ 预变性5min; 94℃变性1min,55℃退火1min, 72℃延伸1min30s, 35个循环; 72℃后延伸10min。
PCR产物在1%的琼脂糖凝胶中电泳分离, 切割目的条带并回收DNA, 然后用pMD18-T载体(TaKaRa)进行连接, 转化到DH5α感受态细胞中, 50 μL/mL氨苄青霉素的LB固体培养基均匀涂布并培养过夜。挑取阳性克隆, 再经过PCR筛选, 将阳性克隆菌液送华大基因测序公司进行测序。
分子鉴定将获得的指环虫的DNA序列, 在NCBI中进行BLAST比对分析, 通过与数据库中已提交的相似度最高的序列进行对比, 并比较其序列的碱基位点差异, 进一步确定指环虫的种类。
系统发育分析本研究共获得5种指环虫DNA序列, 另从GenBank获得其他用于构建系统发育树的指环虫DNA序列37条,Thaparocleidus vistulensis作为系统发育分析的外类群(表1)。将提取的序列导入MAFFT v7.313[22], 对DNA序列进行多重序列比对, 随后将序列导入PhyloSuite v1.2.2[23]的Gblocks进行修剪校正。用ModelFinder[24]软件对优化的数据进行最优模型选择。采用MrBayes v3.2.6[25]构建贝叶斯推论法(Bayesian inferences, BI)的系统发育树, 模型选择为GTR+F+I+G4, 马尔科夫链的蒙特卡洛方法(Markov chain Monte Carlo process,MCMC)设置为2条链独立运行2000000代。每1000代对系统树抽样, 前25%舍弃, 根据剩余样本构建一致树(Consensus tree)并计算相关参数, 以后验概率(Posterior probability, PP)来表示各分支的可信性。使用IQ-TREE v1.6.8[26]构建最大似然法(Maximum likelyhood, ML)的系统发育树, 自动选择最优模型,模型选择为TIM3+F+I+G4, 标准自展值设置为1000代。系统发育树的美化则通过iTOL v6 (https://itol.embl.de/)[27]及Adobe Illustrator 2021 25.0完成。

表1 系统发育分析中所用指环虫种名及其宿主种名、GenBank 登录号、18S+ITS1+5.8S rDNA长度及采样地点Tab.1 List of Dactylogyrus species used in the study and their host, GenBank accession number, 18S+ITS1+5.8S rDNA length and sampling location
此外, 在系统发育树的基础上, 选取部分坏鳃指环虫和中型指环虫序列, 基于双参数模型 (Kimuratwo-parameter) 来计算序列的遗传距离。
2 结果
2.1 指环虫的形态学鉴定
通过对鲫寄生指环虫的形态特征比较, 分别鉴定为坏鳃指环虫(D.vastatorNybelin, 1924)、中型指环虫(D.intermediusWegener, 1909)、弧形指环虫(D.arcuatusYamaguti, 1942)、美丽指环虫(D.formosusKulwiec, 1927)、弓茎指环虫(D.baueriGussev, 1955)、望外指环虫(D.inexpeatatusIsjumova, 1955)和叉茎指环虫(D.dulkeitiBychowsky,1936)。下面是7种指环虫的形态学特征描述, 形态测量均采用μm为度量单位。
坏鳃指环虫虫体体型较大, 长467—1016(n=8), 宽124—353。锚钩长30—72, 较粗壮, 钩柄长35—84。腹叶长13—44, 背叶长9—25, 钩尖长5—15, 呈波浪状。后吸器的7对边缘小钩发育完全,形状相似, 长27—77。联结棒单一, 长47—106, 宽4—15。交配管呈斜管状, 末端较尖, 长31—72, 后端带一基座, 长21—65, 宽6—24, 辅助交配器与交配管基部连接, 末端分两支, 其中一支横向朝前勾着交配管, 长26—73 (图1)。

图1 坏鳃指环虫整体及几丁质结构(a.坏鳃指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.背联结棒; d′.交配器)Fig.1 D. vastator and its sclerotized structures (a.whole worm; b.anchors and marginal hooks; c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
中型指环虫虫体长268—901(n=13), 宽108—251。锚钩长16—27, 较为纤细, 钩柄长15—25。腹叶明显长于背叶, 腹叶长8—16, 背叶长3—7, 钩尖长6—13。后吸器的7对边缘小钩发育完全, 形状相似, 长19—36。联结棒单一, 长23—39,宽2—4。交配管呈斜管状, 末端较尖, 长23—53, 后端带一基座, 长11—55, 宽6—22, 辅助交配器与坏鳃指环虫相似, 末端分两支, 其中一支横向朝前勾着交配管, 长22—59 (图2)。
弧形指环虫虫体较小(n=28), 长96—490,宽23—106。锚钩较长, 长81—160, 伸出体外很多,钩柄长52—85。腹叶较长, 长27—106, 背叶不明显,钩尖长20—74。后吸器的6对边缘小钩发育完全,第7对边缘小钩雏形, 发育完全的边缘小钩长15—37。联结棒单一, 相对较短, 长17—51, 宽8—11。交配管为一弧形细管, 基部较宽, 逐渐向末端变细,长30—76, 辅助交配器较短, 长23—53 (图3)。
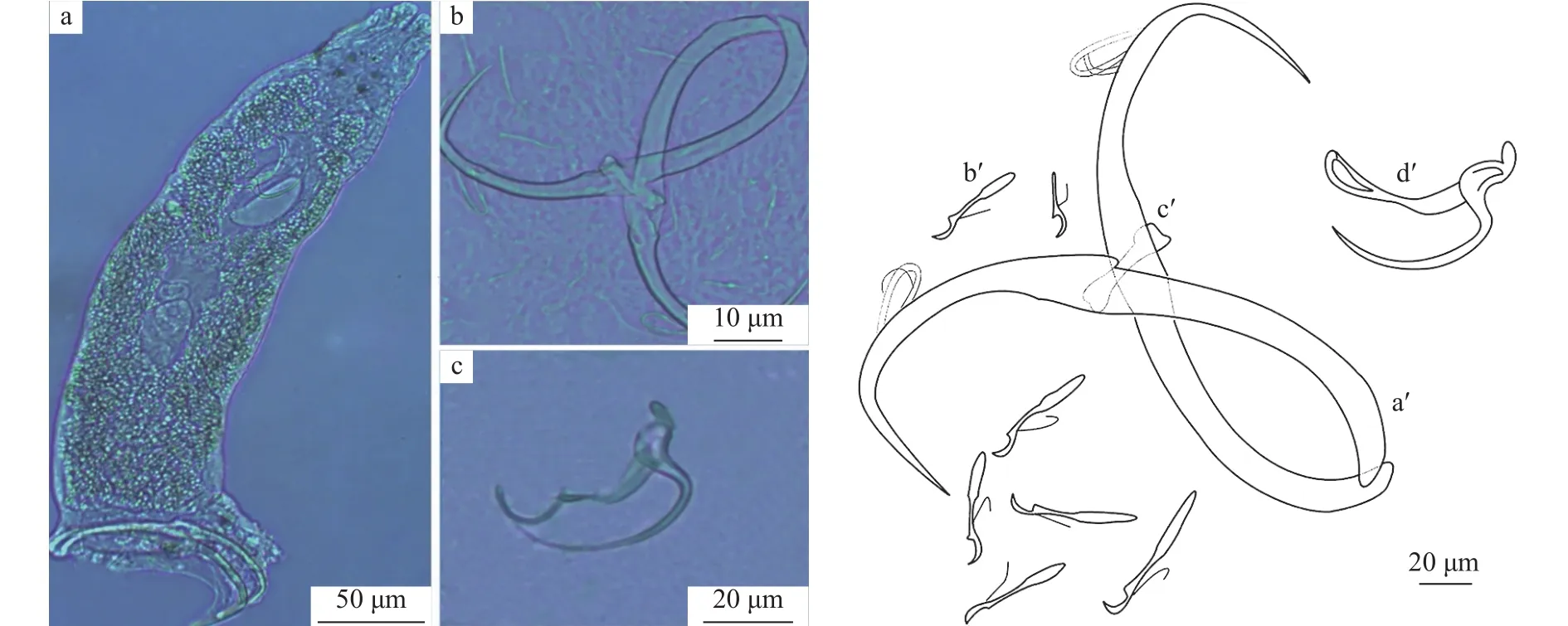
图3 弧形指环虫整体及几丁质结构(a.弧形指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.联结棒; d′.交配器)Fig.3 D.arcuatus and its sclerotized structures (a.whole worm; b.anchors and marginal hooks; c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
美丽指环虫虫体较小, 长218—462(n=11),宽48—97。锚钩较短, 长49—79, 钩柄长28—52。腹叶长24—45, 钩尖长10—22。后吸器发育完全的边缘小钩有6对, 形状相似, 第7对雏形, 长17—31。联结棒单一, 两端向内弯曲膨大, 类似马蹄形, 长18—42, 宽2—7。交配管较短, 呈斜管状, 末端较尖,长14—30, 后端带一基座, 呈贝壳状, 辅助交配器分三支, 其中一支略微朝前勾着交配管, 长11—25 (图4)。

图4 美丽指环虫整体及几丁质结构(a.美丽指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.联结棒; d′.交配器)Fig.4 D.formosus and its sclerotized structures (a.whole worm; b.anchors and marginal hooks; c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
弓茎指环虫虫体较小, 长120—369(n=6),宽19—73。锚钩较短, 长40—52, 钩柄长23—30。腹叶长18—30, 背叶不明显, 钩尖长16—21。后吸器具有7对发育完全的边缘小钩, 长14—21。联结棒单一, 较长, 中间有一波形, 长26—37, 宽2—4。交配器似弓形, 交配管为弧形管状, 长26—38, 辅助交配器分为3支, 前两支弯曲包着交配管, 长25—40(图5)。
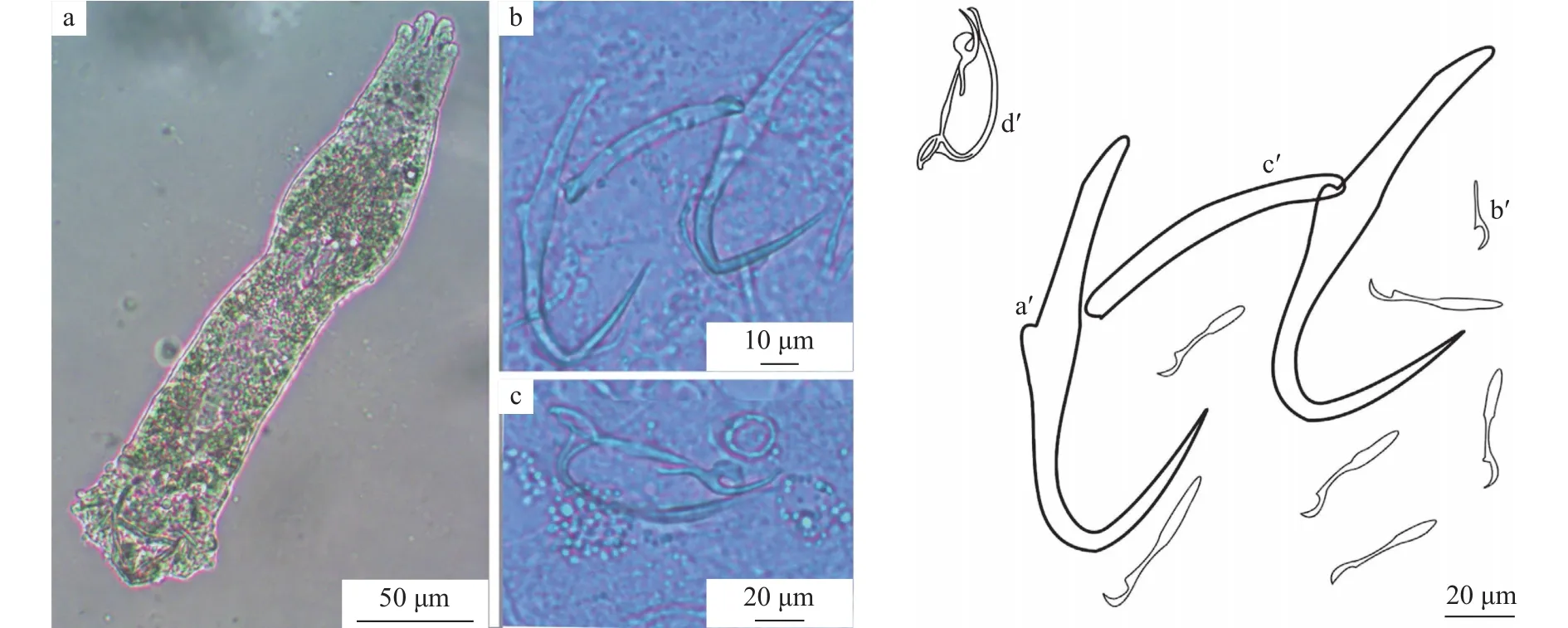
图5 弓茎指环虫整体及几丁质结构(a.弓茎指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.联结棒; d′.交配器)Fig.5 D.baueri and its sclerotized structures (a.whole worm; b.anchors and marginal hooks, c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
望外指环虫体型较小, 长110—301(n=6),宽16—57。锚钩较短, 长54—60, 钩柄长32—35。腹叶长25—34, 背叶不明显, 长2左右, 钩尖长21—26。后吸器具有7对发育完全的边缘小钩, 长15—25。联结棒单一, 较长, 长32—39, 宽2—4。交配器为管状, 末端细长, 长17—54, 辅助交配器呈棒状, 末端膨大, 上部分似尖刀状, 长20—36 (图6)。

图6 望外指环虫整体及几丁质结构(a.望外指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.联结棒; d′.交配器)Fig.6 D.inexpeatatus and its sclerotized structures (a.whole worm; b.anchors and marginal hooks; c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
叉茎指环虫体型较小, 长357—480(n=17),宽65—106。锚钩纤细, 长37—57, 钩柄长21—31。腹叶长19—25, 有稍稍扭转的现象, 背叶不明显, 钩尖长15—31。后吸器具有7对发育完全的边缘小钩,长15—31。联结棒细长, 长30—46, 宽2—4。交配管细长, 末端膨大, 长22—30, 辅助交配器底部细长,在上部分为两支, 呈树枝状, 整个轮廓略似“Y” 字形, 长18—33 (图7)。
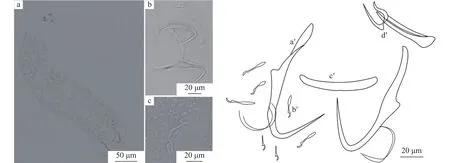
图7 叉茎指环虫整体及几丁质结构(a.叉茎指环虫整体; b.锚钩及边缘小钩; c.交配器; a′.中央大钩; b′.边缘小钩; c′.联结棒; d′.交配器)Fig.7 D.dulkeiti and its sclerotized structures (a.whole worm; b.anchors and marginal hooks; c.copulatory organ; a′.anchors;b′.marginal hooks; c′.dorsal bar; d′.copulatory organ)
2.2 分子鉴定与系统发育分析
本研究共获得坏鳃指环虫、中型指环虫、弧形指环虫、美丽指环虫与叉茎指环虫5种指环虫的18S+ITS1+5.8S rDNA序列。BLAST结果显示, 坏鳃指环虫的1045 bp的序列(KC876016)与捷克的坏鳃指环虫(AJ564159)相似度为96.37%, 存在38个位点差异; 中型指环虫的1013 bp的序列(KC876017)与捷克的中型指环虫(AJ564139)相似度为99.01%,存在10个位点的差异; 弧形指环虫的997 bp的序列(KC876019)与捷克的锚钩指环虫(D.anchoratus)(AJ564111)相似度为95.29%, 在GenBank中为首次提交; 美丽指环虫的1002 bp的序列(KC876018)与捷克的美丽指环虫(AJ564135)相似度为99.90%, 仅存在1个位点的差异; 叉茎指环虫的1049 bp的序列(KC876020)与捷克的叉茎指环虫(AJ564126)相似度为99.71%, 仅存在3个位点的差异。
用贝叶斯法和最大似然法构建的BI树和ML树树形完全一致, 支持率大小略有差别。不同地区的中型指环虫聚集在一起, 再与坏鳃指环虫形成一支;叉茎指环虫、弧形指环虫与美丽指环虫聚为一支,寄生于鲫的所有指环虫聚为一大支(BI 0.99/ML 100; 图8)。
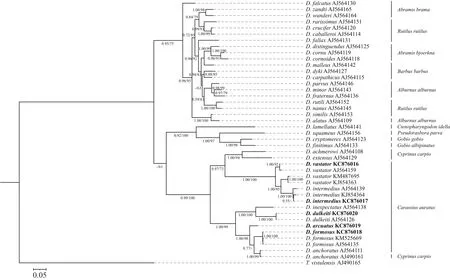
图8 基于18S rDNA+ITS1 rDNA联合序列构建的(BI/ML)指环虫系统发育树, 分支上的数字为贝叶斯后验概率(BPP)/最大似然法(ML)自展支持率Fig.8 Phylogenetic trees (BI/ML) of Dactylogrus spp.based on combined partial 18S rDNA and ITS1 sequences, rooted Thaparocleidus vistulensis.Numbers along branches represent the Bayesian posterior probability value and the bootstrap values of maximum likelihood
基于18S+ITS1 rDNA联合序列遗传距离分析显示, 坏鳃指环虫的种内遗传距离为0.004—0.058,中型指环虫的种内遗传距离为0.007—0.013, 坏鳃指环虫与中型指环虫的种间遗传距离为0.046—0.064(表2)。其中坏鳃指环虫(KJ854363与AJ564159)、(KM487695与AJ564159)和(KJ854363与KC876 016)的种内遗传距离(0.058/0.051/0.048)较大, 仅稍小于坏鳃指环虫与中型指环虫的种间遗传距离。进一步分析发现, 与18S区相比, 坏鳃指环虫的变异主要发生在ITS-1区(表3)。

表2 感染鲫的坏鳃指环虫和中型指环虫的18S+ITS1 rDNA联合序列基于双参数模型的遗传距离Tab.2 The genetic distances based on Kimura-two-parameter between Dactylogyrus vastator and Dactylogyrus intermedius infecting C.auratus, for combined partial 18S rDNA and ITS1 sequences
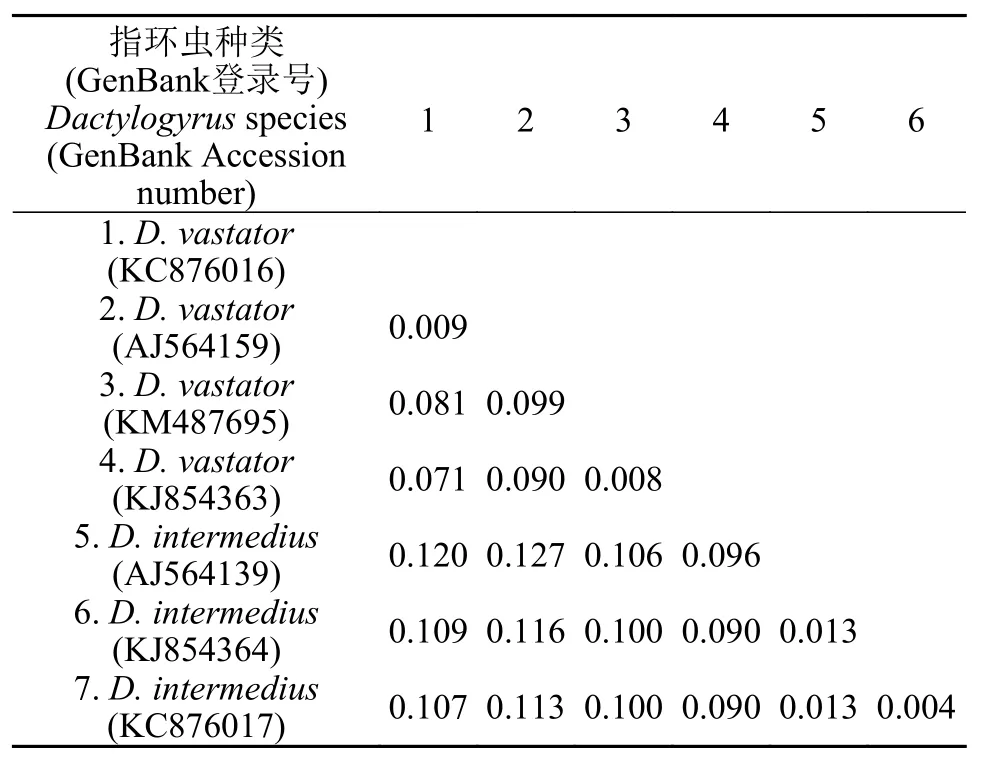
表3 感染鲫的坏鳃指环虫和中型指环虫的ITS1序列基于双参数模型的遗传距离Tab.3 The genetic distance based on Kimura-two-parameter between D.vastator and D.intermedius infecting C.auratus, for ITS1 sequences
3 讨论
通过形态学和分子鉴定, 在梁子湖的鲫鳃部共发现7种指环虫, 即坏鳃指环虫、中型指环虫、弧形指环虫、弓茎指环虫、美丽指环虫、望外指环虫及叉茎指环虫。
3.1 指环虫的形态学鉴定
形态上, 坏鳃指环虫与其他6种指环虫相比: 虫体体型较大, 其锚钩背叶和腹叶粗壮, 分叉明显, 靠近钩尖有波浪状突起。与国内外陈启鎏[7]、Ling等[10]、Ogawa等[11]和Borisov[12]报道的同种个体相比, 本研究报道的坏鳃指环虫锚钩总长(30—72vs.32—39vs.30—36vs.37—44vs.35—45)较长; 同时我们观察到坏鳃指环虫背联结棒有凹痕, Borisov[12]也报道了该现象, 而Ling等[10]报道的坏鳃指环虫其背联结棒较为光滑。
中型指环虫与其他6种指环虫相比: 其锚钩背叶和腹叶分叉明显, 但较为纤细; 辅助交配器末端分两支, 其中一只横向朝前勾着交配管。中型指环虫的后吸器几丁质结构的形态与坏鳃指环虫较为相似, 但坏鳃指环虫的锚钩总长(30—72vs.16—27)和背联结棒[(47—106)×(4—15)vs.(23—39)×(2—4)]比中型指环虫更大一些, 且钩尖呈波浪状。本研究报道的中型指环虫与国内外报道的同种个体的形态特征描述[7,10—12]基本一致。
弧形指环虫与其他6种指环虫相比: 其锚钩较长, 伸出体外很多; 具有较长的腹叶, 背叶不明显;交配管为弧形细管, 较为弯曲, 辅助交配器较短, 呈鞭状[7]。其锚钩与美丽指环虫和弓茎指环虫的较相似, 但弧形指环虫的锚钩更长(81—160vs.49—79vs.40—52)。与国内陈启鎏[9]报道的同种个体相比, 本研究报道弧形指环虫的背联结棒较大[(17—51)×(8—11)vs.(18—24)×(3—5)]。
美丽指环虫与其他6种指环虫相比: 其锚钩背叶不明显; 联结棒两端向内弯曲膨大, 类似马蹄形;交配管呈斜管状, 辅助交配器为树状分枝。美丽指环虫的后吸器几丁质结构的形态与弧形指环虫较为相似, 但美丽指环虫的锚钩总长仅为弧形指环虫的一半, 且通过交配器官形态可以区别这两种指环虫。与国内外陈启鎏[7]、Tu等[9]、Ogawa等[11]、Borisov[12]和Tancredo等[17]报道的同种个体相比, 本研究报道美丽指环虫的背联结棒较长(18—42vs.16—24vs.16—19vs.16—23vs.13—20vs.23—26)。
弓茎指环虫与其他6种指环虫相比: 其锚钩背叶不明显; 交配器似弓形。本研究报道的弓茎指环虫与国内外报道的同种个体的形态特征描述[7,11,12,17]基本一致。望外指环虫与其他6种指环虫相比: 其锚钩背叶不明显; 辅助交配器呈棒状, 末端膨大, 上部分似尖刀状。与陈启鎏[7]报道的同种个体相比,本研究报道望外指环虫的锚钩总长较长(54—60vs.37—48)。叉茎指环虫与其他6种指环虫相比: 其锚钩腹叶有稍稍扭转的现象, 背叶不明显; 辅助交配器底部细长, 在上部分为两支, 整个轮廓略似“Y”字形[7]。叉茎指环虫、望外指环虫和弓茎指环虫的锚钩形态相似, 主要依据交配器官形态来区分。
3.2 分子鉴定与系统发育分析
18S rDNA V4区和内转录间隔区(ITS1)现已成为指环虫分子鉴定的常用分子标记[10,28]。本研究共获得5种指环虫基因序列, 中型指环虫、美丽指环虫和叉茎指环虫与Šimková等[21]的研究中测得的相应的指环虫序列相似度高达99.0%以上, 与形态鉴定的结果一致。弧形指环虫为首次提交的基因序列, 所测得3个弧形指环虫的基因序列完全相同。坏鳃指环虫与捷克坏鳃指环虫(AJ564159)[21]相似度最高, 为96.37%。进一步基于18S+ITS1 rDNA联合序列的遗传距离分析发现, 坏鳃指环虫的种内遗传距离为0.004—0.058。Ling等[10]和Chaudhary等[13]也报道了坏鳃指环虫较低的18S+ITS1 rDNA联合序列相似性, 及较大种内变异的现象。在其他指环虫中也发现类似的现象, Benovics等[29]调查发现基于18S+ITS1 rDNA联合序列,D.folkmanovae的种内遗传距离为0.002—0.037,D.rarissimus的种内遗传距离为0.001—0.030。
较高的种内遗传变异可能与指环虫的宿主特异性有关。寄生于鲫的坏鳃指环虫的宿主除了同属的银鲫(C.gibelio)[30]和黑鲫(C.carassius)[31], 还有鲤科鱼类中的鲤(C.carpio)[7,32]、黑龙江野鲤(C.haematopterus)[32]、Cyprinion watsoni[31]、B.barbus[33]和Vimba vimba[34], 甚至在麦穗鱼(P.parva)[35]、棒花鱼(Abbottina rivularis)[35]和管臀鱼(Aulopyge huegelii)[36]上也有坏鳃指环虫的报道。这些调查结果表明坏鳃指环虫的宿主特异性并没有其他指环虫那么严格。Benovics等[29]在调查巴尔干地区鲤科鱼类寄生指环虫系统发育关系时, 发现低宿主特异性的指环虫具有广泛的种内遗传变异, 即使是在寄生于同一鱼类地理区域的同一宿主中的同种指环虫的两个种群之间, 也发生了种群间的遗传变异。Benovics等[37]还发现低宿主特异性的维氏指环虫(D.vistulae)的同一种群内的个体间也存在较高的遗传多样性。
宿主特异性是单殖吸虫与其宿主协同进化的驱动力, “宿主内成种”(Intrahost speciation)和“宿主转换”(Host switching)是单殖吸虫发生遗传变异和呈现物种多样性的重要进化事件, 这可能也是坏鳃指环虫存在较高种内遗传变异的原因。尽管坏鳃指环虫具有较高的种内遗传变异, 但其种内遗传距离仍小于坏鳃指环虫与中型指环虫之间的遗传距离。
猜你喜欢
科学大众(2021年8期)2021-07-20 07:38:52
四川蚕业(2021年1期)2021-02-12 02:03:12
好孩子画报(2020年10期)2020-11-02 02:52:37
昆虫学报(2020年6期)2020-08-06 06:42:52
歌剧(2017年12期)2018-01-23 03:13:33
作文周刊·小学三年级版(2017年22期)2017-07-05 08:17:56
时尚北京(2017年2期)2017-02-28 19:01:12
童话王国·文学大师班(2016年10期)2016-02-28 16:46:36
中学生物学(2016年8期)2016-01-18 09:08:26
中国马铃薯(2015年3期)2015-12-19 08:03:56
