
马泰勒虫AMA1基因的真核表达与鉴定
2023-01-17 09:01:12范士龙王水怡刘丹丹王金明李思媛刘明明刘雨桐巴音查汗
中国兽医学报 2022年12期
范士龙,芦 星,王水怡,刘丹丹,王金明,李思媛,刘明明,刘雨桐,巴音查汗,张 伟
(新疆农业大学 动物医学学院,新疆 乌鲁木齐 830052)
马泰勒虫(Theileriaequi,T.equi)是一种寄生在马属动物红细胞及网状内皮细胞内、由硬蜱传播的血液性原虫,与马驽巴贝斯虫(Babesiacaballi)一起统称马梨形虫[1]。马梨形虫病在世界范围内广泛流行,世界动物卫生组织(OIE)于2008年将其列为B类疫病,属法定报告疫病,我国于1999年将其列为二类疫病[2]。该病典型临床症状包括贫血、黄疸、体温升高、呼吸困难和多器官衰竭,严重者导致马属动物死亡[3]。该病多呈急性经过,对引进赛马、纯种马以及幼驹的危害较大[4],成为困扰马匹规模化养殖业扩大发展的制约因素之一。目前,对马梨形虫病的防治尚无有效的治疗方法,主要依靠化学药物,但因化学药物毒性较大且易产生耐药性,治疗效果不佳[5]。
顶复门原虫的顶膜抗原1(apical membrane antigen 1,AMA1)由微线体分泌,存在于多种顶复门原虫中,是一种高度保守的入侵相关蛋白,在裂殖子和子孢子入侵宿主细胞过程中起重要作用[6]。大量研究表明,顶复门原虫AMA1胞外区与棒状体颈部蛋白2(rhoptry neck protein 2)结合后,能够继续结合其他棒状体颈部蛋白连接形成复合物,进而形成运动连接环(moving juction,MJ)结构,直接介导虫体入侵宿主细胞过程[7-8]。因此阻断两者之间的相互作用可为抑制虫体的入侵提供新的参考思路[9]。研究发现,AMA1抗体通过破坏其与RONs的相互作用从而达到抑制裂殖子入侵,且相关研究已在疟原虫(Plasmodium)和弓形虫(Toxoplasma)中得到了验证,AMA1蛋白也因此被视为重要的疫苗候选抗原[10-11]。但T.equiAMA1蛋白是否可作为预防虫体入侵宿主细胞的保护性抗原,目前尚未见相关报道。
为了探究T.equiAMA1基因及其编码的蛋白功能和结构,本试验以GenBank中公布的美国WA株AMA1基因序列为目的基因设计特异性引物,以采集自新疆地区的T.equi样本基因组DNA为模板,扩增出AMA1目的基因片段后并在HEK293T细胞中进行真核表达,利用AMA1原核重组蛋白制备多克隆抗体通过间接免疫荧光和Western blot鉴定AMA1真核重组蛋白的表达,研究结果为进一步利用AMA1蛋白开展T.equi抗虫疫苗研发和入侵宿主机制研究奠定基础。
1 材料与方法
1.1 样品、细胞及主要试剂T.equi阳性抗凝马全血采自新疆塔城地区。pEGFP-N1载体、pMD19-T、E.coliDH5α、Prime STAR Max Premix(2×)、T4DNA连接酶、XhoⅠ、EcoRⅠ、DNA Marker等均购自宝生物工程(大连)有限公司;人胚肾细胞(HEK293T)购自武汉普诺赛生命科技有限公司;Lipfectamine 2000 Transfection Kit购自赛默飞世尔科技有限公司;DNA片段纯化回收试剂、质粒小量提取试剂盒购自OMEGA生物试剂公司;血液/组织 DNA 提取试剂盒、辣根过氧化物酶(HRP)标记的兔抗马IgG、DAB辣根过氧化物酶显色试剂盒购自北京博奥森生物技术有限公司。
1.2 引物设计与合成用GenBank中公布的T.equistrain WA(XM_004833042.1)AMA1基因序列为目标,下载Theileriaorientalis(NC_025260.1)、Theileriaannulata(KX231671.1)、Theileriaparva(NC_007344.1)基因组数据后,进行本地BLAST比对,获得泰勒虫属AMA1保守基因序列。利用 Primer Premier 5.0 软件针对该基因片段设计特异性引物(表1),其中划线部分为酶切位点EcoRⅠ和XhoⅠ,送上海生工生物工程公司合成。

表1 AMA1基因引物序列
1.3AMA1基因克隆及测序按照动物全血基因组DNA提取试剂盒说明书提取T.equi基因组DNA。以基因组DNA为模板进行PCR扩增,PCR扩增产物经纯化、回收后与载体pMD19-T连接,转化到E.coliDH5α,37℃过夜培养,将PCR鉴定为阳性的转化子送往上海生工生物工程公司进行测序。
1.4AMA1基因核苷酸和氨基酸序列相似性分析利用MEGA 7.0软件分析T.equi新疆株与其他顶复门原虫AMA1基因的核苷酸序列,构建Maximum parsimany(MP)系统发生树,分析以AMA1基因为标记的顶复门原虫分类进化关系。此外,用DNAStar Lasergene v7.1软件分别对AMA1基因的核苷酸和氨基酸序列进行比对,分析其核苷酸和氨基酸序列相似性。
1.5AMA1基因真核表达载体的构建利用XhoⅠ和EcoRⅠ限制性内切酶分别酶切基因和载体,回收后进行连接和转化,对菌液进行质粒提取以及测序和酶切鉴定,构建pEGFP-N1-AMA1真核表达质粒。
1.6 重组质粒转染参考Lipfectamine 2000 Transfection Kit说明书,将pEGFP-N1-AMA1 重组质粒转染到6孔板培养的HEK293T细胞中。将细胞培养板于37℃、5%CO2培养箱中培养4~6 h,更换为不完全DMEM培养基(含4%胎牛血清,不含双抗),继续培养36~48 h后进行下一步检测。
1.7 间接免疫荧光与Western bolt 验证转染pEGFP-N1-AMA1 重组质粒的HEK293T细胞在37℃、5%CO2细胞培养箱中培养 24 h后,使用5%多聚甲醛固定15 min,1%Triton X-100室温下处理15 min,PBS 洗涤3~5次。用5%脱脂乳 4℃封闭过夜,PBST 洗涤3~5 次。以鼠抗GFP单抗作为一抗,室温孵育1 h,PBST洗涤3~5次。用1∶500稀释的CoraLite594山羊抗鼠二抗室温孵育1 h,用PBST洗涤3~5次。DAPI孵育10 min,PBST洗涤3~5次,倒置荧光显微镜下观察结果。
收集转染 48 h 后pEGFP-N1-AMA1组和pEGFP-N1组细胞,加入WIP裂解液(PMSF∶RIPA=1∶100),置于冰上裂解细胞 25 min,每隔 5 min 吹打以充分裂解细胞。裂解后于 4℃、12 000 r/min 离心15 min 收上清。将处理的蛋白样品进行 Western blot分析,200 mA恒流、35 min 湿转印到 PVDF 膜上。加入 5%脱脂奶粉溶液,放置4℃ 封闭过夜,加入 GFP单抗和 AMA1鼠多抗室温摇床反应 2 h;经 PBST 充分洗涤后,再加入 HRP 标记的山羊抗鼠二抗室温摇床反应 1 h,充分洗涤后进行 DAB显色。
1.8 多克隆抗体制备GST-AMA1重组蛋白已在前期试验中制备并纯化,具体方法参见文献[12]。将前期纯化好的GST-AMA1重组蛋白用BCA蛋白浓度测定试剂盒检测浓度,步骤参照说明书;与相同体积的弗氏完全佐剂混合,以50 μg/只的剂量对小鼠进行皮下注射,共进行4次;4次免疫后7 d收集动物血液,分离血清,收集并分装上清,即得抗GST-AMA1的多克隆抗体,存放于-20℃备用。
1.9 多克隆抗体的效价测定及Western blot 鉴定利用间接 ELISA方法测定所产生多抗的效价,采用方阵滴定法筛选抗原包被的最佳浓度后加入酶标板各孔中,4℃过夜包被后,封闭液封闭2 h,洗涤3 次;用封闭液将上述收集的免疫后小鼠血清做 1∶100 稀释后,再做倍比稀释,同时设立阴性对照(免疫前小鼠血清)及空白对照(封闭液),封板,37℃孵育 1 h;加入稀释过的二抗(HRP 标记的山羊抗鼠 IgG),37℃孵育 1 h;加入底物显色液显色 10 min,每孔加入终止液 100 μL,测定D450 nm值。抗体样品孔的读数值除以阴性孔的读数值,结果大于该值即可判定为阳性。
SDS-PAGE结束后,将蛋白转印至 PVDF 膜上并进行封闭,将小鼠多抗血清按 1∶200 的比例稀释作为一抗,二抗使用 HRP 标记的山羊抗鼠 IgG,使用DAB显色液进行显色。
2 结果
2.1T.equiAMA1基因克隆及真核表达载体的鉴定结果对pMD19T-AMA1菌液进行PCR扩增,获得约为768 bp目的片段(图1A);将阳性菌液进行测序,序列比对结果显示,与GenBank收录的T.equi美国WA株AMA1基因(XM_004833042.1)核苷酸和氨基酸序列同源性分别为76.34%和78.64%。双酶切结果显示,AMA1目的基因插入的大小和pEGFP-N1载体位置均正确,表明AMA1基因已正确克隆到真核表达载体上(图1B)。

A.pMD19T-AMA1菌液PCR结果;B.pEGFP-AMA1酶切结果。M.DL2000 DNA Marker;A1,A2.样品;B1.样品;B2.空载
2.2AMA1基因的遗传进化分析结果以AMA1基因为标记的进化树聚类关系符合物种分类,核苷酸与氨基酸进化关系一致,不同种属的原虫被分为3个分支,在进化关系上T.equi新疆株聚类到泰勒虫属,与T.equi美国WA株位于同一支,两者进化关系最为接近(图2,3)。

图2 基于AMA1基因核苷酸DNA序列构建的顶复门原虫系统发育树

图3 基于AMA1蛋白氨基酸序列构建的顶复门原虫系统发生树
2.3T.equiAMA1真核重组蛋白间接免疫荧光与Western bolt 鉴定结果间接免疫荧光结果显示,pEGFP-N1-AMA1质粒转染的细胞中可观察到GFP自带的绿色荧光信号,以鼠抗GFP单抗作为一抗时,可在594 nm光源下观察到红色荧光信号(图4)。Western blot检测 AMA1 蛋白的表达情况,结果显示(图5),在以鼠抗GFP单抗作为一抗时,空载对照组在约30 kDa出现条带;以鼠抗GST-AMA1多克隆抗体作为一抗时pEGFP-N1-AMA1质粒转染组在约55 kDa处出现特异性条带,与预期大小相符。结果表明,AMA1蛋白已经在293T细胞中正确表达。
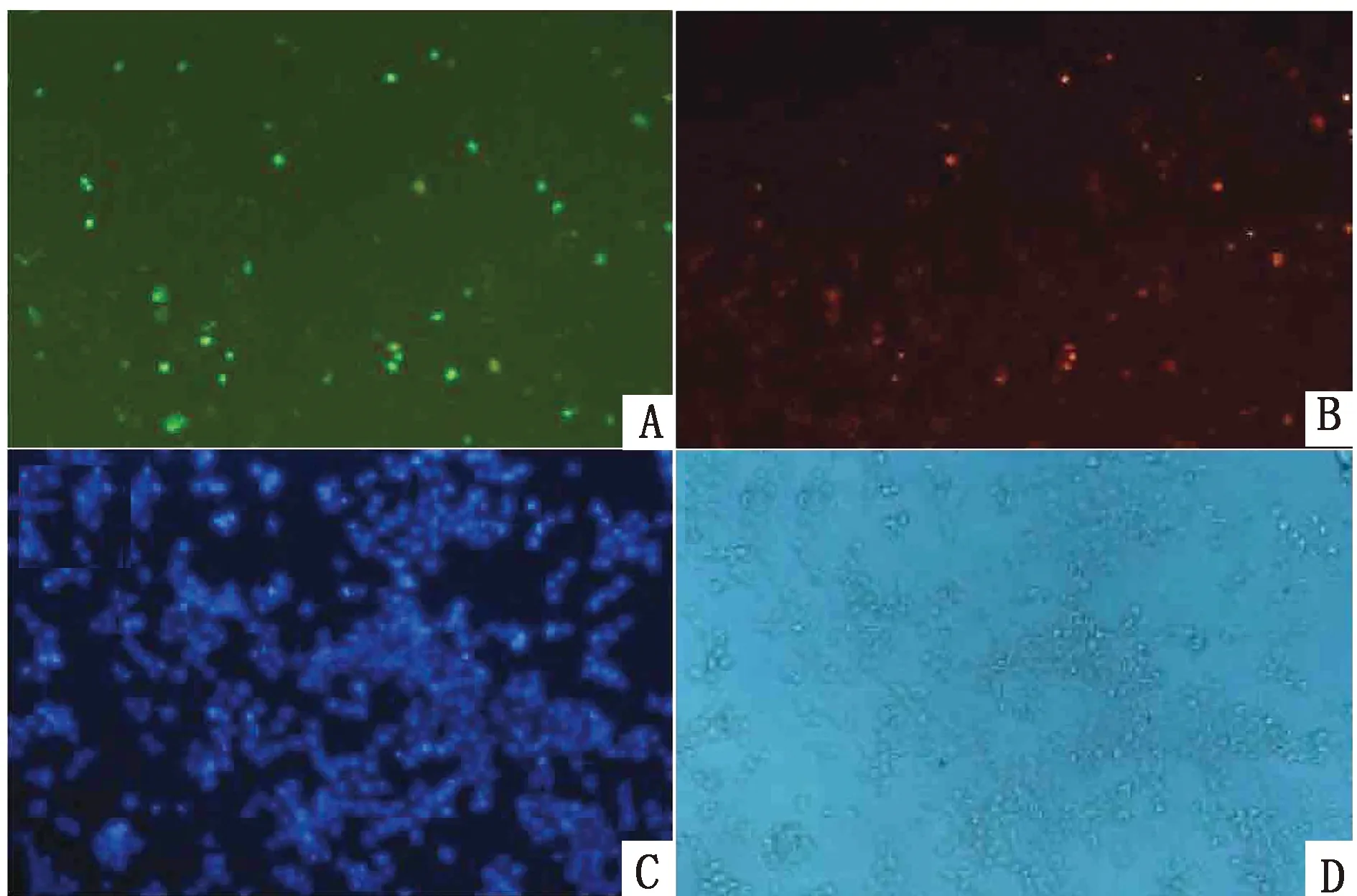
A.GFP绿色荧光;B.间接免疫Cy3荧光;C.紫色荧光;D.无荧光

A.EGFP标签Western blot结果;B.GFP-AMA1 Western blot结果。M.120 kDa 蛋白 Marker;A1.EGFP标签;B1.EGFP-AMA1蛋白
2.4 GST-AMA1多克隆抗体效价测定及Western blot 鉴定结果以2 mg/L重组抗原包被酶标板,利用间接ELISA检测其血清抗体效价,结果显示阴性血清的平均值为0.150,根据P/N2.1计算阳性的临界值为0.315,其效价约是1∶819 200(图6)。Western blot结果显示,在约60 kDa处有一明显条带(图7),表明T.equi GST-AMA1重组蛋白可与其免疫小鼠获得的多克隆抗体产生特异性反应。

图6 AMA1重组蛋白小鼠免疫后效价检测

M.蛋白 Marker;1.样品
3 讨论
AMA1蛋白是一种由顶复门原虫微线体分泌的蛋白,在虫体入侵宿主细胞时,释放并暴露在整个虫体表面[13]。研究发现,顶复门原虫在与宿主细胞膜接触后,所释放的微线体蛋白直接参与入侵宿主细胞的识别、黏附以及入侵过程[14]。目前,关于梨形虫AMA1蛋白报道较少,其具体功能还未深入探究,仅在东方巴贝斯虫(Babesiaorientalis)和牛巴贝斯虫(Babesiaboives)中有过相关AMA1蛋白的报道,研究发现在上述2种巴贝斯虫中,通过制备AMA1蛋白的多克隆抗体,将巴贝斯虫裂殖子与AMA1重组蛋白多克隆抗体孵育之后,可使裂殖子对红细胞的侵袭效率明显降低[15-16]。
本试验成功获得T.equiAMA1真核重组蛋白(约为55 kDa),其可与多克隆抗体产生特异性反应,多克隆抗体效价为1∶819 200,具有良好的抗原性。AMA1蛋白优良的免疫保护效果已经在许多顶复门原虫中得到了验证,而且多种与之相关的疫苗相继被研究报道。HOAN等[17]和王晔等[18]曾利用真核表达质粒pCAGGS-Et AMA1、pVAX1-Eb AMA1以及原核AMA1重组蛋白观察AMA1蛋白抗球虫的免疫保护试验,证明AMA1蛋白对鸡球虫感染均具有一定的免疫保护力,真核质粒组的卵囊减少率为74.55%,AMA1重组蛋白组的卵囊减少率为47.74%;使用真核质粒EtMIC2-pcDNA免疫后的试验结果显示,鸡体质量增加且鸡球虫卵囊孵化率降低[19]。此外,基于AMA1抗原的疟原虫疫苗已经在非洲疟疾流行地区进行了2期临床试验[20]。研究证明,AMA1免疫能促进IFN-γ的产生,比MIC2、M2AP基因免疫有更好的保护效果[21],且AMA1为交叉抗原,可利用其构建弓形虫与新孢子虫(Neosporacaninum)联合疫苗[22-23]。以上研究表明,AMA1蛋白在顶复门原虫病的防治工作中具有良好的研究前景,因此,开展T.equiAMA1蛋白的免疫保护相关研究,对于进一步研制抗T.equi疫苗具有重要的应用价值。目前,关于T.equiAMA1蛋白的特异性抗体是否能够与其他顶复门原虫一样,可以对宿主起到保护作用,还有待于进一步研究探索。
此外,研究发现AMA1蛋白在顶复门原虫入侵宿主细胞的过程中也发挥着重要作用。王旭[9]和MICHELLE等[24]曾分别在柔嫩艾美尔球虫(Eimeriatenella)、分歧巴贝斯虫(Babesiadivergens)和新孢子虫中,通过免疫共沉淀、双分子荧光、pull down和酵母双杂交试验验证了AMA1和RON2相互作用关系。并且在刚地弓形虫和疟原虫中,通过AMA1蛋白和RON2蛋白制备的多克隆抗体,成功在天然虫体中定位到这两个蛋白,并证实两者产生了互作[20,25]。T.equiAMA1蛋白是否与其他顶复门原虫的AMA1蛋白一样,参与到虫体对宿主细胞的入侵过程中,并且与它们产生相互作用,还有待于进一步研究。
综上,本试验成功获得T.equi新疆株AMA1基因片段,并且成功在真核表达系统中表达并鉴定;获得的AMA1真核重组蛋白可与多克隆抗体产生特异性反应,且多克隆抗体效价水平较高,可将AMA1蛋白作为抗T.equi候选保护性抗原。本研究为今后T.equi疫苗开发提供了候选靶点,为探究T.equi入侵宿主细胞机制奠定了理论基础。
猜你喜欢
昆明医科大学学报(2021年5期)2021-07-22 07:31:58
中国动物传染病学报(2021年3期)2021-07-21 03:20:06
新农业(2020年18期)2021-01-07 02:17:10
科学(2020年3期)2020-11-26 08:18:22
当代水产(2020年3期)2020-06-15 12:03:02
兽医导刊(2015年9期)2016-01-04 12:00:04
实用皮肤病学杂志(2015年4期)2015-12-22 11:21:42
医学研究杂志(2015年12期)2015-06-10 06:57:46
川北医学院学报(2015年5期)2015-02-22 14:16:28
现代检验医学杂志(2014年2期)2014-02-02 02:40:34
