
贵州习水国家级自然保护区习水杜鹃种群空间分布格局和生态位特征分析*
2023-01-16 09:17李崇清
贵州科学 2022年6期
王 逍,李崇清,罗 康,穆 璁,杨 瑞
(1贵州习水国家级自然保护区管理局,贵州 习水 564600;2.贵州大学林学院,贵州 贵阳 550025)
种群空间分布格局是指组成种群的个体在其生活空间中的位置状态或布局,是物种的生物学特性、种内竞争、种间竞争、干扰以及群落内环境条件等综合作用的结果[1-2]。分析种群空间分布格局有助于认识种群的生物学特性、种内和种间关系、种群与环境因子之间的相互关系,不同物种空间分布格局的类型多不相同,而对于同一物种,在其不同年龄结构、生长环境和管理措施等情况下,其空间分布格局的类型也有差别[3-4]。开展不同生长阶段空间格局的研究可以为揭示种群的发展潜在过程提供深入的视角,便于理解群落的空间结构和时间动态过程[5-6]。生态位是一个种在生态系统中和其生存的群落里的地位和状况[7]。研究种群生态位可以探讨群落内优势种的组成结构和空间分布,分析群落中种内和种间竞争来揭示种群生活对策和动态机制[8-9]。生态位理论除了有助于解释种群关系、群落物种多样性、群落进化过程、种群(群落)在生态系统中的地位及作用[10-11],还能一定程度揭示群落中物种共存、种间相互作用、环境对物种的影响等生态过程[12-15],从而有效预测植物种群的发展[16-17]。
习水杜鹃(Rhododendronxishuiense)是贵州习水国家级自然保护区(小地名:扬大老林)发现的杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)新植物种,目前仅见分布于贵州习水国家级自然保护区,该种近似于猫儿山杜鹃,但其花冠呈漏斗形,子房不被短柄腺体,叶较小[18]。本研究以贵州习水国家级自然保护区习水杜鹃所在森林群落调查数据为基础,采用Ripley’s K函数Levins和Shannon-Wiener生态位宽度指数和Pianka生态位重叠指数对习水杜鹃群落里各层次优势种的生态位进行量化,旨在探究习水杜鹃空分布成因及生态适应对策,为揭示习水杜鹃濒危原因和天然更新障碍机制,加强资源的保护、恢复和发展提供建议。
1 研究区概况
贵州习水国家级自然保护区(以下简称习水保护区),位于贵州省遵义市,地跨习水县和赤水市,地理位置为东经105°50′~106°29′,北纬28°07′~28°34′。保护区总面积51911 hm2,其中核心区面积21231.31 hm2,缓冲区面积11303.8 hm2,实验区面积19375.89 hm2,是贵州省面积最大的国家级自然保护区。保护区地形以山地为主,地貌主要为白垩系发育的丹霞地貌。年平均气温在10.3~17.2 ℃,年日照数1091~1246 h,年降水量770~1661 mm。植物种类有331科1001属2539种,其中分布有红豆杉(Taxuschinensis)、南方红豆杉(Taxuswallichianavarmairei)、福建柏(Fokieniahodginsii)、伯乐树(Bretschneiderasinensis)、鹅掌楸(Liriodendronchinense)、桫椤(Alsophilaspinulosa)等国家重点保护树种,近年来还发现了习水杜鹃、习水报春(Primulalithophila)、习水秋海棠(Begoniaxishuiensis)等新种。
2 研究方法
2.1 样地设置
因在习水保护区其他地暂未发现习水杜鹃的分布,结合实际,选择在现有习水杜鹃的分布地设置样地开展调查(图1)。但习水保护区地形特殊(高海拔的山脊),无法设置大面积样地,且习水杜鹃分布范围较小约300 m2,故设置10 m×30 m的样带开展调查。将样带分为3个10 m×10 m的乔木样地,12个5m×5m的灌木样地及草本样地进行统计分析。对样带内所有植物进行普查,记录所有木本植物种名称、胸径(地径)、树高、枝下高、冠幅等信息;草本植物记录物种名称、平均高度、盖度、多度等信息。以西南角为坐标原点,记录所有木本植物坐标位置,画草图标示样方位置和顺序,按样方顺序调查。另外用平板电脑、GIS软件对坡度、坡位、海拔等环境因子进行调查。

图1 习水杜鹃分布位置示意图Fig.1 Distribution of Rhododendron xishuiense
2.2 数据处理
2.2.1 种群空间分布格局分析
植物主要与其近邻相互作用,小尺度空间结构在植物生态学中具有重要意义[19]。Ripley于1977年首次提出点格局分析,该方法能够分析任意尺度下的空间分布格局,相比于传统格局分析法有明显的优越性,该方法在研究濒危植物方面受到越来越多的关注[20-21]。
Ripley’s K函数公式如下:
式中,A为样地面积,n为样地中某一植物种群的个体总数;uij为i、j两点间的距离,r表示空间距离;Wij为以点i为圆心,以uij为半径的圆周长在面积A中的比例,目的是消除边界效应;当uij≤r时,Ir(uij)=1,否则Ir(uij)=0。
在实际的应用中,为了消除尺度依赖性和稳定方差,常采用L(r)代替K(r),其表达式为:
结果,若L(r)=0,为随机分布;若L(r)>0,为聚集分布;若L(r)<0,为均匀分布。采用完全随机模型对函数进行检验,并用Monte-Carlo拟合检验种群的空间分布状况,即计算L(r)=0时的上下包迹线坐标值(置信区间99%),模拟次数199。以空间尺度r作为横坐标,上下包迹线坐标值为纵坐标制图。基于种群实际分布的L(r)值与理论值对比,验证其属于哪种分布模式。若低于下包迹线,为均匀分布,若位于上下包迹线之间,为随机分布;若高于上包迹线,为聚集分布。
2.2.2 生态位分析
生态位研究有助于了解群落中不同物种之间的竞争与共存关系[22-23]。生态位宽度、生态位重叠度是常用的生态位量化指标[21]。其计算过程如下:
(1)重要值(IV)计算
乔木层
重要值(I)=(相对密度+相对频度+相对盖度)/3
灌木层
重要值(I)=(相对密度+相对频度)/2
草本层
重要值(I)=(相对密度+相对频度)/2
(2)生态位宽度计算
对各层次中重要值较大的优势物种,以多度为参数计算生态位宽度,采用Levins和Shannon-Wiener生态位宽度的计算方式,Levins(1968)生态位宽度(Bi)侧重于分析物种在样方中出现的概率情况,Shannon-Wiene生态位宽度(Ba)侧重反映种群个体对资源的利用程度。
Levins(1968)生态位宽度计算公式:
Shannon-Wiener生态位宽度计算公式:
式中,j表示样方,Pij为物种i在第j样方的个体数占该物种的所有个体数的比例,t为样方总数,Bi的值域为[1,t],若物种i只出现在一个样方中时,Bi=1,当物种i以近似的数量出现在所有样方中时,则Bi越接近t。Ba值域为[0,lnt]。Bi或Ba值越大,生态位越宽,反之则越窄。
(3)生态位重叠计算
Pianka生态位重叠指数能直观的反映出物种种对间生态位重叠及变化情况,其计算公式如下:
式中,Pij为物种i在第j样方的个体数占该物种的所有个体数的比例,Pkj为物种k在第j个样方的个体数占该物种的所有个体数的比例,t为样方总数,Oik域值为[0,1]。当生态位重叠值接近0时,说明两个物种种对间生态学相似性低,不存在竞争或者竞争很小;当越接近1时表明两物种种对间生态学相似性高,利用环境资源的方式相似,资源有限时竞争激烈。
种群空间格局采用R软件中spatstat包完成,生态位分析采用SPSS统计完成[24-25]。
3 结果与分析
3.1 种群空间分布格局
3.1.1 不同发育阶段的习水杜鹃种群的空间分布格局
采用径阶(地径)结构代替年龄结构的方式,结合调查所得数据,按0<Ⅰ≤5 cm,5<Ⅱ≤10 cm,10<Ⅲ≤15 cm,15<Ⅳ≤20 cm,20 cm<Ⅴ的径阶划分方式对习水杜鹃种群的径级进行统计,将龄级Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ依次划分为幼苗、幼树、中树、大树、老树。得到习水杜鹃种群不同发育阶段株树空间分布图(图2)。从图看出,该种群广泛分布在样地中,但在各个年龄阶段的植株密度不同,样地中甚至出现了无大树分布的情况,种群的密度由小到大依次为:大树、老树、中树、幼树、幼苗。结合不同发育阶段植株的位置变化不难发现,习水杜鹃在老树阶段(即个体建群初期)分布在样地的东北角,中树的时候扩展到西南角,幼树阶段占据了样地中部,最新出生的幼苗在样地西北部出现了分布,可以看出习水杜鹃种群具有一定的扩散能力,并且种群在不断扩张发展。
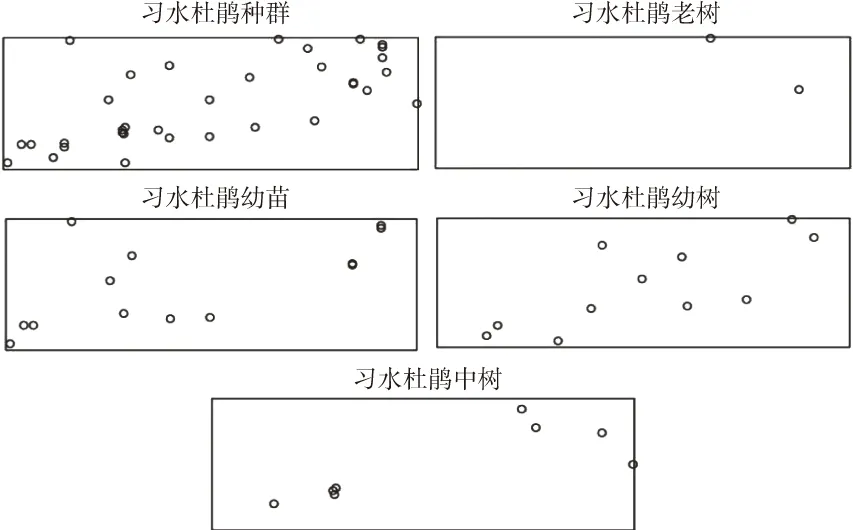
图2 习水杜鹃不同发育阶段的空间分布Fig.2 Spatial distribution of Rhododendron xishuiense at different development stages
习水杜鹃种群空间点格局(L(r)函数)分析结果如图3所示,习水杜鹃种群(a)、习水杜鹃幼苗(b)、习水杜鹃幼树(c)、习水杜鹃中树(d)L(r)基本位于置信区间内,总体上呈现显著随机分布,仅所有习水杜鹃个体在0.2~0.7 m尺度,习水杜鹃中树在0.3~1.2 m尺度上高于置信区间,表现为集群分布。随着龄级的增加,习水杜鹃的空间分布格局并未发生明显的变化。
3.1.2 习水杜鹃分布群落中优势种群空间分布格局
在习水杜鹃分布的群落中,对灌木层优势树种杜鹃,乔木层优势树种光枝杜鹃、桂南木莲、岭南石栎、小果润楠等优势种群空间位置和空间点格局进行分析(图4、图5)。从空间分布的位置可以看出,在300 m2的样地中,共分布了363株木本植物,密度大,其中光枝杜鹃(b)、桂南木莲(c)、岭南石栎(d)、小果润楠(f)更集中分布在某个较狭窄的区域,杜鹃(a)作为灌木层的绝对强势物种,分布范围相对广泛,但也倾向于几个小的集中区域。这与物种繁殖方式紧密相关,乔木层光枝杜鹃、桂南木莲、岭南石栎、小果润楠等因为种子掉落在母树下,所以表现出围绕母树的集中分布,灌木层杜鹃因其根萌蘖产生新的个体的繁殖方式,整体上较为集中。相比较以上的优势物种,习水杜鹃在空间分布的宏观比较上更加均匀。
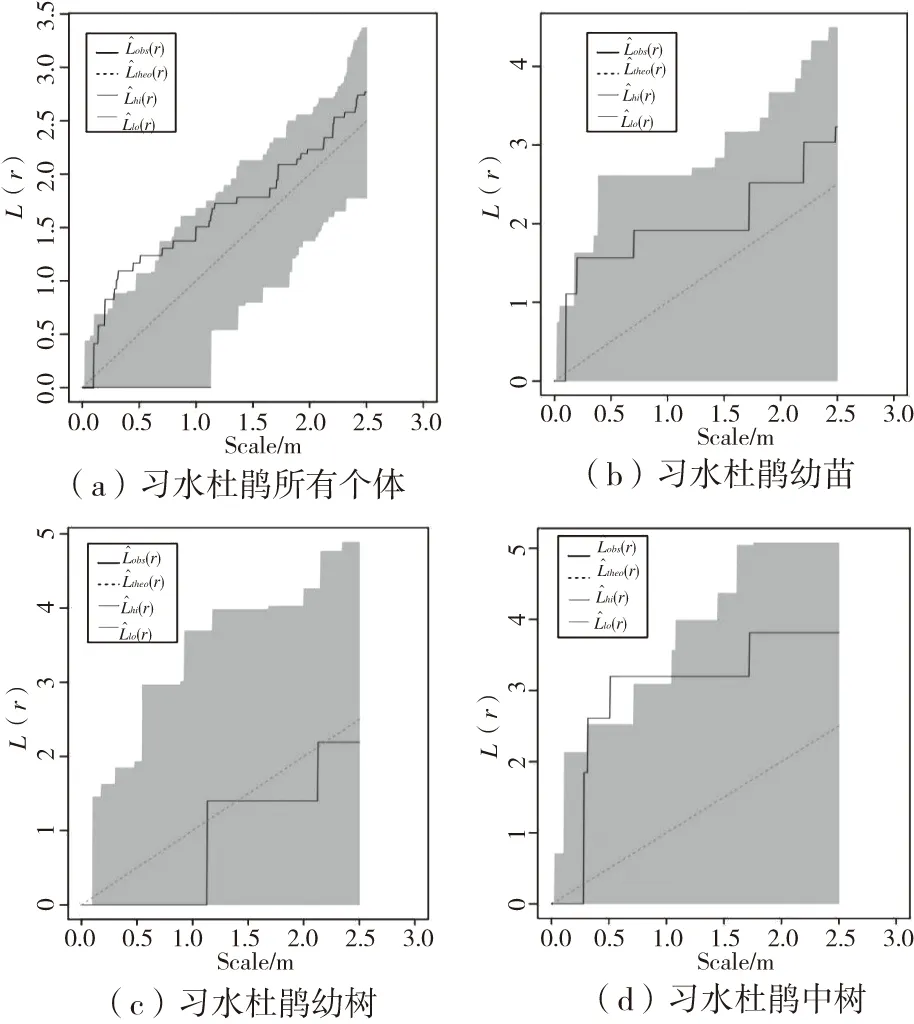
图3 习水杜鹃种群及不同发育阶段的空间点格局Fig.3 Spatial point pattern of Rhododendron xishuiense population at different development stages

图4 习水杜鹃群落中优势物种空间分布图Fig.4 Distribution of dominant species in Rhododendron xishuiense community
点格局分析结果与位置图的直观感受一致(图5),在小尺度的视角下,所有物种(h)在0~2.5 m整个空间尺度范围内L(r)值高于置信区间,呈现聚集分布,且L(r)值随着尺度的增大逐渐增加,表明聚集强度随尺度增大而增大。灌木层杜鹃表现出同样的聚集趋势。光枝杜鹃(b)在空间尺度内L(r)值基本位于置信区间内,空间分布表现为随机分布;桂南木莲(c)在0.6 m以下表现为聚集分布,大于0.6 m表现为随机分布;岭南石栎(d)在0.2~1.2 m表现为随机分布,在大于1.2 m和小于0.2 m的尺度下表现为聚集分布;小果润楠小果润楠(f)在0.2~1.4 m表现为随机分布,在大于1.4 m和小于0.2 m的尺度下表现为聚集分布。

图5 习水杜鹃群落中优势物种空间格局Fig.5 Spatial pattern of dominant species in Rhododendron xishuiense community
3.2 种群生态位特征
3.2.1 种群生态位宽度
在可利用资源有限的情况下,生态位宽度一般会增加,以使种群获得更多的资源,相反,在可利用资源丰富的环境中,可导致物种选择性利用资源,使得生态位变窄。其中生态位宽度较大的物种对资源利用更充分,对环境适应能力更强[26-27]。
由表1可看出,用不同的计算方法计算出的生态位宽度值结果呈现相同趋势。其中,在乔木层种,统计的重要值靠前的6个物种中,习水杜鹃生态位宽度值最大,Bi值和Ba值分别达到2.83和1.07,远超排在其后的硬壳柯(Bi值1.96,Ba值0.68)、石栎(Bi值1.80,Ba值0.64),约是其他乔木层物种的2倍,说明它在该层次中利用资源的竞争能力最强,环境适应性也最好。在灌木层中,统计重要值靠前的10个物种中,重要值最大的杜鹃Bi值和Ba值分别达到6.38和2.03,是其他灌木层植物的2~3倍,习水杜鹃在该层次中的Bi值和Ba值分别是3.24和1.27,仅次于杜鹃,但与杜鹃差距较大,且相较排在其后的江南樾桔(Bi值3.00,Ba值1.10)、贵州琼楠(Bi值3.00,Ba值1.10)差距较小,说明杜鹃生态位宽度在灌木层中占绝对的优势,而习水杜鹃在灌木层虽然有一定优势,但并不明显。在草本层中,箭竹Bi值和Ba值可分别达到4.39和1.65,是其他草本层植物的2~4倍,与灌木层杜鹃一样在同层次种占有绝对优势。对各层次内的生态位宽度比较还可以看出,乔木层的中厚皮香、岭南石栎、桂南木莲和灌木层中光枝杜鹃、南方荚蒾、格药柃、硬壳柯、石栎生态位宽度接近,反映出它们的竞争力相近,对环境资源的利用能力和环境适应能力具有相似性。
3.2.2 优势物种生态位重叠
当两个物种占有利用同一资源时,便会出现生态位重叠现象,在不同发育阶段,林分中种对间的生态位重叠状况不同。整个群落中,乔木层生态位重叠值如表2所示、灌木层生态位重叠值如表3所示、草本层生态位重叠值如表4所示。从以上表中可以看出,乔木层中优势种生态位重叠种对有15对,占比100%,习水杜鹃生态位与其他优势种重叠值均大于0.70,其中重叠最大的是岭南石栎达0.97,表明乔木层各物种间竞争较强,尤其习水杜鹃遭受的竞争最为突出。灌木层中优势种生态位重叠种对有21对,占物种种对的46.7%,其中习水杜鹃灌木竞争的物种有4种(重叠值小于0.4的占3种),占优势物种数44.4%,说明习水杜鹃在灌木层中遭受的竞争强度较为适中,草本层中优势种重叠有8对,占物种种对的80%。在各层次优势种生态位重叠值中,乔木层桂南木莲、厚皮香,草本层中蕙兰、矛叶荩草两对重叠值为1,表明它们生态学或生物学特性相近,拥有相似的资源利用方式,同时也存在极强的竞争关系;生态位重叠值为0的有26对,其中灌木层硬壳柯、草本层稀羽鳞毛蕨与多对多对物种间重叠值为0,表明它们占据着相对独立的资源,在环境不发生较大变化的情况下,相较其他物种更具发展潜力。对比物种的生态位宽度值和生态位重叠值,大多数物种生态位宽度表现与重叠值表现差异较小,而乔木层的桂南木莲生态位宽度较小,其生态位重叠值却在0.08~1之间浮动,灌木层杜鹃生态位宽度最大,但与其它灌木的生态位重叠值在0~0.86之间,草本层箭竹(在0~0.89之间浮动)也表现出同样的特征,这是说明它们与同层次物种间生态学或生物学特性存在差异性,从而与不同物种展现出不同的竞争强度。

表1 各层次优势种重要值及生态位宽度Tab.1 Importance values and niche widths of dominant species at all levels

表2 乔木层物种种对间生态位重叠值Tab.2 Niche overlap values between species pairs in the arbor layer

表3 灌木层物种种对间生态位重叠值Tab.3 Niche overlap values between species pairs in the shrub layer

表4 草本层物种种对间生态位重叠值Tab.4 Niche overlap values between species pairs in the herb layer
4 结论与讨论
不同发育阶段的习水杜鹃、乔木层光枝杜鹃空间分布格局表现为随机分布,乔木层桂南木莲、岭南石栎、小果润楠、灌木层优势种杜鹃在尺度范围表现为更多的聚集分布。竞争和空间异质性是形成种群空间格局的重要因素[28]。在环境因子中,地貌和气候对植物种群的空间格局影响最大[29-30]。在大的尺度下,气候和地貌特征往往存在较大差异,而小尺度消除了空间异质性(气候、地貌、土壤、湿度等)对植物的影响,提高了小地块上对空间格局分析的可靠性[31]。小尺度视角下的物种分布格局主要受种子扩散或萌蘖新苗等方式的影响[32]。本次研究在习水杜鹃的仅有的分布范围内,对所有物种进行了普查,全面准确的揭示了各物种在群落中的分布状况,从小尺度的视角对群落中优势种群的小尺度空间格局进行了分析。其中,习水杜鹃、光枝杜鹃是群落中同科同属小乔木,具有种子成熟后较轻的生物学特性,在风力等外界环境因子的作用下,易出现不规则的散落,故在空间尺度内表现为随机分布;灌木层杜鹃因丛生的生物学特性,萌蘖繁殖比种子繁殖方式能力更强,在空间尺度内表现为较强的聚集分布;桂南木莲、岭南石栎、小果润楠等物种种子多呈簇生状,且种子较重,不易受风力等外界因数的影响,故在小尺度视角下表现为围绕母树的更小的尺度内的聚集分布,较大的尺度内表现为随机分布。对比习水杜鹃各阶段的分布位置,可以看出习水杜鹃的种子具有一定的扩散能力,但草本层分布大面积的箭竹,加之林下枯落物丰富,使得散落的种子不易进入土壤中,能进入土壤中的种子受箭竹的影响也难以萌发生长,这与树枫杜鹃遭受的种群增长障碍相似[33]。从习水杜鹃幼苗的空间位置也可以看出,受群落中部山脊的密布箭竹影响,幼苗主要位于样地边缘。调查中还发现习水杜鹃种子易腐烂,花易遭受虫害,综合以上多种因素,导致了习水杜鹃种群当前的空间分布格局。而种子扩散及繁殖方式是影响群落中优势种群空间分布格局的主要原因。
物种濒危的主要原因是自身生存能力的缺陷、生物因素和非生物因素影响,物种间的竞争和人类活动是生物因素的主要组成部分[34]。在资源有限的环境中,共存物种之间对资源的竞争和分配加剧[35]。通过生态位宽度值大小结合生态位重叠情况,能够较好的解释物种遭受的竞争强弱。与重要值变化趋势一致,各层次中重要值最大的物种生态位宽度也占据优势,利用资源能力和环境适应性也最强。各层次生态位重叠物种种对占比乔木层(100%)>草本层(80%)>灌木层(46.7%),就数据而言,乔木层物种间竞争强度更大,且习水杜鹃与其他优势种生态位重叠值均大于0.70,竞争尤为突出,但综合考虑整个群落,乔木层能够获得更多的资源(如光照、发达根系对土壤养分的吸收等),而灌木层植物在资源有限的环境里,遭受的竞争应该更强。生态位多样化假说指出,物种共存需要沿着生态位轴(如光、土壤水分和有效养分)进行专门化,从而减少种间竞争使种群获得发展。因为这种专门化,小尺度上的共存物种生态位更相似[36-37]。乔木层中厚皮香、岭南石栎、桂南木莲,灌木层中光枝杜鹃、南方荚蒾、格药柃、硬壳柯、石栎等多种植物生态位宽度接近的可能就是专门化的体现。通过重要值和生态位重叠值对比还可以发现,与桂林岩溶石山常绿落叶阔叶混交林生态位研究结果一致[38],桂南木莲、杜鹃、箭竹生态位重叠值的浮动,说明重要值并不是影响生态位的唯一因素,还应考虑物种间生态学或生物学特性差异而展现出的竞争和适应的能力。如灌木层硬壳柯、草本层稀羽鳞毛蕨与多对物种间重叠值为0,他们通过占据着相对独立的资源,以获得相较于其他的物种更强的发展潜力。
总的来说,小尺度下可以忽略环境异质性对种群空间格局的影响,在相似的环境条件下,种群自身的生物、生态学特性对空间格局的形成过程中的作用得到充分的展现[32]。习水杜鹃种群的不同发育阶段的空间位置变化空间格局分析表明其种子具有一定扩散能力,但种子自身易腐烂和萌发受箭竹影响等多种因素,种群更新受到限制。多数关于濒危物种的研究均提出了人为干扰的措施,但也有研究表明,随着物种丰富度的增加,功能重叠增加,但平均功能总量没有下降[39]。自然保护区的管理更多倾向于尊重自然、顺应自然、保护自然,本次调查是对习水杜鹃短暂静态的种群判断,不建议据此便进行粗暴的人工干预,但是我们的成果形成了的一个关于习水杜鹃的“生态档案”,可以通过回访,对我们保护的目标进行长效的监测,从而更加客观的反映出种群的变化,并及时制定科学保护对策。另外,上述习水杜鹃更新因素主要侧重于对外界环境的分析,其自身种子萌发障碍情况还不清晰,建议开展一些促进习水杜鹃种子萌发的研究,并进行培育,以应对种群遭受的衰退威胁。
猜你喜欢
心声歌刊(2021年3期)2021-08-05
小聪仔(幼儿版)(2020年12期)2020-02-01
星星·散文诗(2019年15期)2019-12-29
诗潮(2019年10期)2019-11-19
星星·散文诗(2019年5期)2019-07-19
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
诗潮(2017年2期)2017-03-16
大众考古(2015年7期)2015-06-26
