
中街山列岛海域食物网结构初步研究及主要消费者潜在碳源分析
2023-01-12 09:01杨凡印瑞范江涛蒋日进肖祎王静徐汉祥刘明智
海洋学报 2023年1期
杨凡 ,印瑞,范江涛,蒋日进*,肖祎 ,王静 ,徐汉祥 ,刘明智
( 1. 浙江海洋大学 海洋与渔业研究所,浙江 舟山 316021;2. 浙江省海洋水产研究所 农业农村部重点渔场渔业资源科学观测实验站 浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江 舟山 316021;3. 中国水产科学研究院南海水产研究所,广东 广州 510300)
1 引言
食物网结构研究是海洋生态学研究的一个重要领域,海洋生物的营养结构、营养级和食源分析等方面是其重要组成部分[1-2]。探究海洋生态系统中生物的食源特征和营养关系,建立基础食物网结构,可以更好地了解复杂生态系统中的物质循环和能量流动规律[3]。在复杂的海洋生态系统中,悬浮颗粒有机物(POM)、沉积物有机物(SOM)、浮游植物、大型海藻、海草和陆源性有机质等是鱼类、大型无脊椎动物和浮游动物的重要碳源和能量来源[4],碳源又是各类生物重要的营养物质[5],但由于生态系统的复杂性,不同碳源对食物网的贡献程度各不相同且难以确定[6]。研究食物网能量来源的传统方法是胃含物分析法,由于此方法具有瞬时性,不能代表长时间的摄食情况,因此结果存在一定的误差[7]。近年来,稳定同位素技术不断发展,碳、氮稳定同位素也在食物网结构研究中得到广泛应用,其中,δ13C与饵料生物相近,可以指示食物来源的信息[8],δ15N在相邻营养级之间存在3‰~4‰的富集效应,可用于估算生物体的营养级[9]。因此,稳定同位素可以确定食物来源及食物贡献,更加准确的表征食源贡献和食物网中不同生物的营养位置[10]。此外在碳、氮稳定同位素比值基础上发展来的一系列多变量统计指标为比较不同群落的营养生态位宽度、查明营养关系、探明营养结构提供了重要方法[11-12]。
中街山列岛海域地处舟山群岛的东部,属于浙江北部近岸海域,该海域受众多水流交汇的影响,营养物质大量聚集,饵料生物丰富,成为多种海洋生物生长繁殖的重要栖息场所,历来有“东海鱼仓”之美誉[13]。由于其具有重要的生态价值,早在2006年就在此设立了国家级海洋特别保护区、国家级海洋牧场示范区[14]。目前关于该海域的研究主要集中于资源状况、岛礁生境群落结构等方面[15-17],有关该海域营养结构或食物网结构的系统研究尚未报道,而阐明中街山列岛海域营养结构对海洋牧场的建设至关重要。因此,本研究基于碳、氮稳定同位素技术初步建立中街山列岛海域食物网,分析主要消费者及潜在碳源的碳、氮稳定同位素特征,揭示不同消费者的潜在碳源,分析营养级层次并探明了食物网的连续营养谱,计算营养结构特征参数,旨在为该生态系统的可持续发展和管理提供科学依据,并为了解中街山列岛国家级海洋牧场的食物网提供基础资料。
2 材料与方法
2.1 样品采集与处理
于2020年7月在中街山列岛海域共布设10个站点进行生物样品采集(图1)。鱼类、虾蟹类和贝螺类的采集主要使用三重流刺网和底拖网采集。流刺网网长15 m,内网目直径为26 mm,外网目直径为210 mm,每个调查站位放置两组刺网,放置时间为24 h。底拖网为有翼单囊贴底拖网(网口周长50 m,网身长48 m,网目直径24 mm),拖网时间30 min,拖速3 kn。浮游动物使用浅水2型浮游生物网在表层海水中水平拖曳(拖网时间约10 min)采集;大型海藻主要由阿氏拖网采集,-20℃保存;浮游植物使用标准的小型生物网从水底定点垂直拖网至水面采集;悬浮颗粒有机物使用采水器在距离水面0.5 m处采集15 L海水来获得;使用抓斗式采泥器垂直采集表层沉积物(1~2 cm)样品,于-20℃冷冻保存。所有样品带回实验室作进一步分析。

图1 中街山列岛海域采样站位Fig. 1 Sampling stations in Zhongjieshan Islands sea area
在实验室中,对鱼、虾蟹等消费者进行种类鉴定。鱼类取背部肌肉,虾类取腹部肌肉,蟹类取螯足部位,头足类取胴体肌肉,贝类取闭壳肌。浮游动物在海水中空腹处理24 h后,经过80目的筛绢过滤,在显微镜下挑出杂质后用10% HCl酸化处理以消除碳酸盐的影响[18];大型海藻用蒸馏水清洗干净后,将其置于洗净的干燥器中,用1 mol/L的盐酸蒸汽熏蒸24 h;浮游植物样品的处理首先在显微镜下尽量挑出其中的浮游动物和杂质,用蒸馏水静置,取上清液通过泵系统抽滤至经马弗炉450℃灼烧6 h的玻璃纤维滤膜上(GF/F);POM样品先通过80目筛绢过滤,滤液抽滤至玻璃纤维滤膜上,滤膜样品(浮游植物、POM)同样采取盐酸熏蒸的方法;先取适量SOM样品干燥,磨成粉末,经过孔径为63 μm的筛绢分筛,用1 mol/L的盐酸酸化,充分反应后用去离子水洗涤[19]。所有样品在待测前均置于-20℃冰箱冷冻保存。
2.2 稳定同位素分析
把上述预处理的样品置于Alpha 1~2 L Dplus冷冻干燥机中干燥24 h,并研磨成粉末状。通过DELTA V Advantage同位素质谱仪(Isotope Ratio Mass Spectrometer)测定稳定碳、氮同位素比值。质谱仪检测样品高温燃烧后生成CO2的13C/12C比率和N2的15N/14N比率,并分别与国际标准物箭石和空气中的氮气对比后计算出样品的δ13C值、δ15N值。计算方法为
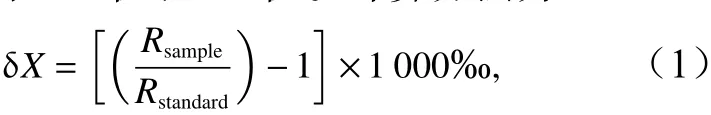
式 中,δX为δ13C值 或δ15N值,δ13C值 的 测 定 精 度 小于±0.1‰,δ15N值的测定精度小于±0.2‰;Rsample为所测样品的同位素比值;Rstandard为标准物质的同位素比值,R=13C/12C或15N/14N。
2.3 数据处理
根据生物的δ15N值计算营养级以体现其在食物链和食物网中的营养位置,其计算公式如下所示[20]:
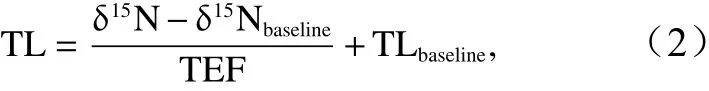
式中,TL表示 营 养 级;δ15N为生物 样 品 的δ15N值;δ15Nbaseline为基线生物的δ15N值,本研究选择浮游动物为基线生物,其δ15N平均值为6.37‰;TEF为相邻营养级间的营养富集程度,本研究为3.4‰[21];TLbaseline为基线生物的营养级,此处为2。
使用R语言(4.0.2)中的“SIBER”模型计算Layman等[11]提出的群落营养结构范围指标,包括以下6个参数:δ13C差值(CR,摄食来源多样性水平)、δ15N差值(NR,营养级长度)、同位素坐标点形成的多边形面积(TA,生物对生态空间的利用情况)、平均离心距离(CD,群落的营养多样性)、平均最近相邻距离(MNND,群落的营养相似性)和最近相邻距离的标准差(SDNND,群落的营养均匀度)。MNND和SDNND可以表示营养结构的冗余程度。R语言中的“SIAR”模型可以估计多个碳源对不同消费者的贡献率[22]。利用稳定同位素(δ13C和δ15N)和siarmcmcdirichletv4命令估算中街山列岛海域食物网中不同碳源对消费者类群的贡献率,模型中碳、氮稳定同位素营养富集因子(TEF)参考文献[21]中的富集因子,δ13C值、δ15N值分别为(0.4±1.3)‰和(3.4±1.0)‰。
通过ArcGIS 10.2软件绘制采样站位图;使用Excel 2016软件处理数据及绘制表格;通过SPSS 24.0软件进行单因素方差分析;碳源贡献率及连续营养谱的绘制运用Origin 2022软件。
3 结果
3.1 碳、氮稳定同位素特征
3.1.1 潜在碳源稳定同位素特征
中街山列岛海域共采集潜在碳源样本44个(表1)。碳源的δ13C值范围在-22.93‰~-9.73‰之间,分布范围较广,平均值为(-17.31±2.60)‰,从高到低依次为SOM、大型海藻、POM和浮游植物;δ15N值范围在1.72‰~7.68‰之间,平均值为(4.86±1.46)‰,从高到低依次为POM、SOM、大型海藻和浮游植物。分析结果显示,4种碳源之间的碳、氮稳定同位素具有显著性差异(p<0.01)。

表1 潜在碳源的δ13C值和δ15N值Table 1 Values of δ13C and δ15N for the potential carbon sources
3.1.2 消费者稳定同位素特征
共采集到49种消费者,以鱼类和虾蟹类居多,分别为24种和18种;头足类只有1种,为曼氏无针乌贼(Sepiella maindroni);贝螺类共有6种(表A1)。把消费者分为鱼类、虾蟹类、头足类、贝螺类和浮游动物,主要消费者类群的δ13C值范围在-21.91‰~-12.55‰之间,平均值为(-16.38±1.30)‰,从高到低依次为虾蟹类、头足类、鱼类、贝螺类和浮游动物;δ15N值范围在4.13‰~12.92‰之间,平均值为(8.93±1.71)‰,从高到低依次为头足类、鱼类、虾蟹类、浮游动物和贝螺类(表2)。不同类群消费者之间的碳、氮稳定同位素均有显著性差异(p<0.01)。

表2 主要消费类群的δ13C值和δ15N值Table 2 Values of δ13C and δ15N for the major consumer groups
结合消费者和潜在碳源δ13C值、δ15N值的双位图(图2)来看,潜在碳源的δ13C值分布范围较广,基本涵盖了消费者的δ13C范围,而δ15N值呈现明显阶层分布,鱼类、虾蟹类和头足类δ15N值较大,分布在上层,而贝螺类和浮游动物δ15N值较小,分布在下层。
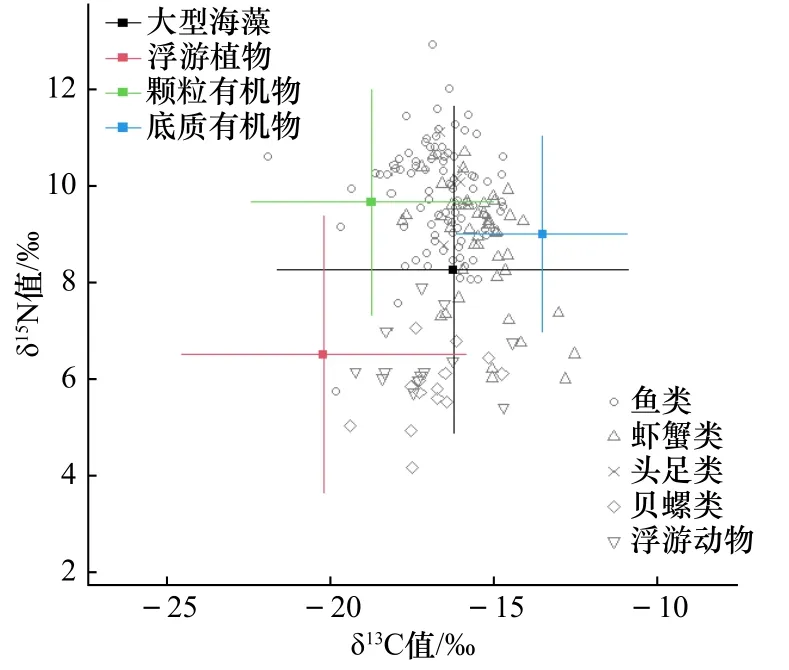
图2 中街山列岛海域主要消费者和潜在碳源的δ13C和δ15N值双位图Fig. 2 Stable isotope biplots of δ13C and δ15N values of major consumers and the potential carbon sources in Zhongjieshan Islands sea area
3.2 营养结构指标
基于SIBER模型计算除浮游动物以外的消费者稳定同位素的营养结构指标[23],结果如表3所示。中街山列岛海域主要消费者的 δ13C差值(CR)、δ15N差值(NR)、多边形总面积(TA)和平均离心距离(CD)均大于其他海域,表明该海域生物摄食来源广泛,对空间的利用程度高,群落的营养多样性也较高。平均最近相邻距离(MNND)和最近相邻距离标准差(SDNND)较高,与海州湾的结果较为相似,表明营养冗余程度较低。
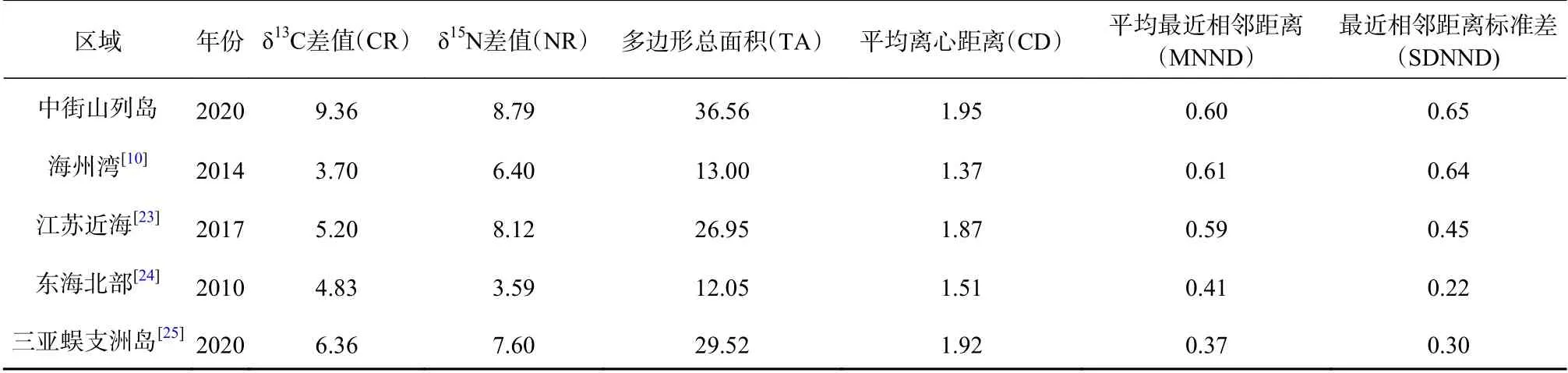
表3 不同区域主要消费者的营养结构指标Table 3 Trophic structure indicators of major consumers in different regions
3.3 不同碳源对消费者的贡献
基于4种潜在碳源的SIAR模型计算结果表明,大型海藻对所有生物的碳源贡献范围为0~46.94%,其中对虾蟹类的贡献最大,平均值为26.58%;浮游植物的贡献范围变化较大,对所有生物的碳源贡献范围为0~80.58%,对浮游动物的贡献最大,平均值为65.15%;POM的贡献范围在0~79.74%之间,其中对鱼类的贡献最大,平均值为70.19%;SOM对所有生物的碳源贡献范围为0~49.94%,其中对虾蟹类的贡献最大,平均值为35.09%(图3)。综合来看,浮游植物和POM对中街山列岛海域消费者的碳源贡献最高,平均值约为29.63%和28.72%;鱼类和头足类的主要碳源为POM和SOM;虾蟹类主要碳源为大型海藻、POM和SOM;贝螺类和浮游动物的主要碳源为浮游植物。
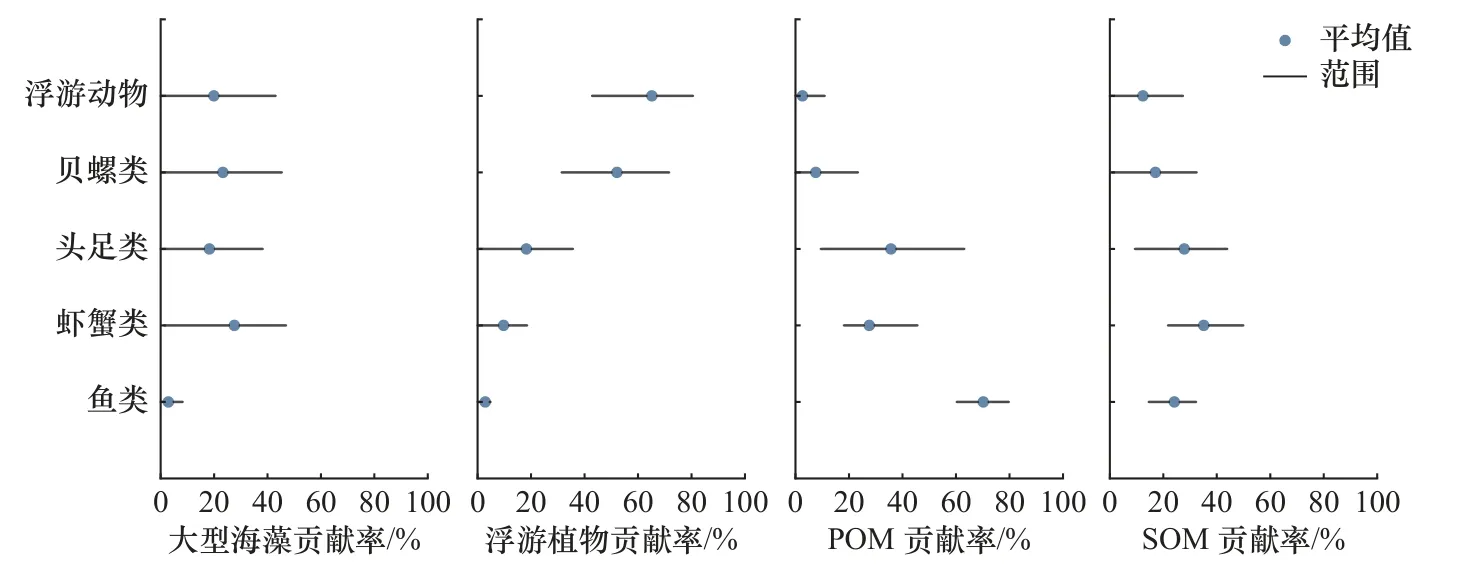
图3 潜在碳源对主要消费类群的贡献率Fig. 3 Contribution of potential carbon sources to major consumer groups
3.4 营养级分析
以浮游动物为基线生物,计算中街山列岛海域主要消费者的营养级并绘制连续营养谱。由图4 可知,主要消费者的平均营养级在1.58~3.63之间,均值为2.82±0.47,属于3个营养层次。其中鮸鱼(Miichthys miiuy)的营养级最高,紫贻贝(Mytilus edulis)的营养级最低。各类群的营养级差异较大,鱼类的营养级在2.59~3.63之间,平均值为3.00±0.32;虾蟹类营养级在1.88~3.01之间,平均值为2.70±0.36;头足类营养级在2.69~3.39之间,平均值为3.09±0.25;贝螺类的营养级在1.34~2.19之间,平均值为1.82±0.22。
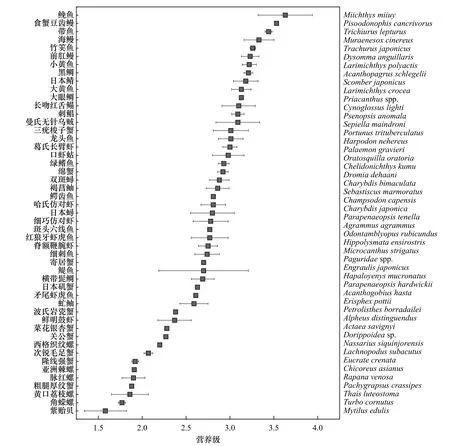
图4 中街山列岛海域主要消费者连续营养谱Fig. 4 Continuous nutritional profile of major consumers in Zhongjieshan Islands sea area
由图4可知鱼类和虾蟹类的营养级跨度较大,为了更准确地计算潜在碳源对鱼类和虾蟹类的贡献,根据国内学者对于营养层次的划分标准[26],按其营养级细分为低营养级鱼类(2~2.8)、中营养级鱼类(2.9~3.4)和高营养级鱼类(3.5~4),以及低、中营养级虾蟹类,计算4种潜在碳源对其贡献。由图5可知,大型海藻对不同营养层次的鱼类、虾蟹类的碳源贡献范围为0~48.93%,其中对低营养级鱼类、虾蟹类的贡献较大,平均值分别为26.37%和24.19%;浮游植物对不同营养层次的鱼类、虾蟹类的碳源贡献范围为0~30.77%,对高营养级鱼类的贡献最大,平均值为14.32%;POM对不同营养层次的鱼类、虾蟹类的碳源贡献范围为0~68.13%,其中对中营养级鱼类的贡献最大,平均值为60.71%;SOM对不同营养层次的鱼类、虾蟹类的碳源贡献范围为0~68.36%,其中对中营养级虾蟹类的贡献最大,平均值为50.29%。综合来看,不同营养层次鱼、虾蟹类主要潜在碳源为POM和SOM,此外大型海藻对低营养级鱼类、虾蟹类贡献也较大,这与图3所示结果较为相符,即鱼类主要碳源为POM和SOM,虾蟹类的主要碳源为大型海藻、POM和SOM。
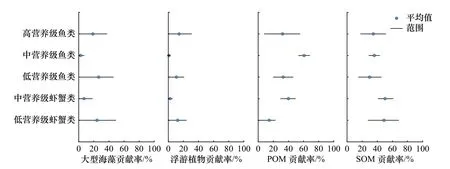
图5 潜在碳源对不同营养级鱼类和虾蟹类的贡献率Fig. 5 Contribution of potential carbon sources to different trophic levels of fishes and shrimps and crabs
4 讨论
4.1 中街山列岛海域潜在碳源及主要消费者的碳、氮稳定同位素特征
有关近海生态系统食物网的研究表明,浮游植物、大型藻类、POM和SOM 是海洋消费者的潜在碳源,也是驱动食物网碳循环的主要动力[4]。本文结果显示,中街山列岛海域潜在碳源主要由大型海藻、浮游植物、POM和SOM组成,这些碳源的δ13C值跨度较大(-22.93‰~-9.73‰),δ15N值跨度较小(1.72‰~7.68‰)。碳源稳定同位素值通常受水域环境、有机质来源等多种因素影响[22]。大型海藻的δ13C值在不同季节存在较大差异,随着气温升高,大型海藻利用CO2的能力下降,δ13C值升高[27]。本次采样时间在夏季,水体温度较高,因而导致大型海藻的δ13C值较高;浮游植物作为初级生产者,具有较低的δ13C值和δ15N值,其中δ13C组成主要来自海洋水体中的溶解无机碳(DIC),本研究浮游植物的δ13C值范围与DIC的δ13C值范围(-22‰~-18‰)也基本吻合[28];POM主要包括浮游植物、浮游动物、无机颗粒等[29],POM的δ13C值、δ15N值往往与浮游植物等的δ13C值、δ15N值具有相关性[28];SOM的来源包括藻体凋落物、动物腐尸、粪便、底泥以及来自陆地的POM输入等[30],有研究表明,大型海藻处于7月凋亡期时对SOM的贡献最大[31],而本研究SOM的δ13C值与大型海藻的δ13C值更为接近,这有可能说明大型海藻对SOM有较大贡献。
碳稳定同位素在食物网中富集较低,可用于示踪食物来源[8]。本研究中不同类群消费者之间的δ13C值存在显著性差异(p<0.01),其中浮游动物和虾蟹类的δ13C最大差值为6.66‰。研究表明栖息水层影响生物获取不同的能量来源,由此表现出不同的δ13C值特征,由于底栖生物具有较强的碳同位素富集能力,因此碳同位素比值高的生物与底栖食物网有关,低的与浮游食物网有关[32],这也说明中街山列岛海域存在底栖食物链与浮游食物链。中街山列岛位于浙江北部,高春霞等[33]调查过浙江南部近海主要渔业生物的碳、氮稳定同位素比值,其中δ13C值范围为-19.71‰~-14.01‰、δ15N值范围为7.05‰~13.69‰。本研究与邻近的浙江南部海域结果相近,但生物δ13C最大值要更大,因为初级生产者的δ13C值随离岸距离的减小而增大,δ13C值通过食物链传递给消费者,所以生活在近岸的消费者会有较高的δ13C值[34]。中街山列岛整体属于近岸水域,水域范围较小,因此消费者的δ13C最大值会比浙江南部近海的偏大,并且近岸水域更容易受到人类活动的影响,陆源物质如动植物碎屑等的输入使得该海域的基础食源更加丰富,进而影响消费者的δ13C值。较高的日照水平会引起初级生产者的碳富集[35],随着纬度的升高,水域食物网中最负δ13C值会出现贫化趋势,因此本研究与浙江南部的最负δ13C值相比稍贫化。本研究δ15N值与浙江南部相比较低,这可能与样品差异有关,本研究中鱼类的δ15N值在5.71‰~12.92‰之间,最大值要小于浙江南部六指马鲅(Polydactylus sextarius)、黄鮟鱇(Lophius litulon)等高级消费者鱼类的δ15N值(最大值约13.5‰);贝螺类的δ15N值在4.13‰~7.03‰之间,同样远低于浙江南部双壳类和腹足类的δ15N值,除采样区域、样品种类和大小规格的差异,沉积物和水体中氮的含量也会影响生物的δ15N值,如氮含量相对较高,底栖生物的δ15N值也会偏大[36]。
4.2 营养结构指标特征
CR越大代表食物网中初始食源越多[11]。中街山列岛海域夏季的浮游植物丰度比其他季节要高[15],而浮游植物作为近海初始食源对消费者的贡献率较大,此外海洋牧场、贻贝筏式养殖区的建立以及部分陆源有机物的输入使得该区域食源更为丰富,因此该海域的CR值高;NR较大表示消费者占有更多的营养层次,食物链更长[11]。中街山列岛海域夏季鱼卵、仔稚鱼[13]以及游泳生物[16]的资源量要高于其他季节,丰富的资源量使得该海域的营养多样性水平较高;由于中街山列岛海域的食源丰富,营养层次多,因此TA值、MNND值和SDNND值同样较高,说明了该海域营养冗余性低的特性。海洋生物群落中饵料生物的变动会引起消费者竞争的变化,进而导致具有相似营养生态位的消费者增多或减少,影响生态系统的冗余程度[24]。冗余可以提高生态系统的可靠度,维持其结构与功能的正常运行,营养级内的冗余如物种冗余时,某些物种的丧失不会对生态系统的结构和功能产生很大的影响,但由于生态系统不同营养级内物种之间复杂的组合方式,一个物种的丧失可能会引起营养级内或营养级之间的波动[37]。由于本研究只有不同海域营养结构的对比,因此为了更好地评估海洋牧场建设对中街山列岛海域生态系统稳定性的影响,将来需要从时间尺度上综合阐述该海域的营养结构变化。
4.3 潜在碳源分析
近岸海域由于其特殊的地理位置往往具有十分明显的海陆相互作用[22]。中街山列岛位于浙江北部近岸水域,沿岸水流和内陆径流的交汇以及岛礁近岸的大型海藻和贻贝筏养殖区为该海域提供了丰富的物质来源[17]。浮游植物、POM、SOM和大型海藻对中街山列岛海域主要消费者类群的贡献率平均值依次为29.63%、28.72%、23.28%和18.37%。整体来看,浮游植物和POM是该水域的重要碳源,两者对消费者的贡献范围分别在0~80.58%和0~79.74%之间。浮游植物是海洋重要的初级生产者,是海洋食物网的重要基础,中街山列岛海域浮游植物的碳源比例最大,这与崂山湾[38]、海州湾[10]研究结果一致。相较于以往研究,中街山列岛海域浮游植物的碳源贡献略低,这可能是由于温度、海流和营养盐浓度引起的环境变化影响了浮游植物的含量。此外大型藻类会与浮游植物产生资源竞争[22],而养殖贝类也会影响浮游植物的丰度,从而降低浮游植物的生物量,减少其碳源贡献。POM对中街山列岛海域消费者的贡献率较大,因中街山列岛属于近岸水域,水体中颗粒悬浮物的含量较高,并且大型海藻降解形成的POM和陆源POM会随降雨流入,这也可能增加POM的含量,从而导致其更高的碳源贡献率[39]。SOM的贡献比例次之,尤其是对于底栖虾蟹类,平均贡献比例为35.09%。这与尹洪洋等[25]的结论相一致,即SOM对海洋底栖生物的贡献较大。大型海藻在近岸海域食物网中同样占有重要的地位,它不仅通过光合作用为食物网提供了大量的初级生产力,也因其适口性和高营养价值,为小型无脊椎动物等提供了饵料来源[35]。头足类、虾蟹类等无脊椎动物的直接或间接啃食作用可获得能量并向上传递初级生产力,因此,大型海藻对虾蟹类、头足类、贝螺类和浮游动物的贡献比例要明显大于鱼类。
4.4 营养级分析
基线生物的选取对定量研究生物体的营养位置至关重要,由于基线反映的是最初物质来源的同位素特征,因此一般选择初级生产者或初级消费者作为基线生物[40]。本研究选择浮游动物作为基线生物,以其氮稳定同位素比值的平均值(6.37‰)作为基准值计算中街山列岛海域主要消费者的营养级,结果显示,主要消费者的平均营养级范围为1.58~3.63,营养层次为3级,其中头足类的平均营养级最高(3.09),其次是鱼类(3.00)、虾蟹类(2.70)和贝螺类(1.82)。营养级的高低与消费者的食性有关,头足类会摄食鱼类,如曼氏无针乌贼具有强大的摄食器官,发达的角质颚和齿舌既能破坏鱼类的头骨,也能穿破甲壳类的外壳,因此食物来源广泛,主食龙头鱼(Harpodon nehereus)、带鱼(Trichiurus lepturus)、中华管鞭虾(Solenocera sinensis)、口虾蛄(Oratosquilla oratoria)等游泳生物,故营养级较高[41]。鱼类营养级在2.59~3.63之间,营养级最低的为虻鲉(Erisphex pottii,2.59),其次为矛尾虾虎鱼(Acanthogobius hasta,2.61)。中街山列岛海域矛尾虾虎鱼的营养级远低于东海矛尾虾虎鱼的营养级(3.24)[42],原因是本次调查采集到的矛尾虾虎鱼体长偏小,饵料也以钩虾、糠虾等小型低营养级的生物为主[43];营养级最高的为鮸鱼(3.63),由于本研究与吕泗渔场鮸鱼的样本规格较为一致,因此营养级结果也相似(吕泗渔场的为3.61)[44]。有些虾蟹类如口虾蛄、三疣梭子蟹(Portunus trituberculatus)、葛氏长臂虾(Palaemon gravieri)等营养级较高,甚至超过部分鱼类,一方面是因为SOM对底栖虾蟹类的贡献最大,SOM较高的δ15N值可以通过食物链富集在虾蟹类体内;另一方面虾蟹类的摄食类型也会随着生长发育而转变,由幼时的滤食性转变为以小型鱼类、海洋无脊椎动物为食的肉食性[3],本研究中口虾蛄等个体规格较大,因此营养级较高。贝螺类一般以海藻或碎屑为食,故营养级最小[45]。
根据国内学者对于营养层次的划分标准[26],中街山列岛海域主要消费者基本处于第二至第四营养级。其中高级肉食性动物有带鱼、食蟹豆齿鳗(Pisoodonophis cancrivorus)和鮸鱼,即营养级位于3.5~4之间,占总数的6.12%;由龙头鱼、长吻红舌鳎(Cynoglossus lighti)、曼氏无针乌贼等为代表的中级肉 食 性 动 物(2.9~3.4),和 以 鳀 鱼(Engraulis japonicus)、虻鲉、鲜明鼓虾(Alpheus distinguendus)为代表的低级肉食性动物分别占总数的42.85%和36.73%;而亚洲棘螺(Chicoreus asianus)等草食或杂食性动物占14.28%。由此可见,该海域食物网的主要消费者为低、中级肉食性动物,在两端(杂食性和高级肉食性动物)的较少。与邻近海域相比,中街山列岛海域食物网的营养层次范围接近于黄、东海(1.43~3.62)[42],小于浙江南部近海(2.66~4.21)[33]。一方面,本研究中低营养级的贝螺类较多,鱼类也以中小型鱼类为主,鲨类、鳐类和鮟鱇等大型肉食性鱼类并没有采集到,这会导致营养级偏低;另一方面生物体长会影响营养级,有些鱼类如带鱼、小黄鱼(Larimichthys polyactis)等体长往往与δ15N呈正相关趋势[42],不同研究区域,体长有差异的同一种生物营养级也会不同,中街山列岛海域的绿鳍鱼(Chelidonichthys kumu)、前肛鳗(Dysomma anguillaris)等的营养级远低于浙江南部,原因是本研究的体长规格较小。此外,基线生物的选择以及营养富集因子也会影响营养级结果。高春霞等[33]选择双壳类(5.34‰)作基线生物,而本研究选取的浮游动物(6.37‰),但浮游动物不同大小、不同种类以及季节性变化等都会对营养级估算产生影响,有一定的局限性[25]。营养富集因子本研究选择国际通用的3.4‰,而蔡德陵等[42]通过室内饲养鳀获得氮富集因子为2.5‰,高世科等[44]发现叠加潜在碳源作为基准值计算生物营养级更适用于低营养级的生物种类。因此选择合适的基准生物和富集因子更有利于评价生态系统营养结构。
5 结论
本研究表明,中街山列岛海域食物来源丰富、营养多样性水平高,营养冗余性与其他海域对比相对较低,说明该海域饵料生物较多,消费者食源竞争小,处于相似营养生态位的物种少;浮游植物和POM是中街山列岛海域的主要碳源,贡献比例平均值分别为29.63%、28.72%;主要消费者的平均营养级范围为1.58~3.63,营养层次为3级,以低、中级肉食性动物为主,杂食性和高级肉食性动物较少。海洋牧场的建设为物种的生存提供了更为有利的生境条件,是中街山列岛海域开展渔业资源养护与修复的有效手段。本研究初步探索了中街山列岛海域的食物网结构,但海洋牧场建设对食物网结构的影响是多方面、长期性的,在将来的研究中,需要结合环境因素(温度、光照、盐度等),从不同时空尺度上综合评估海洋牧场建设对该海域食物网结构以及整个生态系统的影响,也能为近海渔业资源修复等问题提供一定的理论参考。

表 A1 中街山列岛海域主要消费者的体长信息和δ13C、δ15N值Table A1 The body length and δ13C, δ15N values of major consumers in Zhongjieshan Islands sea area
附录
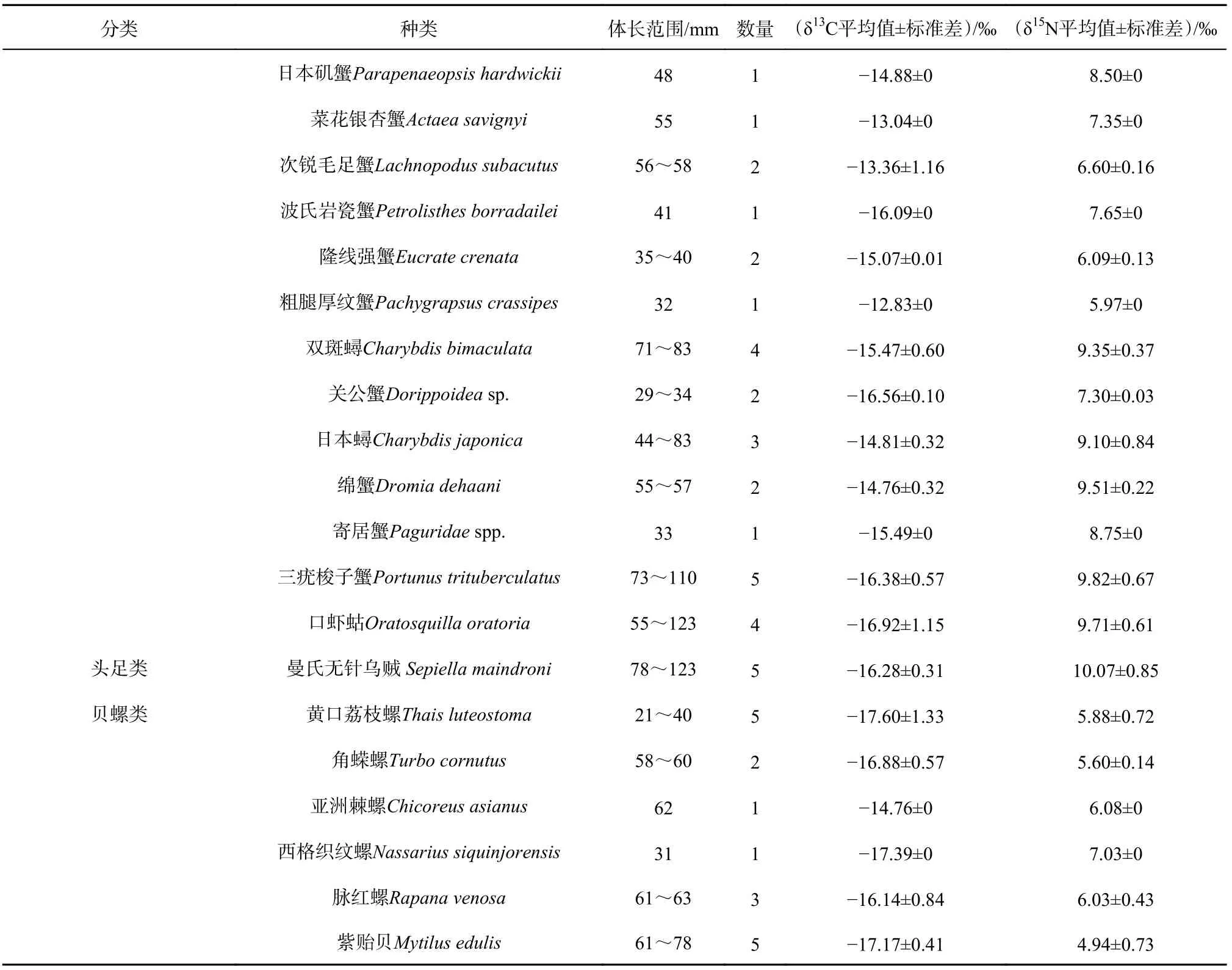
续表 A1
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
天津科技(2020年4期)2020-05-09
同位素(2018年1期)2018-01-18
东南文化(2016年4期)2016-08-22
中学生天地(A版)(2015年7期)2015-08-06
同位素(2014年3期)2014-06-13
同位素(2014年2期)2014-04-16
同位素(2014年2期)2014-04-16
