
长山列岛邻近海域鱼类群落功能多样性的季节和空间变化
2023-01-12 09:01邹建宇刘淑德张崇良薛莹纪毓鹏徐宾铎
海洋学报 2023年1期
邹建宇,刘淑德,张崇良, ,薛莹, ,纪毓鹏, ,徐宾铎, *
( 1. 中国海洋大学 水产学院,山东 青岛 266003;2. 青岛海洋科学与技术试点国家实验室 海洋渔业科学与食物产出过程功能实验室,山东 青岛266237;3. 海州湾渔业生态系统教育部野外科学观测研究站,山东 青岛 266003;4. 山东省渔业发展和资源养护总站,山东 烟台 264003)
1 引言
功能性状指生物体在长期的环境胁迫、历史进化和自然选择的过程中形成的定量或定性的显著本质特征[1],包括生态学性状和生物学性状。生态学性状主要与栖息地、迁移等相关[2],生物学性状主要与生理行为、生活史周期等相关[3]。功能多样性即功能性状多样性[4],指影响生态系统功能的物种性状的数值、范围和分布[5]。
近些年来,α多样性[6-8]、β多样性[9-10]和分类学多样性[11-13]在鱼类群落多样性研究中得到了广泛应用,但传统的多样性分析仅考虑了物种相对生物量信息,将拥有不同形态特征和生活习性的物种看作等价的,未能很好地反映生物对环境的响应[14],功能多样性以长期自然选择的性状为基础,能更好地理解生物与环境之间的关系。鱼类的功能性状主要指其在生理行为、生态适应、种群动态等方面的特征,如适温性、食性、洄游类型、繁殖行为、口的位置、眼间距、鳍的形状、体型等[15]。目前国内外鱼类群落功能多样性的研究主要涉及功能性状的选择[16-17]、功能多样性的时空变化[18]和功能多样性与环境因子的相互作用[19],其研究区域涵盖河流[20-21]、湖泊[22]、海洋[23-25]等。
长山列岛邻近海域地处黄海、渤海生态交错带,位于莱州湾渔场和烟威渔场交界处,地理位置独特,水文条件复杂,捕捞压力较大。由于独特的地理位置,黄海水和渤海南部沿岸水在此处交汇[26],在不同季节受到季风的影响,两大水团和海洋环流发生剧烈变化[27],因此该海域环境因子季节变化显著。夏季黄海冷水团[28]分布范围可达该海域东北部,造成该海域夏季环境因子的空间差异较大;冬季黄海暖流[29]入侵该海域东部,使得该海域冬季环境因子的空间差异较大。此外,海岛与海水的镶嵌式格局[30]影响着海流流向,使得该海域展现出了独特的水文特点。在复杂环境因子的作用下,该海域鱼类群落功能性状及功能多样性是否具有显著的时空差异值得研究,该研究对深入理解长山列岛邻近海域群落结构和功能具有重要意义。
本文根据2016-2017年在长山列岛邻近海域进行的鱼类资源季度调查数据,应用群落特征加权平均数指数、功能多样性指数和Spearman秩相关分析等方法,研究该海域鱼类群落功能多样性的时空格局及其与环境因子的关系,以期为该海域鱼类资源的管理、开发和保护提供参考,同时为未来该海域鱼类群落功能多样性不同年间比较或长期变化研究提供参考。
2 研究区域与研究方法
2.1 调查海区与数据来源
调查海区为长山列岛邻近海域(37°30′~38°30′N,119°50′~121°50′E),共设置42个采样站位(图1)。由于海况条件等限制性因素,春、夏、秋、冬季实际拖网站位数分别为36个、39个、40个、38个[31]。调查船只为功率220 kW的底拖网船,网具网口高度约7.53 m,宽度约15 m,囊网网目大小为17 mm。拖网均在白天进行,预设每站拖网时间1 h,拖速3 kn。水温、水深、盐度等环境因子使用CTD(型号为CTD75M/1167)调查获得。参考《海洋调查规范 第6部分:海洋生物调查》(GB/T 12763.6-2007)[32]对渔获物取样并带回实验室进行分析处理,并将原始调查数据进行标准化处理为拖网1 h、拖速3 kn时的单位网次渔获重量(单位:kg/h)。
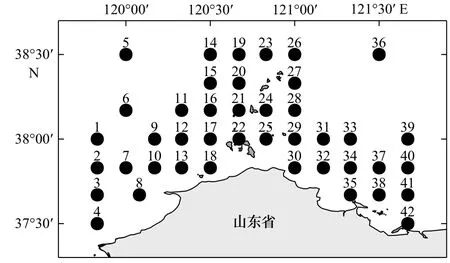
图1 长山列岛邻近海域渔业资源底拖网调查站位Fig. 1 Bottom trawl survey stations for fishery resources in the adjacent waters of the Changshan Islands
2.2 功能性状选择
根据以往对鱼类群落功能多样性的研究[18,25],本研究选取反映鱼类摄食、游泳、生态适应、群落动态和繁殖5种功能类型的性状,其中包括食性、口的大小、口的位置、营养级、体型、洄游类型、最大体长、适温性、栖息位置、生长系数、恢复力、脆弱性和鱼卵类型。各性状指标在分析中所占权重相等,营养级、最大体长、生长系数和脆弱性为连续变量,其余变量为分类变量(表1)。通过查阅世界鱼类数据库(Fish-Base)[33]和相关文献[34-35,18,25]等获得各指标的取值或类型,功能性状取值或类型的确定遵循邻近优先原则,即长山列岛邻近海域相关资料缺失时以黄、渤海等相邻海域替代。各鱼种具体功能性状详见附表A1。

表1 功能性状分类标准及其所含类型Table 1 Classification criteria and types of functional traits
2.3 分析方法
应用群落特征加权平均数指数(Community Weighted Mean, CWM)分析鱼类群落优势功能性状的季节变化,群落特征加权平均数指数为群落内物种功能性状的加权平均值[36],公式为

式中,S为物种数;pi为物种i的相对渔获重量;traiti为物种i的性状值。
功能丰富度指数(Functional Richness Index, FRic)用功能体积指数表示,由功能性状空间内形成的最小凸多边形体积来计算[16]。
功能均匀度指数(Functional Evenness Index,FEve)用多维功能均匀度指数表示,推导公式[16]为
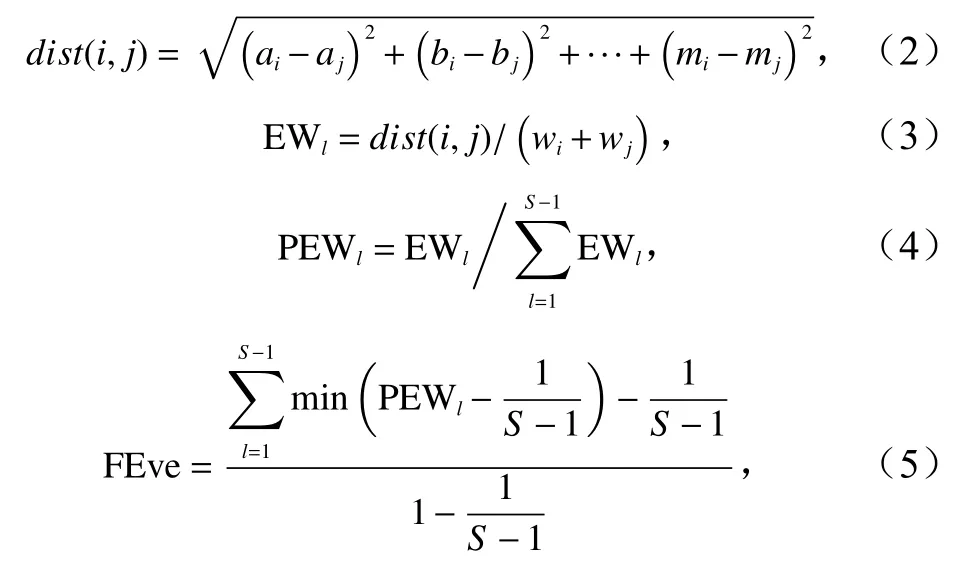
式中,S为物种数;a-m表示多维性状空间中物种i和j的13个功能性状;dist(i,j)表示物种i和j之间的欧氏距离;EWl为第l分支长,即物种i与物种j构成的分支长;wi和wj分别为物种i和物种j的相对渔获重量;PEWl为第l分支长权重。
功能离散度指数(Functional Divergence Index,FDiv)用多维功能离散度指数表示,推导公式为[16]

式中,S为物种数;xik为物种i性状k的值;gk为性状k的重心;T为性状数;dGi为物种i与重心的距离;为dGi的平均值;Δd为以丰度为权重的离散度;Δ|d|为Δd的绝对值;wi为物种i的相对渔获重量。
由于环境变量与功能多样性指数变化复杂,本研究采用Spearman秩相关系数进行相关性分析[37]。
2.4 数据处理
为减小调查采样对功能多样性指数计算的影响,在调查站位数不变的情况下,对原始调查数据进行有放回的随机再抽样99次,获得各调查航次鱼类相对资源量的99组模拟数据,计算得到各功能多样性指数的99个模拟值。应用R语言的FD程序包计算功能多样性指数,应用单因素方差分析和多重比较(α=0.05)进行季节间的差异显著性检验,采用R语言的psych程序包计算功能多样性指数与各环境因子的Spearman相关系数并进行显著性检验。采用R语言的ggplot2程序包和surfer13.0软件进行作图。
3 结果
3.1 优势性状的季节变化
采用群落特征加权平均数指数分析长山列岛邻近海域鱼类群落优势性状的季节变化趋势(图2)。从营养级来看,春、冬季鱼类群落的营养级明显高于夏、秋季。在鱼类的摄食方面,春、夏季优势性状以游泳动物食性为主,冬季以游泳动物食性和底栖生物食性为主,秋季出现了3种食性类型。从口的位置来看,春、冬季优势性状以端位口和上位口为主,夏、秋季以端位口为主。口的大小方面,春季优势性状仅表现为口裂大,夏、秋、冬季均表现为3种口裂大小。从最大体长看,春、冬季优势性状最大体长较大,夏、秋季优势性状最大体长较小。体型方面,春、冬季优势性状表现为平扁形,夏季优势性状表现为侧扁形和纺锤形,秋季优势性状表现为侧扁形、鳗形和平扁形。洄游类型方面,春季优势性状以定居型和近海洄游型两种,夏季优势性状出现外海洄游型,秋季优势性状涵盖了3种洄游类型,冬季优势性状均为近海洄游型。适温性方面,春、冬季优势性状均为暖温性,夏、秋季优势性状为暖温性和暖水性。生长系数方面,夏季生长系数最高,其次为秋季,春、冬季生长系数显著低于夏、秋季。从栖息位置来看,春、秋、冬季优势性状均为底层栖息,夏季优势性状为底层和中上层栖息。恢复力方面,春季优势性状表现出较低的恢复力,夏季表现为中等恢复力,秋季较低、中等、较高均有,冬季表现为较低和中等恢复力。从鱼卵类型来看,春季为黏着浮性卵,夏、秋季均为浮性卵。脆弱性方面,春、冬季明显高于夏、秋季,夏季脆弱性最低。
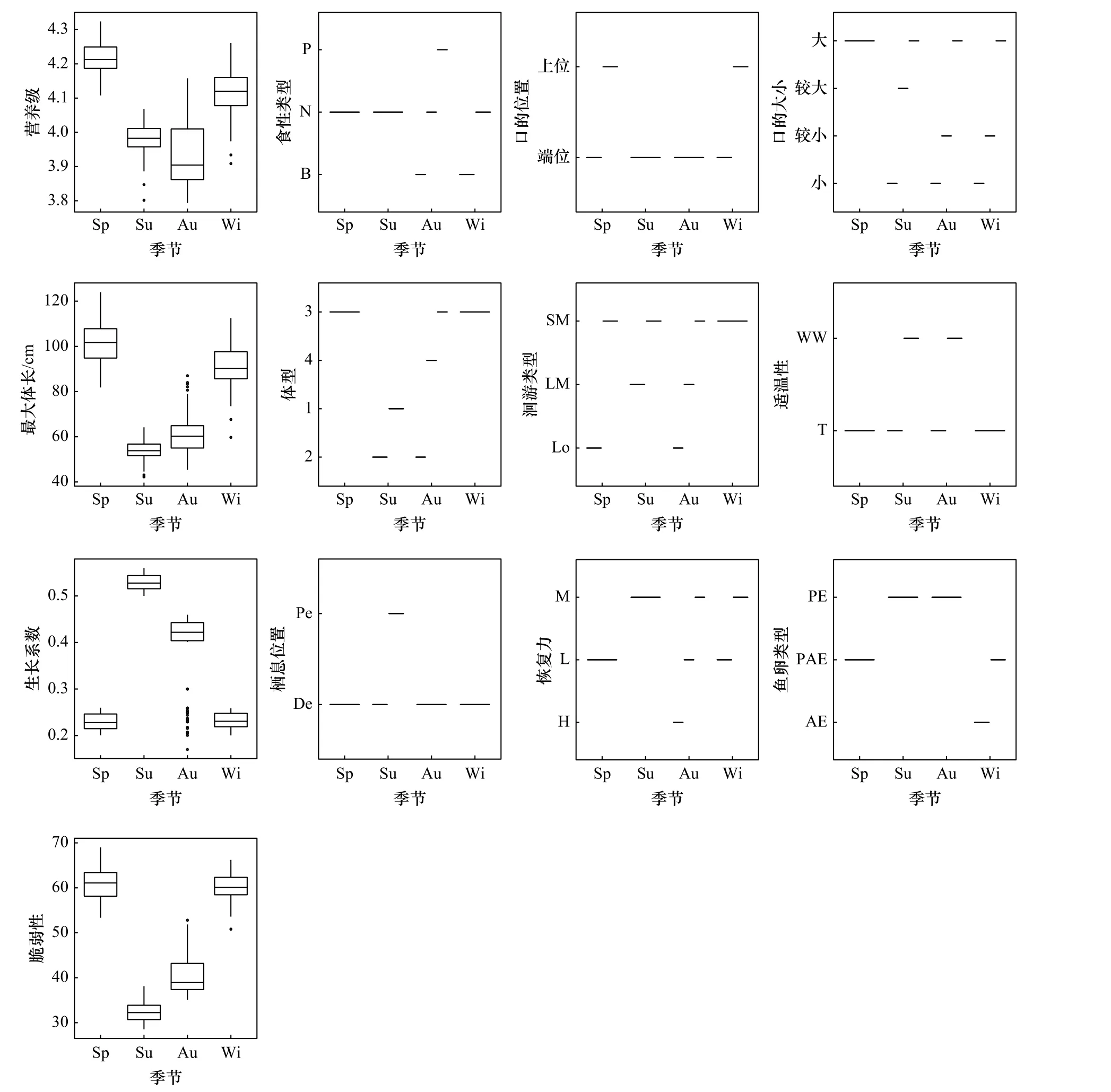
图2 长山列岛邻近海域鱼类群落特征加权平均数指数(CWM)的季节变化Fig. 2 Seasonal variation of community weighted mean index (CWM) of fish community in the adjacent waters of the Changshan Islands
3.2 功能多样性的季节变化
长山列岛邻近海域鱼类群落各功能多样性指数呈现一定季节变化(图3)。单因素方差分析和多重比较表明,功能丰富度指数在两两季节间存在显著差异(p<0.05)。秋季最高,为0.76,模拟值的变化范围为0.69~0.87;其次为夏季,功能丰富度指数为0.62,模拟值的变化范围为0.59~0.73;再次为春季;冬季最低。功能均匀度指数在夏季和冬季间无显著性差异,其余两两季节间差异性显著(p<0.05)。功能均匀度指数春季最高,为0.51,模拟值的变化范围为0.29~0.60;其次为秋季,为0.39,模拟值的变化范围为0.20~0.54;夏季与冬季均表现出较低的功能均匀度指数。功能离散度指数在春季、夏季和冬季两两季节间均无显著性差异,而秋季与春、夏、冬季间存在显著性差异(p<0.05)。秋季功能离散度指数最低,且显著低于其他季节,为0.83,模拟值的变化范围为0.7~0.87。
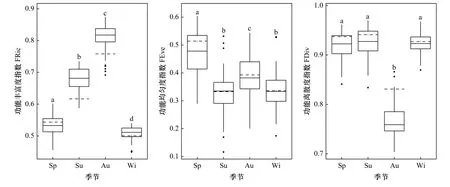
图3 长山列岛邻近海域鱼类群落功能多样性指数的季节变化Fig. 3 Seasonal variation of functional diversity indices of fish community in the adjacent waters of the Changshan Islands
3.3 功能多样性的空间变化
长山列岛邻近水域鱼类群落各功能多样性指数存在一定空间差异(图4)。春季各站位功能丰富度指数的变化范围为0.07~0.89,长山列岛中部海域及东南部明显高于其他海域,最低值出现在长山列岛东北部;功能均匀度指数的变化范围为0.09~0.80,总体表现西高东低,最高值出现在西南部海域;功能离散度指数的变化范围为0.59~0.99,最高值出现在西北部海域。夏季各站位功能丰富度指数的变化范围为0.08~0.83,总体表现东高西低;功能均匀度指数的变化范围为0.14~0.90,分布并无显著规律,最低值出现在中部偏东海域;功能离散度指数的变化范围为0.59~0.98,总体表现东高西低。秋季各站位功能丰富度指数的变化范围为0.16~0.88,大部分海域指数较低,仅在中部偏北海域较高;功能均匀度指数的变化范围为0.07~0.88,大部分海域指数较高,最低值出现在西南角海域;功能离散度指数的变化范围为0.31~0.96,总体表现西高东低。冬季各站位功能丰富度指数的变化范围为0.11~0.81,总体表现东高西低;功能均匀度指数的变化范围为0.10~0.78,总体表现西高东低;功能离散度指数的变化范围为0.59~0.99,大部分海域指数处于较高水平,仅在中部偏南海域较低。

图4 长山列岛邻近海域鱼类群落功能多样性指数的空间分布Fig. 4 Spatial distribution of functional diversity indices of fish community in the adjacent waters of the Changshan Islands
3.4 功能多样性与环境因子的相关性
Spearman秩相关分析表明,长山列岛邻近海域鱼类群落各功能多样性与水深、温度和盐度之间存在一定的相关性(表2)。春季功能均匀度指数与底层水温存在显著的正相关关系(p<0.05)。夏季功能丰富度指数与底层水温存在极显著的负相关关系(p<0.01)。秋季各功能多样性指数与环境因子之间无显著相关性。冬季功能丰富度指数与水深、底层水温、表层水温、底层盐度和表层盐度均存在极显著的正相关关系(p<0.01);功能均匀度指数与底层水温存在极显著的负相关关系(p<0.01),与表层水温均存在显著的负相关关系(p<0.05)。

表2 长山列岛邻近海域鱼类群落功能多样性指数与环境因子的Spearman秩相关系数Table 2 Spearman rank correlation coefficients between functional diversity indices and environmental factors in the adjacent waters of the Changshan Islands
4 讨论
群落特征加权平均数指数反映了鱼类对于环境因子的响应和适应机制[38]。由于环境因子的选择作用,不同季节的优势性状具有显著差异,表明不同季节间鱼类生态位分化明显[39]。从摄食来看,春、冬季的优势性状表现为口裂较大,上位口和端位口并存,游泳动物食性,这类优势性状的代表物种为黄鮟鱇(Lophiuslitulon)。黄鮟鱇主要摄食小黄鱼(Larimichthys polyactis)、矛尾虾虎鱼(Chaeturichthys stigmatias)、带鱼(Trichiurus lepturus)等游泳生物,是凶猛肉食性鱼类[40],黄鮟鱇“最大体长”的功能性状值高,因此春季、冬季在营养级和最大体长方面的优势性状明显高于夏、秋季,且在体型上优势性状表现为平扁形,在栖息类型上表现为底栖。春、夏季是鱼类重要的产卵洄游时期,黄鲫(Setipinna tenuifilis)、赤鼻棱鳀(Thryssa kammalensis)等鱼类到长山列岛邻近海域浅水区产卵[34],因此优势性状表现为近海洄游型。夏、秋季是鱼类索饵洄游和越冬洄游的重要时期,夏、秋季的渔获重量占比最高的物种为鲐(Scomber japonicus)和小眼绿鳍鱼(Chelidonichthys spinosus),鲐是纺锤形、远海洄游的中上层暖温性鱼类[41],因此在夏、秋季出现了远海洄游型、纺锤形体型和中上层栖息的优势性状。长山列岛邻近海域海水温度的季节性变化较大,优势种从黄鮟鱇、矛尾虾虎鱼、赤鼻棱鳀、鳀(Engraulis japonicus)等暖温种到鲐、小眼绿鳍鱼等暖水种更迭,因此在夏、秋季适温性方面优势性状表现为暖水性。夏、秋季种类的生长系数、恢复力均明显高于春、冬季,而脆弱性明显低于春、冬季,这表明夏、秋季鱼类群落受到干扰后更容易恢复,优势鱼类往往是生活史周期较短、小型成体、快速发育的低龄繁殖鱼类[42],例如方氏云鳚(Enedrias fangi)、黄鲫、赤鼻棱鳀、鳀、鲐、矛尾虾虎鱼、小眼绿鳍鱼、斑鰶(Konosirus punctatus)等,而春、冬季鱼类群落则受到干扰后的恢复力较差[42]。因此,该海域秋季鱼类群落可以承受较高的捕捞压力[43];而在冬季进行捕捞作业时,应更加注重鱼类群落的保护与恢复,减少捕捞强度,保护较大个体亲本,以维持各鱼种的补充数量处于较高水平[43]。
鱼类洄游特征是造成长山列岛邻近海域鱼类功能多样性指数季节变化的重要原因。夏、秋季节,鲐、蓝点马鲛(Scomberomorus niphonius)、黄尾鰤(Seriola lalandi)、细条天竺鲷(Jaydia lineata)、长蛇鲻(Saurida elongata)、黑鲷(Acanthopagrus schlegelii)、蓝圆鲹(Decapterus maruadsi)和油魣(Sphyraena pinguis)等多种洄游性鱼类来该海域生殖和索饵,增加了洄游类型、体型、最大体长、适温性、栖息位置、鱼卵类型等功能性状的范围和取值,使鱼类占据的生态位空间得以增加。因此,夏、秋季功能丰富度指数显著高于春、冬季,表明夏、秋季鱼类群落对环境的适应能力和对生态入侵的承受能力更强[44],这也在脆弱性和恢复力指标中得以体现。功能均匀度指数在春季最高,原因是春季黄鲫、赤鼻棱鳀、方氏云鳚等低营养级的浮游生物食性鱼类相对生物量增加,增加了营养级、口的大小等摄食性状的广度。因此,春季鱼类群落生态位更加分化,对于资源的利用更加充分[44]。功能离散度指数在秋季最低,原因是秋季种类数和相对生物量虽然最高,但是例如黄鮟鱇、长蛇鲻、油魣、星康吉鳗(Conger myriaster)、小带鱼(Eupleurogrammus muticus)等较高营养级鱼类等竞争激烈,生态位重叠严重,鱼类群落表现为竞争排斥状态[45]。
环境因子的差异引起了长山列岛邻近海域功能多样性的空间差异。春季随着水深变浅,底层水温逐渐增高,赤鼻棱鳀、黄鲫等暖温种逐渐向温度较高的西部海域迁移,增加了西部海域摄食性状的广度。因此,春季功能均匀度指数表现出西高东低的分布趋势,与底层水温显著正相关。夏季功能丰富度指数与底层水温显著负相关,原因可能是夏季表层水温的变化范围(24.85~27.91℃)超过了绝大多数鱼类的最适水温,较低温区域的鱼类种类数和相对生物量均高于高温区域。黄海影响到长山列岛东北部海域的冷水团,主要为低温高盐水[28],长山列岛东部海域表现出低温高盐的性质,因此夏季功能丰富度指数表现出东高西低的分布趋势。冬季功能丰富度指数与底层水温、表层水温、底层盐度和表层盐度均显著正相关,长山列岛东部海域受到黄海暖流高温高盐水的影响[29],水文条件表现出高温高盐的趋势,因此冬季功能丰富度指数呈现出东高西低的分布趋势。冬季功能均匀度指数与底层水温和表层水温显著负相关,同时呈现出西高东低的分布趋势,这可能是在水温的驱动下,多数暖温种在长山列岛东部海域聚集,但鱼类资源利用方式相同,鱼类群落表现为竞争排斥,而西部海域鱼类群落表现为环境过滤作用[46]。

表 A1 长山列岛邻近海域鱼类名录及功能性状Table A1 List of fish species and their functional traits in the adjacent waters of the Changshan Islands
由于物种本质特征极大地影响了物种在生态系统的功能[47],基于功能性状的多样性研究有助于深入理解物种与生态系统功能之间的联系以及环境因素和干扰对群落结构的影响。除本研究的13种功能性状外,鳍的形状、位置、眼的位置等重要功能性状,在数据充足的情况下,可根据需要适当选择,从而更全面准确地理解鱼类功能多样性的变化。
5 结论
鱼类作为海洋生态系统的重要次级消费者,研究鱼类群落功能多样性具有重要意义。在环境因子的影响下,长山列岛邻近海域鱼类群落优势功能性状的季节变化明显,功能多样性具有一定的季节变化,其空间格局也展现出复杂性、异质性的特点。
附录

续表 A1

续表 A1
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
Plasma Science and Technology(2021年2期)2021-02-27
儿童时代·幸福宝宝(2020年9期)2020-09-08
东南文化(2016年4期)2016-08-22
公务员文萃(2016年7期)2016-07-15
中学生天地(A版)(2015年7期)2015-08-06
读者(乡土人文版)(2013年10期)2013-04-12
杂文选刊(2010年7期)2010-05-14
