
死皮对橡胶树树皮线粒体超微结构及活性氧代谢的影响
2023-01-11 03:00:18王立丰代龙军郭冰冰
植物研究 2023年1期
杨 洪 王立丰 代龙军 郭冰冰
(1. 中国热带农业科学院橡胶研究所,海口 571101;2. 农业农村部橡胶树生物学与遗传资源利用重点实验室,海口 571101;3. 省部共建国家重点实验室培育基地—海南省热带作物栽培生理学重点实验室,海口 571101;4. 农业农村部儋州热带作物科学观测实验站,海口 571101)
橡胶是重要的工业原料和战略物资,根据来源不同分为天然橡胶和合成橡胶。天然橡胶在弹性、耐磨性和延展性等方面具有合成橡胶不可替代的优势,在一些重要工业领域(如航空、航天和重型汽车等制造业)具有广泛的应用。巴西橡胶树(Hevea brasiliensis,简称橡胶树)是重要的热带经济作物,其产生的胶乳是天然橡胶的主要来源。我国是天然橡胶消费大国,但是适宜种植橡胶树的地区和土地面积却十分有限,使得我国的天然橡胶消费不得不长期依赖进口。随着社会经济的不断发展,我国对天然橡胶的需求量还在逐年增加,供需缺口还将进一步增加。目前,我国天然橡胶产业面临植胶区域有限且种植趋于饱和、天然橡胶价格持续低迷等问题,已很难通过扩大种植面积来增加产量[1]。面对严峻的现实情况,大幅度提高橡胶树单位面积产量是促进我国天然橡胶产业健康发展及缓解天然橡胶供需失衡压力的最佳途径。
死皮(tapping panel dryness,TPD)是一种橡胶树所特有的现象,主要表现为橡胶树产排胶过程割面部分或全部丧失排胶能力。与其他作物不同,橡胶树的生产周期长达30 年[2]。死皮是影响橡胶树高产稳产的重要因素之一,也是世界天然橡胶产业面临的共性问题。据统计,我国主要植胶区域死皮发生率高达24.71%,停割率为14.55%[3]。此外,随着高产无性系(一般表现为耐割耐刺激能力差)和乙烯刺激采胶技术的推广,死皮发生率和严重程度呈逐年上升趋势[4-5]。死皮已成为橡胶树单位面积产量提高的主要限制因子。死皮是一种多因素引起的复杂的生理综合症,至今对其发生发展规律的认知还十分有限。已有研究表明,活性氧(reactive oxygen species,ROS)代谢失衡是橡胶树死皮发生的关键过程[6-10]。线粒体是细胞活动的“能量工厂”,同时也是ROS的重要产生部位。在植物细胞质基质和线粒体中,抗氧化酶和抗氧化代谢物共同调节细胞内ROS产生与清除平衡[11]。
目前,围绕橡胶树死皮的发生发展规律[3,12]、生理[13]、分子机制[14-16]及死皮防治[17-18]等多个方面已经进行了大量研究,而在树皮细胞超微结构及抗氧化系统方面的研究较少。本文通过研究不同程度死皮橡胶树树皮线粒体超微结构及ROS相关基因表达量变化规律,从微观角度探讨其与橡胶树死皮动态发生过程的关系,以期深化对橡胶树死皮发生机理的理解,为大幅提高橡胶树单位面积产量和制定有效的TPD 防控措施提供理论依据。
1 材料与方法
1.1 材料
所有样品均采自中国热带农业科学院试验场红洋队开割10 a(种植于2003 年,开割于2010 年)的橡胶树品系热研73397,于2020 年8 月开始割胶,根据连续6 刀(常规割胶,S/2 d/3)的排胶情况确定死皮程度。
1.2 试验方法
1.2.1 死皮等级确定
将所选橡胶树分为健康、轻度死皮和重度死皮,分类标准如下:排胶正常且胶乳流速均匀的树为健康树(标记为H),死皮长度为2 cm至占割线总长度的1/4为轻度死皮(标记为T2),死皮长度介于割线总长度的1/2至3/4为重度死皮(标记为T4)。
1.2.2 电镜样品制备与超微结构观察
将确定部位割胶耗皮用报纸接住后,立即将靠近形成层部位的韧皮组织切下,并在冰上将切下的组织切割成1 mm×1 mm 的小块,然后迅速投入体积分数2%戊二醛固定液中固定,带回实验室用真空泵抽气直至样品沉底,室温放置2 h进行前固定,之后用pH 为7.4 的磷酸缓冲液(PBS)漂洗3 次,每次15 min,再用1%的锇酸进行后固定5 h,并用0.1 mol·L-1PBS(pH 为7.4)漂洗3 次,每次15 min;将固定好的样品依次放入体积分数30%、50%、70%、80%、90%、95%、100%的乙醇溶液进行逐级脱水,每次1 h;然后将样品依次放入丙酮与环氧树脂812包埋剂体积比为3∶1、1∶1、1∶3、0∶1的混合液中进行渗透处理,每次120 min,将渗透好的样品插入包埋板后37 ℃烘箱过夜,再在60 ℃烤箱聚合48 h;最后用Leica 冷冻超薄切片机(UC7,德国)进行超薄切片,厚度为60~80 nm。切片再经醋酸铀和枸橼酸铅各染色25 min 后,在透射电镜(HT7700,日本)下观察并拍照。
1.2.3 RNA提取及cDNA反转录
橡胶树树皮总RNA 采用RNA prep Pure Plant Kit(DP441,天根生化科技(北京)有限公司)试剂盒提取,提取步骤按试剂盒说明书操作。提取的总RNA 通过测定OD260和OD280进行质量分析和定量。cDNA 反转录采用PrimeScriptTMRT reagent Kit with gDNA Eraser试剂盒(TaKaRa)进行。
1.2.4 qRT-PCR检测
根据橡胶树基因组数据库[19]中获得的ROS 相关基因序列,利用Primer Premier 6.0 软件设计qRT-PCR引物。引物序列如表1所示。

表1 本研究所用引物序列Table 1 Primer sequence used in this study
1.2.5 数据统计分析
用Excel 2016 对基因表达数据整理,采用SPSS 23 软件t-test 工具对基因表达倍数进行差异显著性分析,采用Origin 9.1软件作图。
2 结果与分析
2.1 死皮发生过程树皮线粒体超微结构的变化
健康树树皮线粒体(M)未见明显肿胀,结构完整,大部分线粒体膜完整,嵴清晰(见图1A)。轻度死皮树线粒体(M)轻微肿胀,大多膜结构完整,其内基质局部略显变淡,嵴减少(见图1B)。
重度死皮树树皮线粒体(M)严重肿胀,大部分线粒体呈不规则形变大,膜内基质溶解,嵴消失、空泡变,部分严重者,膜破损,基质外溢(见图1C)。
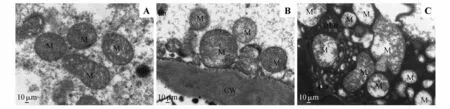
图1 树皮线粒体超微结构A.健康树;B.轻度死皮树;C.重度死皮树;M.线粒体;Pb.质体小球;CW.细胞壁;RER.粗面内质网;V.液泡;Go.高尔基体Fig.1 Ultrastructure of mitochondrial of rubber tree bark A.Healthy tree;B.Mild TPD tree;C.Severe TPD tree;M.Mitochondrial;Pb.Plastoglobuli;CW.Cell wall;RER.Rough endoplasmic reticulum;V.Vacuole;Go.Golgi apparatus
2.2 死皮发生过程抗氧化酶基因的表达
为明确橡胶树死皮发生过程ROS 产生与清除关键基因的表达模式,利用qRT-PCR 技术分析了过氧化物酶(peroxidase,HbPOD1~7)、过氧化氢酶(catalase,HbCAT)基因的表达模式(见图2)。结果显示,HbPOD1、HbPOD4、HbPOD5、HbPOD6和Hb-POD7基因在死皮树树皮中的表达量均显著低于健康树且死皮越严重其表达量越低。HbPOD2基因在轻度、重度死皮树树皮中表达均显著高于健康树,分别为健康树表达量的15倍、12倍。HbPOD3基因在轻度死皮树树皮中的表达量最高,约为健康树的2.6倍,而在重度死皮树树皮中的表达量与健康树基本一致。HbCAT基因在树皮中的表达随着死皮的发生逐渐降低,其中在健康树和轻度死皮树中的表达量分别为重度死皮树中的2.6倍、1.8倍(见图2)。
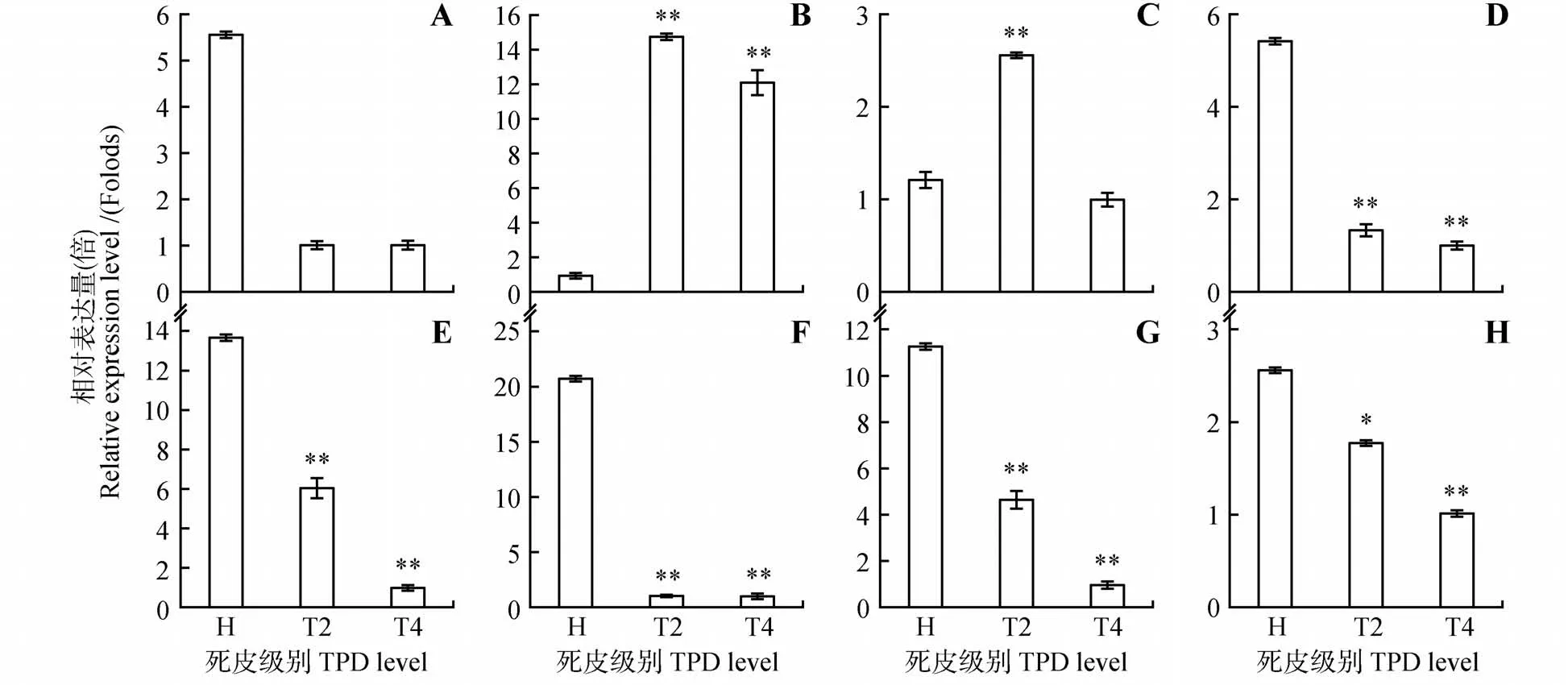
图2 抗氧化酶基因在橡胶树死皮发生过程的表达A~G.HbPOD1-HbPOD7 基因表达;H.HbCAT 基因表达;死皮级别H,健康树;死皮级别T2,轻度死皮树;死皮级别T4,重度死皮树;*和**分别表示死皮树与健康树差异显著(P<0.05)和极显著(P<0.01);下同Fig.2 Expression of antioxidant enzymatic genes during TPD process in H.brasiliensis A-G.The expression of HbPOD1-HbPOD7;H.The expression of HbCAT;TPD level H,Healthy tree;TPD level T2,mild TPD tree;TPD level T4,Severe TPD tree;* and ** indicate significan(tP<0.05)and extremely significan(tP<0.01)differences between TPD trees and healthy trees,respectively;The same as below
2.3 死皮发生过程抗氧化代谢物基因表达模式
通过荧光定量PCR 技术分析了重要抗氧化代谢物基因在橡胶树死皮发生过程的表达量,含谷胱甘肽-S-转移酶(glutathione S-transferase,HbGST1~4)基因4 个、多酚氧化酶(polyphenol oxidase,HbPPO)、L-抗坏血酸氧化酶(L-ascorbate oxidase,HbAAO)基因各1个(见图3)。结果显示,HbGST1、HbGST2、HbPPO3个基因在死皮树树皮中的表达量高于健康树,不同的是HbGST2和HbPPO在重度死皮树的表达略低于轻度死皮树,HbGST1的表达量随着死皮严重程度升高;其余的3个基因(HbGST3、HbGST4、HbAAO)在死皮树树皮中的表达量均低于健康树,其中Hb-GST3和HbAAO的表达量在重度死皮树中表达量最低,HbGST4基因的表达量不受死皮程度影响。
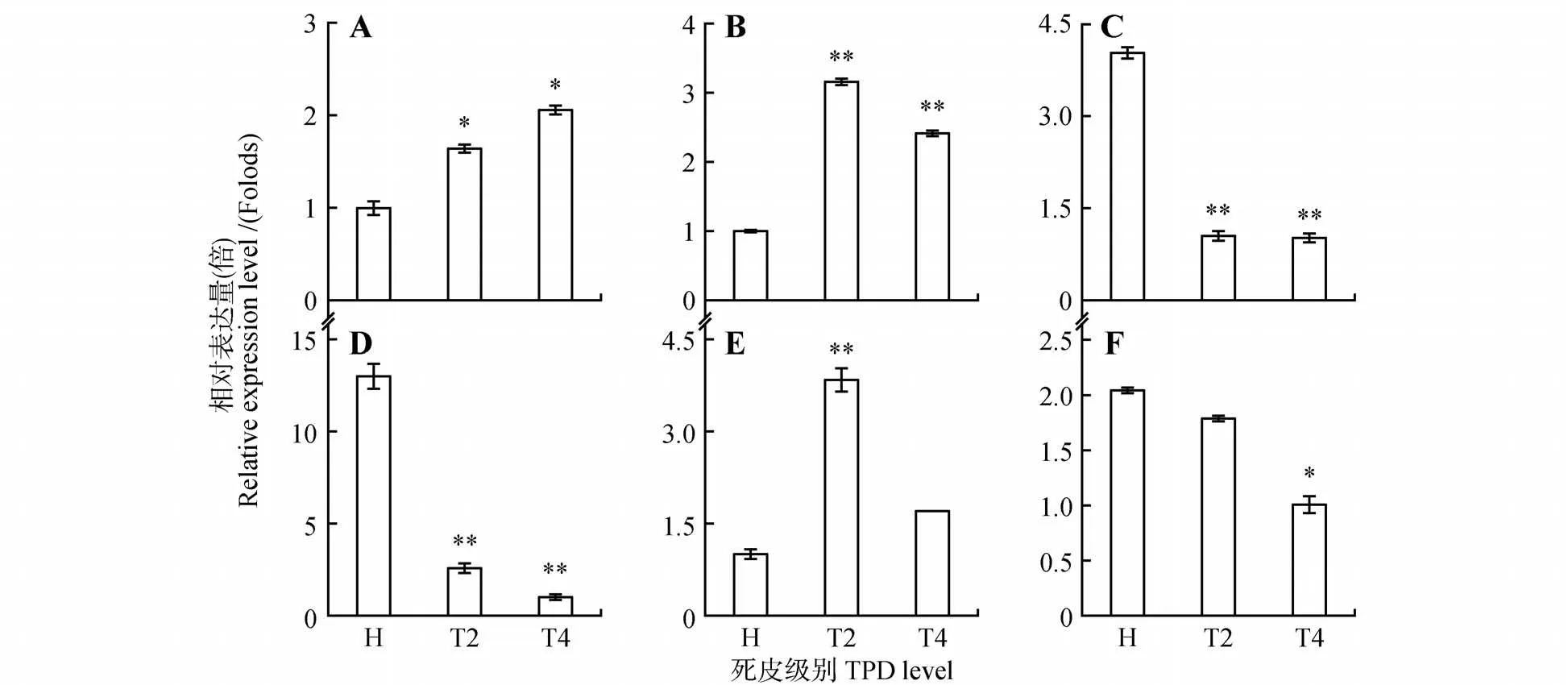
图3 抗氧化代谢物基因在橡胶树死皮发生过程的表达A~D.HbGST1-HbGST4基因表达;E.HbPPO基因表达;F.HbAAO基因表达Fig.3 Expression of antioxidant metabolites genes during TPD process in H.brasiliensis A-D.The expression of HbGST1-HbGST4;E.The expression of HbPPO;F.The expression of HbAAO
3 讨论
细胞及细胞器超微结构分析是研究植物异常生理活动的重要细胞学手段。卢亚莉等[20]利用光学显微镜技术研究了不同程度死皮树树皮中的乳管、筛管、石细胞和单宁等的形态结构的变化,其研究结果表明随着死皮程度的增加乳管排列越加紊乱无规律,膨大乳管数增加,筛管直径减小,石细胞和单宁细胞增多。光学显微镜下,三级树正常排胶部位树皮结构与健康树基本一致[20]。线粒体是细胞中重要的细胞器,也是细胞新陈代谢的重要组成部分。在植物中,线粒体通过与光合作用、光呼吸和细胞质代谢紧密相连维持细胞内氧化还原平衡[21]。因此,线粒体结构的完整与细胞稳态、正常生理状态息息相关。本研究的电镜结果显示,死皮发生过程橡胶树树皮线粒体超微结构发生明显变化(见图1)。健康树树皮大部分线粒体膜完整,嵴清晰,无明显损伤。当死皮发生时,固缩、变大、变长的线粒体基质开始溶解、嵴开始消失,这必将对三羧酸循环和生物氧化代谢产生影响,从而影响细胞内物质和能量代谢。随着死皮程度的进一步加深,线粒体肿胀加深并呈不规则形变,膜内基质大部分溶解,嵴消失、空泡变,部分严重者,膜破损,基质外溢。
过度割胶和过度刺激引起的ROS产生与清除失衡是引发橡胶树死皮的重要因素[14-15,22]。一方面过度割胶和过度刺激增强线粒体中呼吸代谢,从而导致ROS大量产生;另一方面,割胶后ROS清除相关物质随着胶乳排出而含量极低,细胞ROS清除能量减弱,导致ROS 的大量累积[4]。生理生化参数测定显示,死皮树中NAD(P)H 氧化酶和过氧化物酶活性显著提高,黄色体破裂指数和ROS水平升高,还原性硫醇和抗坏血酸等ROS 清除剂含量降低,超氧化物歧化酶、过氧化氢酶、多酚氧化酶活性降低[4,23]。本研究利用荧光定量PCR 技术分析了ROS 产生、清除关键酶基因HbPODs、Hb-CAT(见图2)和ROS 清除相关抗氧化代谢物Hb-GSTs、HbPPO、HbAAO基因的表达(见图3)。基因表达结果显示并不是所有的过氧化物酶在死皮树中高表达,仅有HbPOD2、HbPOD3在死皮树中的表达量显著高于健康树,其他5 个POD基因死皮树的表达量均低于健康树,预示HbPODs基因家族具有不同的分工。许闻献等[24]通过分析过氧化物酶的同工酶分析发现造成死皮的过度割胶和过度刺激是通过诱导一些主酶带的活性增强从而导致过氧化物酶活性提高,并不是所有同工酶活性都增强。根据本研究的结果推测HbPOD2和HbPOD3可能是橡胶树死皮发生过程中过氧化物酶活性增强的主效过氧化物酶,未来可作为开发割胶强度、刺激强度和死皮发生的“标志”基因的备选基因。死皮树HbCAT基因的表达下调且下调程度与死皮严重程度呈正相关,这与郭秀丽等[13]的生理生化测定结果一致。谷胱甘肽-S-转移酶是广泛存在于动植物中的一种多基因编码的多功能超家族蛋白酶,在清除生物和非生物胁迫产生的氧化损伤中起着重要的作用[25-26]。在植物中,GSTs 基因家族成员在25~60,根据序列一致性、基因结构和活性位点可分为6 类[26]。范玉洁等[27]克隆了首个橡胶树GSTs 家族成员HbGSTU1,该基因的表达受逆境胁迫和激素诱导等因素的调控,并且在死皮树胶乳中的表达量较健康树高,且随着死皮程度的增加出现递增的趋势。本研究共分析了4 个不同于HbGSTU1的橡胶树GSTs 家族基因的表达模式,结果显示HbGST1和HbGST2在死皮树树皮中的表达量高于健康树,这与范玉洁等[27]的研究结果一致,而HbGST3和HbGST4的表达趋势则正好相反(见图3)。这些基因表达的差异可能是因为HbGST1、HbGST2与HbGSTU1属于同一亚类,具有类似的生物学功能,而HbGST3、HbGST4属于另一GSTs基因家族亚类,具有不同的生物学功能。PPO 是一种结构复杂的含铜氧化还原酶,也是橡胶粒子凝固不可或缺的组份[28]。本研究也发现HbPPO在死皮树中上调表达(见图3)。抗坏血酸-谷胱甘肽循环(ASA-GSH)是清除ROS 的重要抗氧化途径[29]。AAO 是植物中广泛存在的多铜氧化酶,也是ASAGSH 循环系统中的关键酶之一[30]。本研究发现,HbAAO在死皮树中表达下调(见图3),暗示HbAAOs参与橡胶树死皮发生过程ROS清除。
橡胶树死皮是一个持续的动态发生过程,树皮细胞或细胞器结构的变化可能是其发生的起始。因此,本研究利用透射电镜技术观察了健康树和不同程度死皮树树皮线粒体超微结构,并对ROS代谢相关基因的表达模式进行了分析。本研究发现线粒体等细胞器氧化损伤是橡胶树死皮发生的关键过程,而ROS相关基因可作为下一步开发死皮发生、割胶和刺激强度监测的潜在“标志”基因。
猜你喜欢
西南大学学报(自然科学版)(2024年3期)2024-03-18 08:20:16
农业科技与信息(2021年9期)2021-12-07 01:53:49
农业科技与信息(2021年7期)2021-12-06 07:49:37
农业科技与信息(2021年8期)2021-12-06 04:24:43
北京信息科技大学学报(自然科学版)(2020年5期)2020-11-19 07:05:24
世界热带农业信息(2019年2期)2019-05-17 10:05:04
小资CHIC!ELEGANCE(2017年9期)2017-07-10 10:50:28
热带作物学报(2017年9期)2017-05-30 05:45:25
现代检验医学杂志(2016年3期)2016-11-15 01:59:34
法医学杂志(2015年4期)2016-01-06 12:36:34
