
不同生境的木棉幼苗生长及光合特性1)
2023-01-10 08:23:38王妍方黄郑雯李仕杰杨巧程希平
东北林业大学学报 2023年1期
王妍方 黄郑雯 李仕杰 杨巧 程希平
(西南林业大学,昆明,650224)
植物生长及光合特性变化作为反映植物与生境协同重要的有效标志,能体现植物对环境的适应能力和自身生长发育规律。当今各地区气候变化十分显著,植物生境在时间和空间上不断变化,植物会根据自身各器官的需求和功能,权衡对各器官的资源投入,对转换来的生物量进行适应性分配,促进自身对生境因子的协同和利用[1]。同种植物生长及光合特性差异不仅可以反映生存环境的质量,也可以反映出物种对于环境的忍受力。已有研究表明,植物在不同生境下表现出不同的生理和光合特性,戚文涛等[2]、赵玉文等[3]发现不同地理种源的种子和植株变异与生态环境相关性存在明显差异;也有相关学者发现植物对生境响应强烈,光合生理特性反映出地域性差异[4-5];吴金山等[6]、赵孟良等[7]通过生长和光合特性比较探讨了植物对不同生境的适应习性的异同。物种在适应环境中会表现出不同的生存策略,其中既有遗传因子影响,还与生存环境息息相关。
木棉(BombaxceibaLinnaeus)属木棉科木棉属植物,为落叶大乔木树种,具有适应性强、耐干、喜光的特点,分布于马来半岛、印度尼西亚及我国华南、香港、台湾等地区,对区域内的水土保持和生态恢复建设有很大意义[8-9]。目前,对木棉在景观、种质、纤维成分等方面独特的利用已有相关研究[10-13],而对自然生长状况下不同生境对木棉幼苗生长和光合特性的影响机制研究较少。
本文以广布于元江干热河谷和勐腊热带雨林地区的木棉幼苗为试验材料,对其生长特征和光合特性进行测定,分析木棉幼苗的株高、冠幅、地径等生长特征和光合特性在不同生境下的差异及生存策略,有利于揭示生境对植物生长的影响,以期为木棉的培育、营林以及把握其空间分异机理提供参考。
1 试验地概况
试验地位于云南省元江国家级自然保护区(102°18′~102°20′E,23°24′~23°28′N)(下称元江)和西双版纳国家级自然保护区勐腊片区(100°58′~100°60′E,21°51′~21°55′N)(下称勐腊)。元江地区植被以稀树灌草丛为主,气候干热特征明显,年均降水量较少,雨季(5—10月)降水量约583 mm,占全年降水量的77.5%,季节性干旱明显,温度高,年均气温约24.5 ℃,光照强,年均日照时长约2 397 h,土层浅薄,土壤主要以燥红壤和赤红壤为主,属于典型的干热河谷[14]。勐腊地区植被类型以热带季雨林、热带雨林为代表,植被覆盖率高,气候湿热特征显著,降水充足,年降水量1 200~1 600 mm,湿度大,约82%的降水集中在雨季(5—10月),年均气温约23 ℃,年均日照时长约1 955 h,光能利用潜力大,土壤较为肥沃,主要类型为砖红壤[15]。
2 研究方法
试验材料为广布于云南省元江与勐腊地区的自然生长的木棉实生苗,每个生境下随机选取30株自然生长幼苗,选取规则为长势基本一致,叶片位置、朝向相同,每株进行3组重复测定。
木棉幼苗生长特征测定:于2019年11—12月和次年6—7月,分别测量两地区木棉幼苗样株的地径、株高、一级分枝数、冠幅、枝下高,计算幼苗在干季的增长量。光合测定结束,将幼苗全株挖出,洗净擦干后,分根、茎和叶测算鲜质量后,杀青、烘干至质量恒定,分别计算木棉幼苗各器官生物量的分配比。
木棉幼苗光合特性测定及光合作用光响应曲线绘制:于2020年6—7月,选择晴朗天气,在自然状态下对两地区木棉实生幼苗的光合日变化相应指标进行测定。使用Li-6400XT便携式光合测定仪,从08:00—18:00,每隔1 h测定1次,对每株幼苗测量3~5片正常生长叶片,记录光合有效辐射(PAR)、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2摩尔分数(Ci)等光合参数。水分利用效率(WUE,μmol·mmol-1)利用公式WUE=Pn/Tr进行计算,光能利用效率(LUE,%)利用公式LUE=Pn/PAR进行计算[16]。
在2020年6—7月,同期晴朗天气的上午10:00—12:00,使用Li-6400XT便携式光合测定仪绘制两地区木棉幼苗的光合作用光响应曲线。绘制条件设置如下:叶室温度为(29.0±0.5)℃;气路设置为开放,空气流速控制为500 μmol·s-1;相对湿度控制在50%左右;光照梯度依次设置为2 000、1 800、1 600、1 500、1 000、800、500、200、100、50、30、10和0 μmol·m-2·s-1。木棉幼苗叶片的Pn与PAR之间的关系,通过非直角双曲线模型进行拟合[17]。
数据处理:试验数据处理及制图采用Excel 2019及Origin 2018软件完成。利用SPSS 26软件中的独立样本t检验验证不同生境的木棉幼苗生长性状的差异显著性,使用Pearson相关性分析方法分析生长性状和光合特性之间的相关性。
3 结果与分析
3.1 木棉幼苗在不同生境下的生长特性
从不同生境下木棉幼苗生长性状变化量(表1)可知,2019年11—12月至次年6—7月,元江干热河谷地区的木棉幼苗在地表整体生长的增长量高于勐腊热带雨林地区,但地径增长量略低于勐腊地区。元江地区木棉幼苗的株高、冠幅、枝下高及一级分枝数的增长量大于勐腊地区,存在显著差异(P<0.05);元江地区木棉幼苗地径增长量小于勐腊地区,但差异不显著。这些不同生境下木棉幼苗生长性状变化量体现出植株生长性状对不同生境的权衡关系。
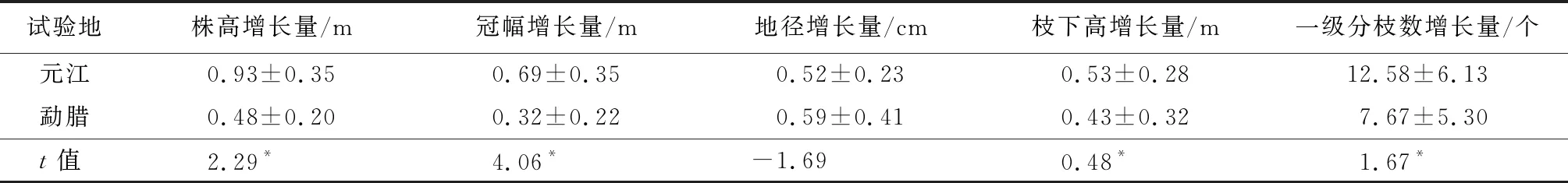
表1 不同生境下木棉幼苗生长变化
生物量是植物生长特征最直接的展示,干热河谷和热带雨林地区这两种极端生境对木棉幼苗的生物量分配具有一定影响。在各器官的生物量占总生物量的百分数方面(表2),元江和勐腊地区木棉幼苗各器官生物量的分配比具有显著差异(P<0.05)。元江地区木棉幼苗的根部生物量占比最高,达49.04%,说明元江地区木棉更倾向于增加根部生长以获取更多的营养物质和水分;而勐腊地区木棉幼苗倾向于提高地上生物量(占总生物量的63.39%)的投入,以便于地表部分向上及向外生长,增强光能捕捉及利用能力,从而产生更多的能量,这是植株为适应生境差异而采用的不同生长策略。
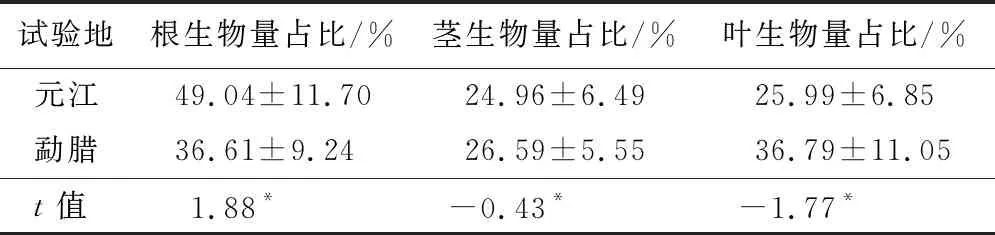
表2 木棉幼苗各器官生物量占总生物量的比例
3.2 木棉幼苗在不同生境下的光合特性
3.2.1 木棉幼苗光合参数变化
由图1可知,两地区木棉幼苗Pn日变化为双峰型曲线,均有“午休”现象。其中,元江地区Pn在11:00达到第1个峰值,为23.47 μmol·m-2·s-1,在14:00出现次峰,为17.46 μmol·m-2·s-1,低谷出现在13:00;勐腊地区Pn也在11:00达到最大峰值,为26.97 μmol·m-2·s-1,在14:00左右出现低谷值,在15:00出现小峰值,为22.11 μmol·m-2·s-1。两地区幼苗净光合速率在达至峰值前增长比较迅速,峰值后下降略慢,勐腊地区幼苗净光合速率整体要高于元江地区。
由两地区Tr日变化(图1)可知,元江地区木棉幼苗Tr呈现单峰型变化,最大值出现在11:00,之后Tr不断下降,直到18:00处于较低水平。而勐腊地区Tr则出现双峰型曲线变化,在11:00达到第一个峰值,随后下降,于13:00出现低谷值,第二个峰值约在14:00出现。勐腊地区木棉幼苗蒸腾速率略高于元江地区。
元江地区及勐腊地区木棉幼苗Gs日变化(图1)均为双峰型变化规律,在早晨出现高值之后呈波状下降。高值过后,元江地区幼苗Gs在11:00出现一个峰值,而勐腊地区木棉幼苗Gs在12:00达到峰值,两地幼苗Gs均在13:00出现低谷值,在14:00达到另一个小高峰,随后逐渐降低。
两地区木棉幼苗胞间CO2摩尔分数变化较为复杂(图1)。两地区幼苗Ci值在早晨08:00为最大值,随时间变化呈波状下降。之后,元江地区木棉幼苗Ci值在14:00出现一个最低值,随后不断上升;勐腊地区木棉幼苗Ci值在08:00之后变化较大,分别于12:00、14:00出现峰值,16:00之后Ci还在上升。
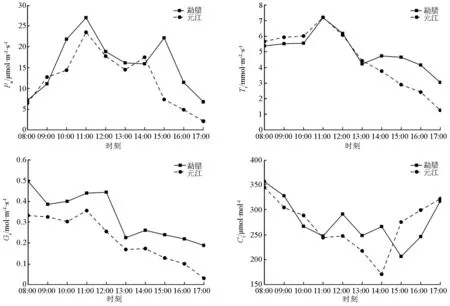
图1 不同生境下木棉幼苗光合参数日变化
3.2.2木棉幼苗光合作用光响应曲线及相关利用效率变化
植物光合作用光响应曲线能够反映自身的光合特性及其对生长环境的协同程度。对元江和勐腊木棉幼苗光合作用光响应曲线实测数据进行拟合,探讨两地区幼苗不同梯度PAR对Pn的影响规律(图2)。两地区木棉幼苗叶片Pn变化趋势相近,两地区木棉幼苗叶片Pn随PAR值的增加呈曲线式上升,当PAR<200 μmol·m-2·s-1时,Pn增加速率较快,接近线性增长,后来逐渐减缓,达到最大净光合速率(勐腊为26.16 μmol·m-2·s-1、元江为23.34 μmol·m-2·s-1)后,出现光饱和现象。当PAR<800 μmol·m-2·s-1时,元江地区木棉幼苗净光合速率要强于勐腊地区,之后勐腊地区幼苗Pn值要高于元江地区,并且在PAR值为2 000 μmol·m-2·s-1时增势仍较为明显,表明勐腊地区木棉幼苗光照强度适应范围及光合作用要强于元江地区木棉幼苗。
植物的水分利用效率能有效评价其水分的利用能力,木棉幼苗能够通过改变WUE状况来适应不同生境,使其在植物群落中表现出更强的竞争能力和生态适应性[18]。由图2可知:元江地区幼苗WUE为双峰型曲线,勐腊地区出现三峰型曲线,同时两者都因为12:00左右的光“午休”现象,出现水分利用率下降现象,元江地区WUE最高值出现在14:00,为4.65 μmol·mmol-1,勐腊地区出现在15:00,为4.75 μmol·mmol-1。在测量中的大部分时刻,勐腊地区木棉幼苗WUE略高于元江地区。
LUE主要用来评估植物通过光合作用将能量转为有机物质的效率,也能初步估算植物生产力大小[19]。图2表明:勐腊地区LUE平均为3.89%,元江地区LUE平均为2.88%,元江地区幼苗光能利用效率略低于勐腊地区;同时两地区幼苗在早晨及傍晚的光能利用率高,中午太阳光照强度大,叶片对光能转化均在11:00左右出现低值;由于植物的“光合午休”,元江地区木棉幼苗叶片LUE在13:00到达全天最低,为0.72%。
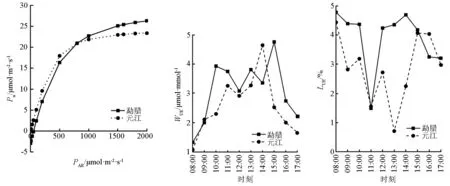
图2 不同生境下木棉光合作用光响应曲线及光能、水分利用效率日变化
3.3 木棉生长性状变化量与光合特性相关性
植物的生长性状和光合特性之间必然存在一定的关联,分析其生物学相关性十分重要。对两地区木棉幼苗的生长性状变化量与最大光强下光合特性进行分析(表3)可知,在生长方面,株高增长量和地径、冠幅、一级分枝数的增长量呈显著正相关(P<0.01),地径增长量和冠幅增长量(P<0.01)、一级分枝数增长量(P<0.05)显著正相关,冠幅增长量和一级分枝数增长量显著正相关(P<0.05),枝下高增长量和株高、地径、冠幅、一级分枝数的增长量为正相关关系,说明木棉幼苗的各生长性状之间相互影响;在光合方面,Pn与Gs、Tr之间呈显著正相关(P<0.01),协同性较高,Pn与Ci呈显著负相关(P<0.05),Gs与Tr显著正相关(P<0.01);此外,受多因素影响,生长性状和光合特性之间显现出复杂关系,但其中株高、地径的增长量与Ci的关系为显著正相关(P<0.05)。
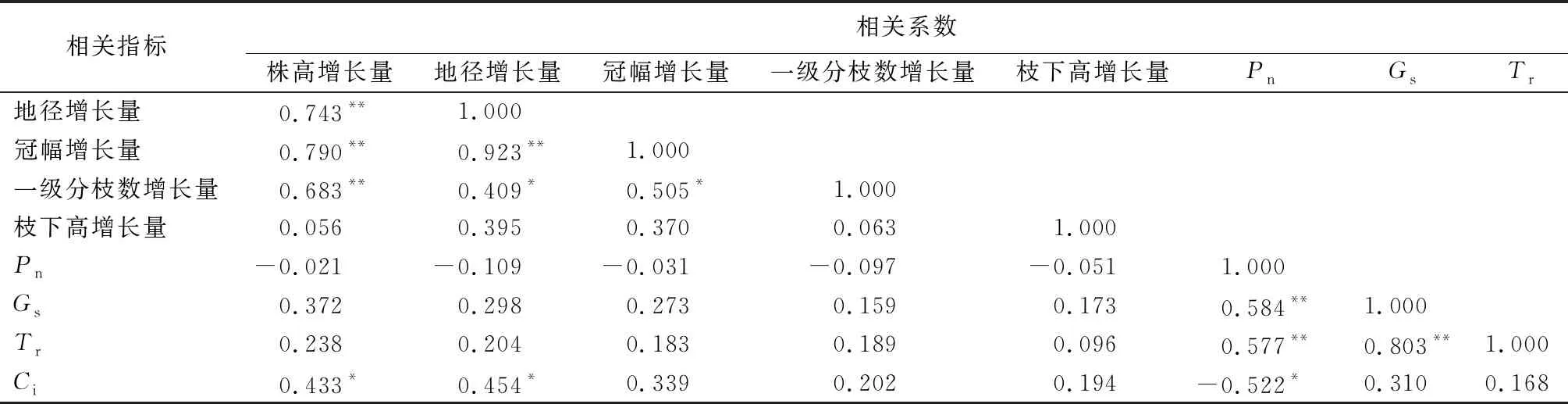
表3 木棉幼苗生长性状变化量及光合特性相关性系数
4 讨论和结论
生境是决定植物生长及光合变化的重要影响因素之一,植物自身生长变化也可以有效反映对生境变化的适应,是评估植物对环境适应性的关键指标,生境中胁迫因子多,植株发育会受到钳制,胁迫越严重受钳制作用越明显[20-21],且同种植物同一季节生长、光合等特性发生差异与生境息息相关[22]。本研究发现,不同生境下木棉幼苗生长特性出现较大差异,元江地区木棉幼苗株高、冠幅、枝下高等生长特征在干季的增长量高于勐腊地区,木棉幼苗根部生物量所占比重大。植物对生境变化十分敏感,尤其是生境往不良方向变化下,植物会通过改变生物量的分配策略来减弱不利环境的影响,生产的光合物质及能量会优先分配到最需要的部位[23-24]。元江干热河谷地区,水热条件组合较差[25],植物幼苗为了竞争资源,充分利用水分、光照等环境资源,会加快其在初期的生长速率,生理反应强,使得地表部分迅速增长[26]。在干热河谷地区,水是植物生长发育的限制因子,幼苗为了更好的吸收水分及养分,将更多地生物量投入根系,体现出植物对干旱的抗逆反应[27]。由于勐腊地区较湿热,水热组合优于元江地区,木棉幼苗在干季和湿季都能得到较好的生长,所以其地表生物量较元江地区高。本研究中木棉幼苗的株高、地径、冠幅及一级分枝数性状增长量之间显著相关,是因为植物性状的生长发育是相互联系的,这是植物在光合作用、养分因素等差异下产生的资源权衡策略[28]。
光合作用是植物生产能量与物质吸收利用的复杂过程,其日动态变化反映了植物自身遗传特征与对生境的适应能力[29-30]。木棉幼苗对生长环境条件的适应导致两地区光合特性存在一定差异。由两地幼苗相关光合参数可知:在自然光照条件下,两地区Pn日变化均出现双峰现象,均有明显的“午休”,导致植物光合速率变化的因子有气孔限制和非气孔限制两个方面[31]。元江地区木棉幼苗净光合速率日变化在13:00达到波谷,此时Gs、Ci值呈下降趋势,说明气孔限制是元江木棉幼苗“光合午休”的主要因素,元江气候更为干旱,植物为了降低对水分的消耗,也会导致植物幼苗气孔导度较低;而勐腊地区木棉幼苗净光合速率日变化达到波谷时,Gs、Ci值波动上升,是由于木棉幼苗叶片吸收的光能多,而出现碳同化能力受限的非气孔限制因素。雨季两地区幼苗LUE值,在早晨及傍晚高,中午出现低值,表明幼苗可以在叶片尺度上通过改变生理结构来适应不同的光环境,以达到最大限度提高整体光合生产,这与张元媛等[32]研究结果具有相似性。元江幼苗LUE整体低于勐腊,因为勐腊地区空气湿度较大,散射辐射增加,受到的光抑制作用较小,叶片光合能力增强。此外,在高光强作用下,元江地区的Pn低于勐腊地区,出现光抑制,表明元江地区木棉幼苗适应光照强度及光合生产力弱于勐腊地区木棉幼苗。同时,两地区雨季木棉幼苗内部水分变化复杂,勐腊地区木棉幼苗WUE高于元江地区木棉,是因为勐腊地区光合有效辐射较高,植物因此加强蒸腾作用来适应高温环境。史晓亮等[33]也发现植物Pn小于Tr,WUE呈下降趋势,说明WUE与地区立地环境和水分差异相关。本研究中相关性分析显示,木棉光合特性有显著相关性,幼苗株高、地径的增长量与Ci显著正相关,说明幼苗生长受光合影响强烈。
综上所述,木棉均能在干热和湿热两种极端生境下得到良好的生长,说明木棉具有较高的适应能力,抗逆性强,并且良好的水热组合更有利于植物进行光合作用。由于本研究仅对木棉实生幼苗的生长特征及光合特性进行了研究,对异质生境下不同树龄木棉在干季及干、湿季生长及光合特性的耦合还有待进一步深入研究。
猜你喜欢
少先队活动(2021年9期)2021-11-05 07:31:10
Asian Herpetological Research(2020年4期)2020-12-30 06:59:14
现代园艺(2020年9期)2020-05-24 09:22:06
现代园艺(2019年1期)2019-02-18 06:59:42
天然产物研究与开发(2018年8期)2018-09-10 05:48:26
南方农业学报(2018年12期)2018-09-10 05:12:39
种子科技(2018年8期)2018-09-10 02:21:52
城市地理(2017年8期)2017-11-02 14:08:27
安徽农学通报(2017年12期)2017-07-13 15:11:24
读写算·素质教育论坛(2015年7期)2015-08-16 12:49:51
