
黄土高原干涸小河流的芦苇分布特征及其环境适应性研究
2023-01-03 05:42:30高紫月
甘肃科学学报 2022年6期
常 虎,高紫月
(1.中国科学院南海海洋研究所中国科学院边缘海与大洋地质重点实验室,广东 广州 510301; 2.中国科学院南海生态环境工程创新研究院南方海洋科学与工程广东省实验室(广州),广东 广州 510301; 3.中国科学院大学地球与行星科学学院,北京 100049; 4.西北大学法学院,陕西 西安 710127)
在全球气候变暖的大背景下,我国正承受着不同程度的区域气候变化的影响,尤其是在黄土高原沟壑区,全年干旱少雨,夏季极端天气发生频繁。历年来,流经黄土高原的黄河经常发生断流现象,黄土高原沟壑区的小河流干涸现象更加严重,破坏了河流生态系统。河流的干涸现象对两岸的耕地产生了负面影响,如土地盐碱化[1]等。
芦苇(Phragmites australis)是禾本科多年生草本湿生植物,世界范围内广泛生存,能够适应不同生态环境,是构成河岸植物群落的代表性物种[2-3]。芦苇在储存营养元素碳、氮、磷方面有重要作用[4-5]。目前,芦苇的研究多见于湿地芦苇的生态描述、生态修复、生态化学计量、群落调节等方面[6-17],对黄土高原沟壑区干涸小河流的芦苇分布特点和环境适应性的关注不够。
为了探索黄土高原沟壑区干涸小河流的芦苇分布特征和环境适应性,研究以艾家畔村小河沟为例,把干涸小河沟分2个断面,设置8个芦苇采样点位,运用多种数据统计方法进行分析研究,旨在为黄土高原沟壑区干涸小河流的生态保护、修复和再利用提供芦苇方面的资料。
1 研究区概况
研究采样地点(37°41′N,109°50′E)位于陕西省子洲县艾家畔村(见图1)。艾家畔村属于黄土高原沟壑区腹地,年降水量428.1 mm,距离县城25 km,属于典型农村。小河沟是艾家畔村唯一的河流,采样实验时,小河沟河道处于干涸无水状态。

图1 艾家畔村地理位置Fig.1 Geographical location of Ai Jiapan Village
2 研究方法
艾家畔村小河沟河宽5 m,采样点位定在小河沟芦苇长势均匀的位置,方案设置2个断面进行对比实验,根据河道的不同深度设置采样点位(见图2)。采样断面1设置B、D、E 3个采样点位,水平间隔1.5 m。由于小河沟处于干涸状态,河床不平坦,根据与河床低地(小)的距离增设A、C 2个采样点位,A、B、C 3个采样点位在深水区,等距间隔30 cm。采样断面2靠近河床低地(大),设置Ⅰ、Ⅱ、Ⅲ 3个采样点位,水平间隔2 m。

图2 采样点位示意图Fig.2 Schematic diagram of sampling points
每个采样点位设置20 cm×20 cm的样方调查。
观察记录芦苇样本的数量和主要性状。方案计划收集的数据在芦苇样本的采样当天全部记录完毕,以防止芦苇脱水变干影响数据分析。采样实验在2021年夏季实施。
3 结果与分析
小河沟2个断面的8个采样点位共获得170株芦苇样本。对各采样点位的芦苇样本数据进行W检验,在0.05的显著性水平下,数据显著地来自正态分布总体。
3.1 采样断面1
采样断面1的河床起伏较大,共3个采样点位,分别是B、D、E,其中B采样点位在河床中心,A、C采样点位与采样断面1较近,因此一并阐述。
B采样点位的样本数量最少(9株);E采样点位的样本数量最多(45株),芦苇分布呈河床中心少,两岸多的态势。有分枝的芦苇在数量上表现为D、E采样点位远多于B采样点位(0株);D采样点位的样本数量少于E采样点位,但是有分枝的样本数量恰好相反(见图3)。
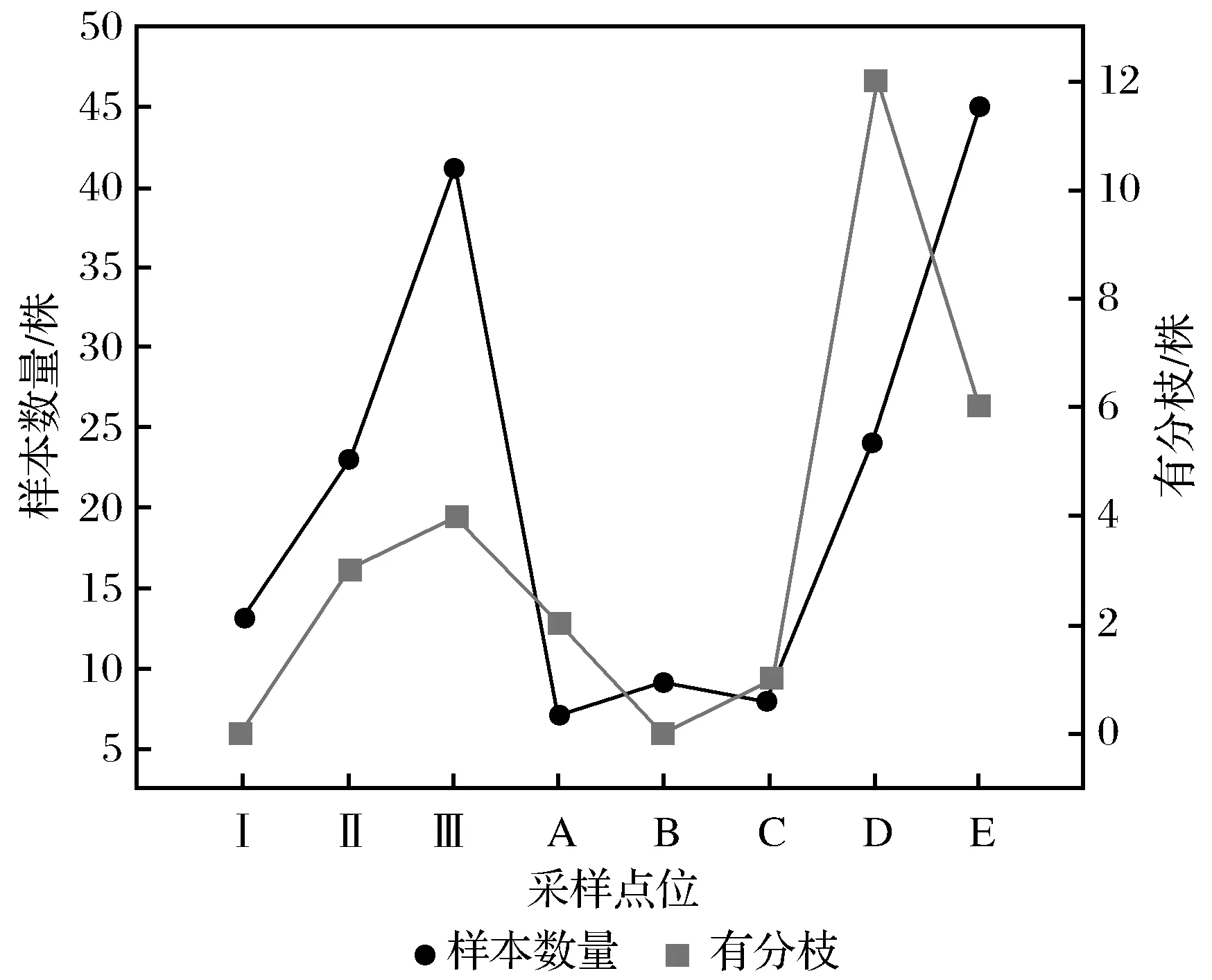
图3 各采样点位样本数量和有分枝的芦苇样本数量Fig.3 The number of samples at each sampling point and the number of samples with branches
B采样点位的直径平均值(3.9 mm)和长度平均值(1 375.4 mm)远大于D、E采样点位;D采样点位的直径平均值和长度平均值都要小于E采样点位(见图4)。
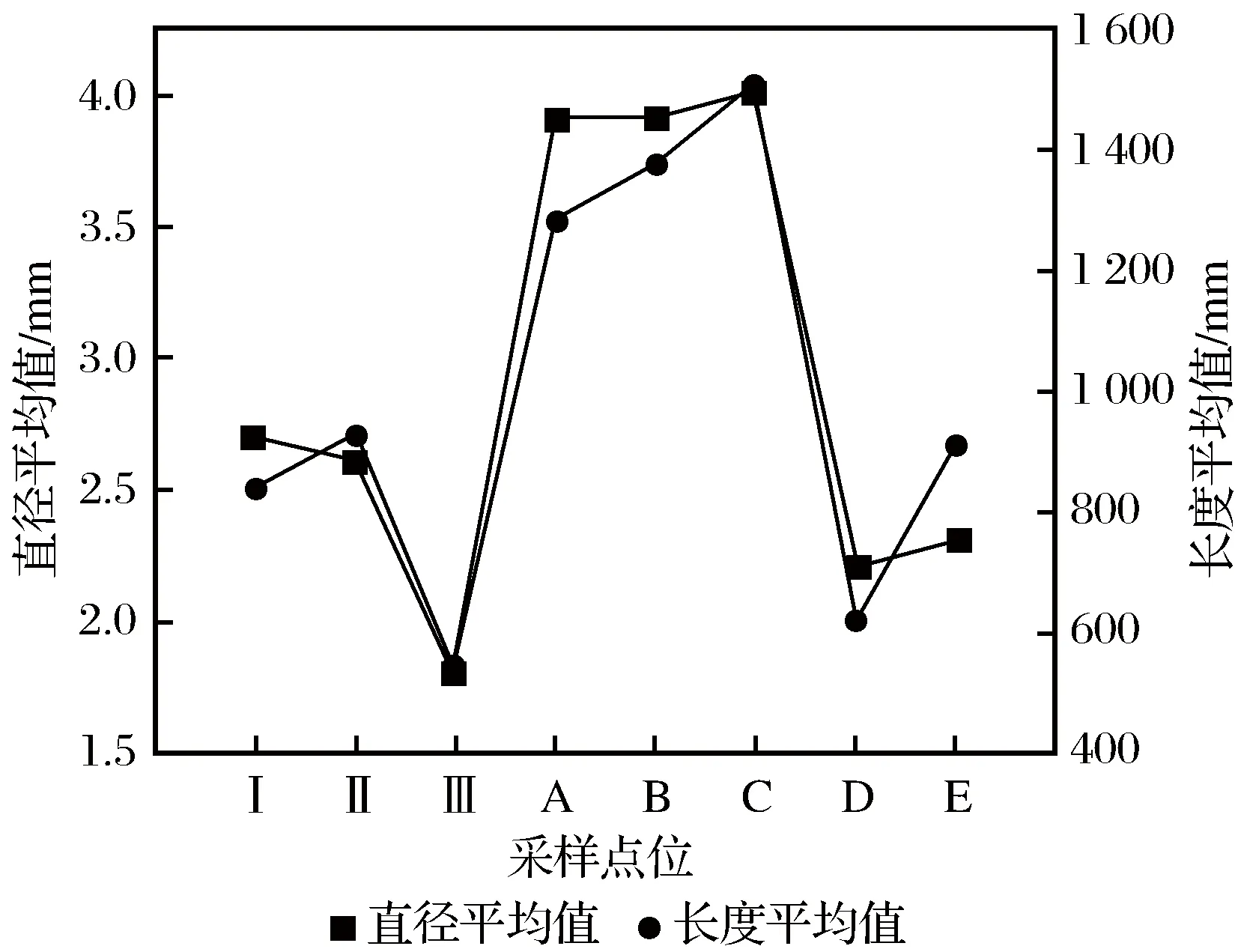
图4 各采样点位样本直径平均值和长度平均值Fig.4 The average value of sample diameter and length at each sampling point
比较采样断面1的3个采样点位,B采样点位的样本分布表现为数量少、无分枝、长度和直径大;D采样点位的样本分布表现为数量适中、分枝很多、长度和直径较小;E采样点位的样本分布表现为数量多、分枝较多、长度和直径适中。
整个采样断面1芦苇的分布表现为河床中心稀疏,但是发育质量好。河床靠近两岸都表现出有分枝的现象,但出现两种变化:一是变矮、变细、分枝增多、数量少;二是长度和直径适中,分枝稍多,数量多。
A、B、C 3个采样点位是河床中心沿河流方向的等距采样,采样点位前方有一河床低地(小)(见图2)。河床低地是河道最深的位置,几乎没有植物生长,可能是由于小河沟干涸之前,水中生物不断向河床低地聚集,生物扰动较大,不具备植物生长所需要的稳定环境,而且干涸前河床低地的生物密度高,水中溶解氧少,河流底泥容易形成缺氧环境,且大量水生生物死亡后有机体分解造成水体pH值降低,导致植物无法生存。
样本数量在A、B、C 3个采样点位的表现基本一致,B、C采样点位的样本数量比A采样点位稍多。有分枝的芦苇数量在A采样点位要多于B、C采样点位,其中B采样点位的芦苇样本没有分枝(见图3)。
A采样点位的芦苇长度平均值(1 278.7 mm)和直径平均值(3.9 mm)最小,C采样点位的芦苇长度平均值(1 504.8 mm)和直径平均值(4.0 mm)最大。A、B、C 3个采样点位的长度和直径平均值呈递增趋势,在河床中心距离河床低地越远,芦苇的分布呈现出数量增、分枝少、变高、变粗的特征。
3.2 采样断面2
Ⅰ采样点位样本数量最少(13株),Ⅲ采样点位样本数量最多(41株)。有分枝的芦苇数量上与采样点位样本数量趋势基本一致(见图3)。
Ⅰ采样点位样本直径平均值最大(2.7 mm),与Ⅱ采样点位非常接近;Ⅲ采样点位样本直径平均值最小(1.8 mm)。Ⅱ采样点位样本长度平均值最大,(925.3 mm),数值接近Ⅰ采样点位;Ⅲ采样点位样本长度平均值最小,仅为546.6 mm(见图4)。
采样断面2的河床剖面较为平坦,对3个采样点位进行比较,Ⅰ采样点位的样本分布表现为数量少、分枝少、长度和直径比较适中;Ⅱ采样点位的样本分布表现为数量较少、分枝较少、长度和直径比较适中;Ⅲ采样点位的样本分布表现为数量多、分枝多、长度和直径小。
整个采样断面2芦苇的分布表现为河床中心发育适中,河床靠近两岸出现两种变化:一是芦苇稀疏,但长势较好;二是芦苇稠密,但变矮变细,而且分枝增多。
3.3 采样断面的比较
采样断面芦苇样本汇总情况见表1。由表1可知,采样断面1和采样断面2的芦苇样本数量基本相同。有分枝的芦苇数量,采样断面1多于采样断面2,主要是因为采样断面1的D、E采样点位离岸较近,而且芦苇样本数量远远大于河床中心的B采样点位。采样断面1的芦苇长度平均值和直径平均值均比采样断面2大,这是因为采样断面2更靠近河床低地,而且采样断面2旁边的河床低地(大)面积显著大于采样断面1附近的河床低地(小)(见图2)。

表1 采样断面芦苇样本汇总Table 1 Summary of Phragmites australis samples from the sampling section
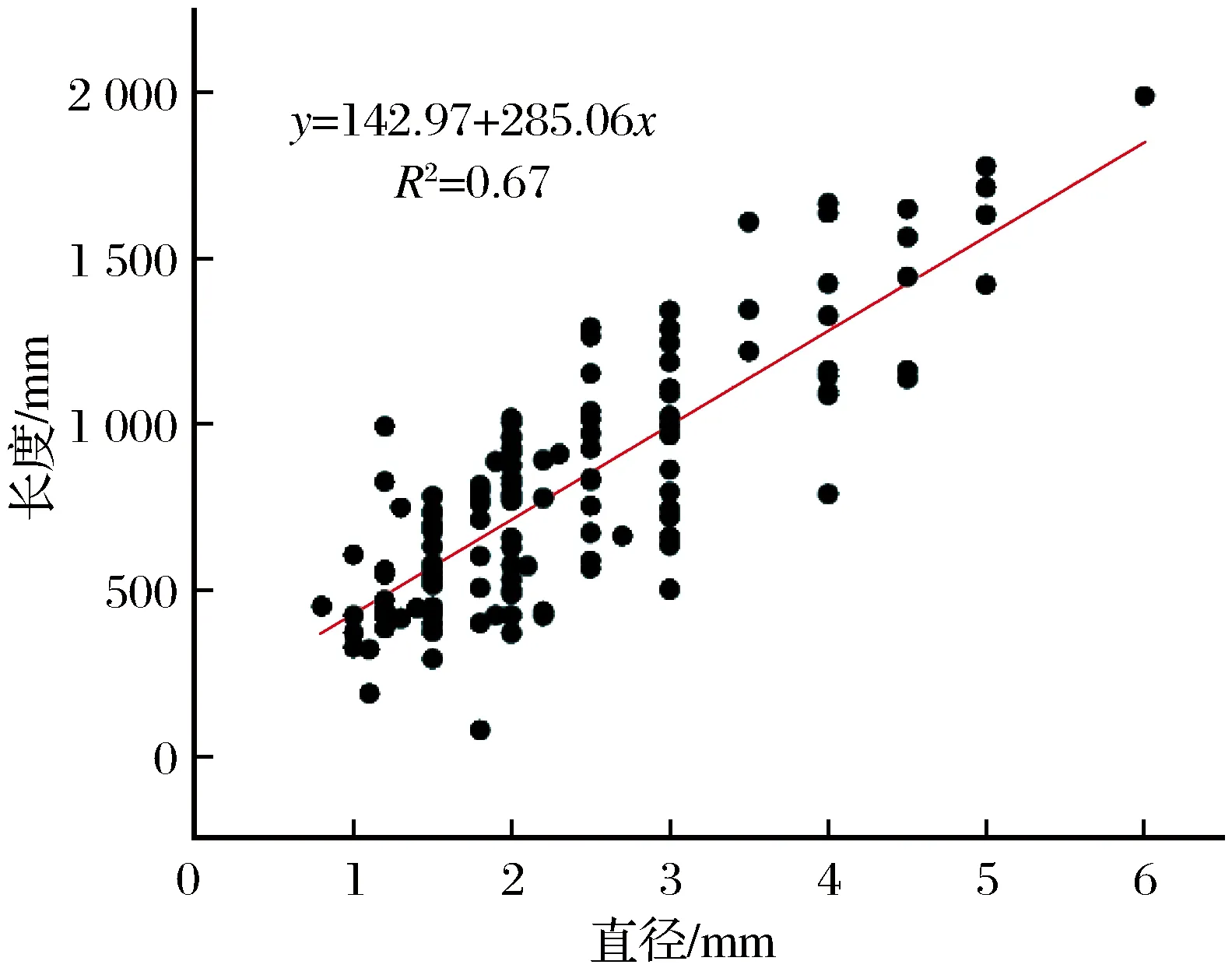
图5 芦苇样本的直径和长度的线性回归分析Fig.5 Linear regression analysis of the diameter and length of Phragmites australis samples
芦苇样本数据统计的过程中,发现存在芦苇主杆折断的现象,具体统计信息见表2。由表2可知,采样断面1主杆折断共8株,采样断面2主杆折断共3株。采样断面1主杆折断主要发生在D、E采样点位,这两个采样点位非河床中心,采样点位河床较浅,靠近岸边。干涸小河沟河道内芦苇密度较大,大型动物活动较为不便。岸边芦苇处于芦苇群落边缘,暴露在群落外环境的干扰之下,受大型动物的干扰相对强烈,而且更容易受到局域小气候变化的影响,例如夏季极端天气冰雹和强降雨造成的洪水[18]。

表2 所有采样点位芦苇主秆折断数量Table 2 Number of broken Phragmites australis rods at all sampling points
采样断面2主杆折断发生在河床中心采样点位Ⅱ和岸边采样点位Ⅲ。采样断面2在河床低地的边缘,河床低地内几乎没有植物生存,采样点位Ⅱ处于芦苇群落边缘地带,暴露在芦苇群落外环境下,因此分布表现与Ⅲ、D、E采样点位一致。
采样断面1的采样点位D、E的样本总量(69株)大于采样断面2的采样点位Ⅰ、Ⅲ的样本总量(54株)。采样断面2的河床比采样断面1更加平坦,环境相对更加稳定,采样点位D、E比采样点位Ⅰ、Ⅲ更早地进入干涸状态,采样断面2的河床低地的蓄水作用延后了干涸时间,一定程度上避免了采样断面2的芦苇被大型动物干扰。
根据统计芦苇主秆折断的采样点位样本,发现主秆折断且有分枝的芦苇与主秆折断的芦苇数量之间的比值接近于1,表明芦苇主秆折断可能促使了分枝的生长。主秆折断且有分枝的芦苇与有分枝的芦苇数量之间的比值说明主秆折断对催生芦苇分枝具有重要作用(见表2)。
3.4 线性回归分析
线性回归是利用数理统计中的回归分析,来确定两种或两种以上变量间相互依赖的定量关系的一种统计分析方法,其运用范围十分广泛。研究对采集的170个芦苇样本进行统计分析,收集芦苇样本的长度和直径作为拟合参数,探究芦苇的长度与直径在数值上的相关性。为了排除其他影响因素,提高结果的准确性,线性回归分析所使用的样本剔除了主杆折断和有分枝的芦苇,筛选后共得到138个满足要求的芦苇样本。
线性回归方程为y=142.97+285.06x,相关系数r=0.82,决定系数R2=0.67。干涸小河沟的芦苇在长度和直径上具有强相关性。长度平均值为822.1 mm,直径平均值为2.4 mm。
4 讨论
光合作用是植物生长的必要条件,研究区芦苇群落的顶部从景观的层次看较为一致且均匀(见图2)。邱天[19]以表观遗传分化解释了不同生境芦苇表型不同的原因,由于表观遗传修饰能在不依赖遗传变异的情况下,在变化的环境中改变基因表达,进而改变表型。干涸小河沟的芦苇在局域生境的适应过程中,形态差异巨大,分布上表现出不同的特点。胡思文等[20]对长江中下游及云贵高原芦苇居群的遗传变异特征的研究表明,不同生境芦苇在形态指标上差异显著,结合遗传差异分析,芦苇的形态变异有一定的遗传基础,且在形态变异上“可塑性”的影响更大。
小河沟河道内河床中心地势相较于两岸低,河床中心芦苇为了获取更多的光照需要更多的营养向上生长,河床中心有水时,河流冲刷力量大,因此河床中心芦苇的数量较少,但是芦苇较高、较粗。
相较于河床中心的芦苇,河床靠近两岸的芦苇生长的地势较高,不需要生长很高就可以得到充足的光照,向上生长的需求不如河床中心的芦苇强烈,因此靠近两岸的芦苇高度比河床中心的芦苇要矮。
干涸小河流内靠近两岸的芦苇数量多、密度大、生存空间有限,但是向上生长的需求不及河床中心的芦苇强烈,因此开始横向占领更多的生存空间。横向占领生存空间的方法就是长出更多的分枝,采样断面1的D采样点位发现一株含有6个分枝的芦苇,但是主杆长度仅为692.0 mm。河床靠近两岸的位置河水的冲刷作用较小,因此不像河床中心的芦苇一样主杆很粗,C采样点位发现一株芦苇直径为6 mm,长度可达1 991 mm,河床靠近两岸的芦苇普遍较细。
同样的证据表现在采样断面2。采样断面2在河床低地的边缘,干涸小河流河床低地的环境类似于河床两岸,采样断面2的芦苇样本长度和直径小于采样断面1,采样断面2的河床中心也发现了带有分枝的芦苇。生长在芦苇群落边缘的芦苇没有像生长在群落中的芦苇具有向上获取更多光照的需求,而且生长在芦苇群落边缘的芦苇为了横向占领生存空间生长出了分枝,所以生长在芦苇群落边缘的芦苇比较矮且分枝多。
焦亮等[21]对敦煌阳关盐沼湿地芦苇种群小尺度空间分布格局的研究指出,轻度盐碱化区(土壤盐分2.61±0.59)、重度盐碱化区(土壤盐分3.42±0.51)对应的芦苇高度分别为(1 044.6±39.6) mm和(573.6±3.94) mm。根据研究区芦苇长度平均值为822.1 mm,可推测艾家畔村干涸小河沟的盐碱化程度可能在重度和轻度之间,但还需要更确切的数据支持。
研究区干涸小河沟芦苇是在冬季火干扰后生长起来的植物群落。王娟等[22]关于火干扰对内陆荒漠湿地芦苇群落影响的研究表明,火干扰降低了研究区植物群落的物种多样性,其中草本物种多样性减少较为明显。干涸小河沟所有采样点位,仅有采样点位Ⅲ、E分别发现3株三毛草和1株锦屏藤,其他采样点位未发现其他植物种类,符合火干扰对芦苇群落影响的结论。
由于采样条件所限,只对黄土高原艾家畔村小河沟的夏季干涸时段河道内芦苇的分布特征和环境适应性开展了相关研究,未来可补充收集芦苇地下部分和小河沟有水时段的数据,检测完整的芦苇生长周期状况,并同步补充水土方面的数据。
5 结论
黄土高原艾家畔村干涸小河沟的芦苇分布特征在数量上表现为:河床靠近两岸>河床中心,远离河床低地>河床低地边缘;有分枝的芦苇和主秆折断的芦苇在数量上表现为:芦苇群落边缘>芦苇群落中央,河床靠近两岸>河床中心,河床低地边缘>远离河床低地;芦苇主秆的长度平均值和直径的平均值表现为:河床中心>河床靠近两岸,远离河床低地>河床低地边缘。
芦苇对干涸小河沟环境的适应性表现为:为了获得光照和生存空间,芦苇在群落内展现出了种内竞争关系,越是在芦苇群落中央,芦苇在长度和直径方面表现出的长势越好。河床中心的芦苇通过长高和变粗来获得光照和抵抗干涸小河沟有水期的冲刷。河床靠近两岸地势较高和河床低地边缘的芦苇普遍较矮且分枝较多,由于处于芦苇群落边缘,没有很强的光照竞争需求,能量主要用以产生分枝来竞争横向生存空间。
猜你喜欢
语文周报·教研版(2021年28期)2021-08-19 02:14:42
装备制造技术(2021年4期)2021-08-05 07:40:24
制造技术与机床(2018年12期)2018-12-23 02:41:18
数学理论与应用(2017年2期)2017-06-27 07:38:56
数学理论与应用(2017年2期)2017-06-27 07:38:54
测绘科学与工程(2016年4期)2016-04-17 06:51:08
数学理论与应用(2016年1期)2016-02-28 09:25:55
星星·散文诗(2015年34期)2015-10-26 19:25:18
军事文摘·科学少年(2014年8期)2014-05-30 01:22:53
河南科技(2014年23期)2014-02-27 14:18:45
