
浙江南部近海前肛鳗营养生态位变化研究
——基于稳定同位素技术
2022-12-26 13:25韩东燕高春霞魏秀锦
生态学报 2022年23期
杨 蕊,韩东燕,2,高春霞,2,3,4,*,魏秀锦,赵 静,叶 深
1 上海海洋大学海洋科学学院, 上海 201306 2 国家远洋渔业工程技术研究中心, 上海 201306 3 大洋渔业资源可持续开发教育部重点实验室, 上海 201306 4 农业农村部大洋渔业资源环境科学观测实验站, 上海 201306 5 浙江省海洋水产养殖研究所,温州 325005
营养生态位是水域生态学研究的重要内容,表示生物在资源利用过程中其自身营养特征及其所处水域生态系统中的功能和位置[1],对其开展研究有利于深入解析生态系统中生物种间营养关系、物种对饵料的利用状况及食物网结构的稳定性机理等[2]。稳定同位素技术是近年来被广泛应用于示踪水域生态系统物质循环和能量流动的一种现代技术,其中的C、N稳定同位素因具有相对稳定的分馏系数被广泛用于揭示生物摄食饵料信息[3]、计算生物营养级位置[4—5]和量化营养生态位[6]。Newsome等[7]将生物组织碳氮稳定同位素比值所占据的“δ-空间”定义为同位素生态位(isotopic niche),用于描述个体、种群或群落营养生态位[8]。Varela等[9]基于同位素生态位评估了直布罗陀海峡大西洋蓝鳍金枪鱼(Thunnusthynnus)和条纹原海豚(Stenellacoeruleoalba)的营养关系,发现两种捕食者营养重叠度较低,种间饵料资源竞争小,在生态系统中发挥不同的营养作用;银利强等[10]将稳定同位素技术与多元统计分析结合,对南海中西部海域春季三种金枪鱼类的营养生态位进行了比较,表明当饵料资源有限时,种间竞争可能会影响种群数量,进而影响群落组成和生态系统功能。Layman等[11]提出δ13C值变幅(CR)、δ15N值变幅(NR)、生态位总面积(TA)等营养生态位度量指标,在量化种群营养多样性中被广泛应用,石焱等[12]根据营养生态位度量指标分析了闽江口凤鲚(Coiliamystus)等鱼类的营养生态位季节变动;黄佳兴等[13]基于碳氮稳定同位素特征值比较了中型群、微型群鸢乌贼(Sthenoteuthisoualaniensis)营养生态位的变化规律,表明不同种群的体型大小和捕食策略会对生态位宽幅产生影响。
浙江南部近海位于东海中南部,属暖温带海区[14],受沿岸流和台湾暖流影响,海域内水文条件适宜,饵料充足[15],是众多经济鱼类、虾类、蟹类的繁殖、产卵等活动场所[16]。前肛鳗(Dysommaanguillare)属暖水性中小型鳗鱼,近海底层鱼类,主要分布于我国东海,是浙江南部近海的重要种[17],在浙江南部近海食物网中属于中高营养级生物[18]。目前,中高营养级鱼类的摄食生态研究主要集中在食性、营养级和营养谱研究[19—20],并未见营养生态位方面的研究。鱼类作为食物网的重要组成部分,在食物网结构稳定性中承担着重要作用,研究其在食物网中的营养生态位分化有助于研究者从营养结构角度解析海洋生态系统的稳态机理。因此,本研究利用2016年和2020年前肛鳗肌肉样品,通过测定其碳氮稳定同位素比值,计算营养生态位度量指标,比较不同发育过程、不同季节及不同年份前肛鳗营养生态位分化变动趋势,探究前肛鳗对饵料资源利用及对生态环境的自适应能力,对调查海域食物网结构格局及稳定性机制研究具有重要的科学意义。
1 材料和方法
1.1 数据来源
本实验样品为2016年2月(冬季)、5月(春季)、8月(夏季)、11月(秋季),2020年8月(夏季)、11月(秋季)和2021年1月(冬季)在浙江南部近海(27°00′N—29°00′N,120°30′E—123°00′E)利用单船底拖作业捕获所得(图1)。调查船为浙洞渔10109号,马力为540、吨位为305 t,调查方式为底拖网作业,网口宽40 m,高7.5 m,底纲和浮子纲为80 m,拖速为2—4 kn,每站拖网时间1 h左右。样品个体经冷冻保存带回实验室分析。
图1 浙江南部近海调查站点Fig.1 Survey stations in offshore waters of Southern Zhejiang站点WT为温州市与台州市缩写
1.2 样品处理
本研究选取71尾前肛鳗样品(表1),测量包括全长(吻端至尾部末端)、肛长(吻端至肛门)及体质量等生物学性状,其中,全长及肛长测量结果精确至0.1 cm,体质量测量结果精确至0.1 g。此次调查前肛鳗的肛长范围为52—89 mm,为探究前肛鳗营养生态位随个体发育的变化,以10 mm为间隔将前肛鳗样品按肛长分组,分别对应为LG1、LG2、LG3、LG4(表1)。采集每个肛长组样品的背部肌肉组织,肌肉样品经超纯水清洗后置于-55℃的冷冻干燥机(Christ 1- 4α)中干燥24 h,使用混合型球磨仪(Retsch MM400)研磨成粉末,过100目筛。

表1 浙江南部近海前肛鳗样品信息
1.3 稳定同位素比值测定
研磨后的前肛鳗样品取1.5 mg粉末包埋于锡舟(型号4 mm×4 mm×11 mm)中,利用元素分析-稳定同位素比质谱仪(ISOPRIME 100,Isoprime Corporation, Cheadle, UK)测定样品的碳、氮稳定同位素比值(δ13C、δ15N)。
测定结果以碳稳定同位素比值(δ13C)和氮稳定同位素比值(δ15N)表示,计算公式如下:
δX=[(Rsample/Rstandard)-1] ×103
式中,X为13C或15N;R为13C /12C 或者15N/14N的丰度比值。
样品稳定同位素测定在上海海洋大学大洋渔业资源可持续开发教育部重点实验室分析测试中心进行。为保证测量样品的准确性,将国际通用的美国拟箭石(PDB)和大气中的氮气(N2-atm)分别作为碳、氮稳定同位素测定的标准物质,并采用USGS 24和USGS 26对同位素仪器进行校正,每10个待测样品放入3个实验室Ⅱ级蛋白质标准品(Protein:δ13C=-26.98‰,δ15N=5.94‰)进行校准,仪器测定精度为δ13C<±0.2‰,δ15N<±0.3‰。
考虑到肌肉脂质含量对样品中碳稳定同位素及脱脂试剂对样品氮同位素的影响,本文基于杨蕊等[21]提出的浙江南部近海前肛鳗脂质校正公式对本研究中的前肛鳗样品δ13C值进行校正。
1.4 营养生态位指标
Layman等[11]基于以碳氮稳定同位素为横纵坐标的双位图,提出6个量化种群营养生态位的指标,其中4个营养多样性指标:δ13C范围(δ13C range, CR)表示碳同位素最大值与最小值的差值(δ13Cmax-δ13Cmin),表征食源多样性;δ15N 范围(δ15N range, NR)表示氮同位素最大值与最小值的差值(δ15Nmax-δ15Nmin),表征物种的营养多元化程度;生态位总面积(total area, TA)表示在碳氮稳定同位素双位图中,物种的所有个体代表的坐标点组成的凸多边形面积,表征物种占据的营养生态位空间总量;平均离心距离(mean distance to centroid, CD)表示每一个物种所代表的坐标点到碳氮稳定同位素双位图重心的平均欧氏距离,重心代表群落中所有物种的δ13C和 δ15N 的平均值,表征物种营养多样性;2个营养冗余指标:平均最近相邻距离(mean nearest neighbor distance, MNND)表示在碳氮稳定同位素双位图中每个物种所代表的坐标点与其最近的相邻物种的坐标点的平均欧氏距离,表征营养密度,是度量营养相似性的指标;最近相邻距离的标准差(standard deviation of nearest neighbor distance, SDNND)表示碳氮稳定同位素双位图中个体与其最近相邻坐标点欧氏距离标准偏差,衡量种群营养均匀度。
指标中TA受样品大小和双位图中δ13C轴或δ15N轴极端值影响,因此本研究利用Jackson等[22]提出的校正标准椭圆面积(corrected standard ellipse area, SEAc)代替TA指标,SEAc是对核心同位素生态位面积的校正,是衡量营养生态位的重要指标,常用此指标量化物种的营养生态位宽度,估算个体间营养生态位重叠面积,公式如下:
SEAc=SEA(n-1)×(n-2)
式中,SEAc为校正标准椭圆面积,SEA为标准椭圆面积,n为样品数。
本研究中将两个体间的核心同位素生态位重叠面积定义为OA(overlap area),OA则作为衡量两个体间营养生态位变化和营养生态位重叠的度量指标[23—24]。
1.5 数据处理与分析
开展统计分析前,利用Kolmogorov-Smirnov检验对样品碳、氮稳定同位素数据开展正态分布检验,若检验数据符合正态分布和方差齐性,则采用单因素方差分析(one-way ANOVA)检验前肛鳗不同发育过程及不同季节间δ13C、δ15N是否存在差异性,显著水平为P<0.05;数据处理在Excel 2019和SPSS 24软件中开展。利用R4.0.5中的SIBER软件包计算5项营养生态位度量指标和校正标准椭圆面积(SEAc)及重叠面积。
2 结果与分析
2.1 碳、氮稳定同位素比值
2016年浙江南部近海前肛鳗δ13C值范围为-16.17‰—-14.73‰,平均值为(-15.19±0.31)‰,最大差值为1.44‰;δ15N值范围为11.56‰—13.40‰,平均值为(12.42±0.45)‰,最大差值为1.84‰(图2)。利用单因素方差分析发现前肛鳗不同发育过程δ13C值差异不显著(P>0.05),δ15N值差异显著(P<0.05);不同季节间δ13C值存在极显著差异(P<0.01),δ15N值差异显著(P<0.05)。

图2 浙江南部近海不同发育过程前肛鳗的δ13C和δ15N值双位图Fig.2 The biplots of δ13C and δ15N values of the different developmental stages of Dysomma anguillare from the offshore waters of Southern Zhejiang
2020年浙江南部近海前肛鳗δ13C值范围为-16.76‰—-14.82‰,平均值为(-15.90±0.45)‰,最大差值为1.94‰;δ15N值范围为12.29‰—13.36‰,平均值为(12.92±0.25)‰,最大差值为1.07‰(图3)。利用单因素方差分析发现前肛鳗不同发育过程δ13C值差异不显著(P>0.05),δ15N值差异不显著(P>0.05);不同季节间δ13C值存在显著差异(P<0.05),δ15N值差异不显著(P>0.05)。
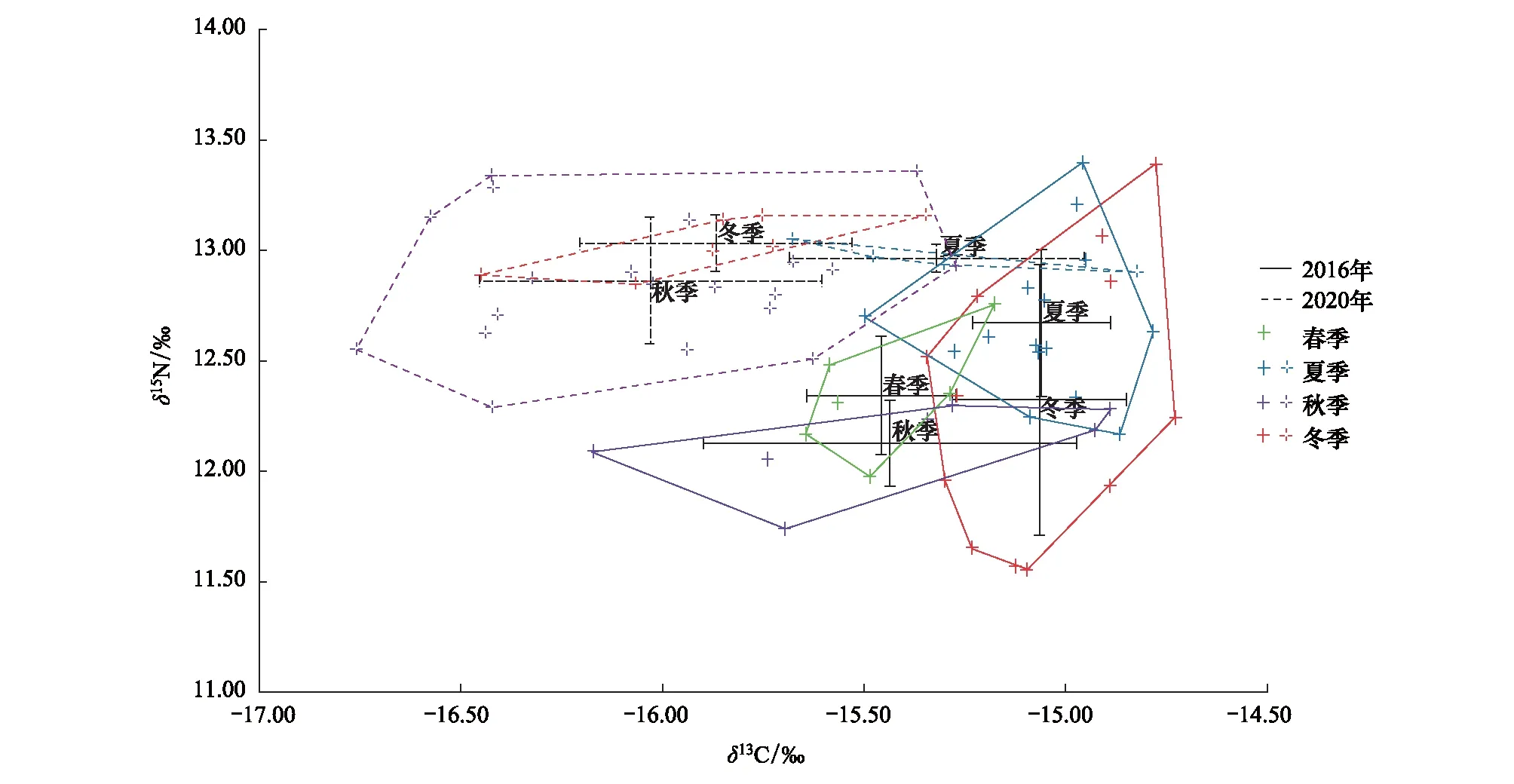
图3 浙江南部近海不同季节前肛鳗的δ13C和δ15N值双位图Fig.3 The biplots of δ13C and δ15N values in different seasons of Dysomma anguillare from the offshore waters of Southern Zhejiang
2.2 营养生态位特征的个体发育变化
分析2016年4个肛长组的营养生态位量化指标(表2),其中CR指标结果显示LG4大肛长组的食源多样性低于其他组;NR指标结果表征大个体组的营养多元化程度整体要略高于小个体组;CD指标在体长组间并无明显差异,LG1和LG2个体组略高于另外两组;SEAc指标表示营养生态位宽幅,其值随个体发育呈降低趋势;营养冗余指标MNND和SDNND在各肛长组间变动趋势不明显,仅见LG4大个体组的SDNND略小于其他组。
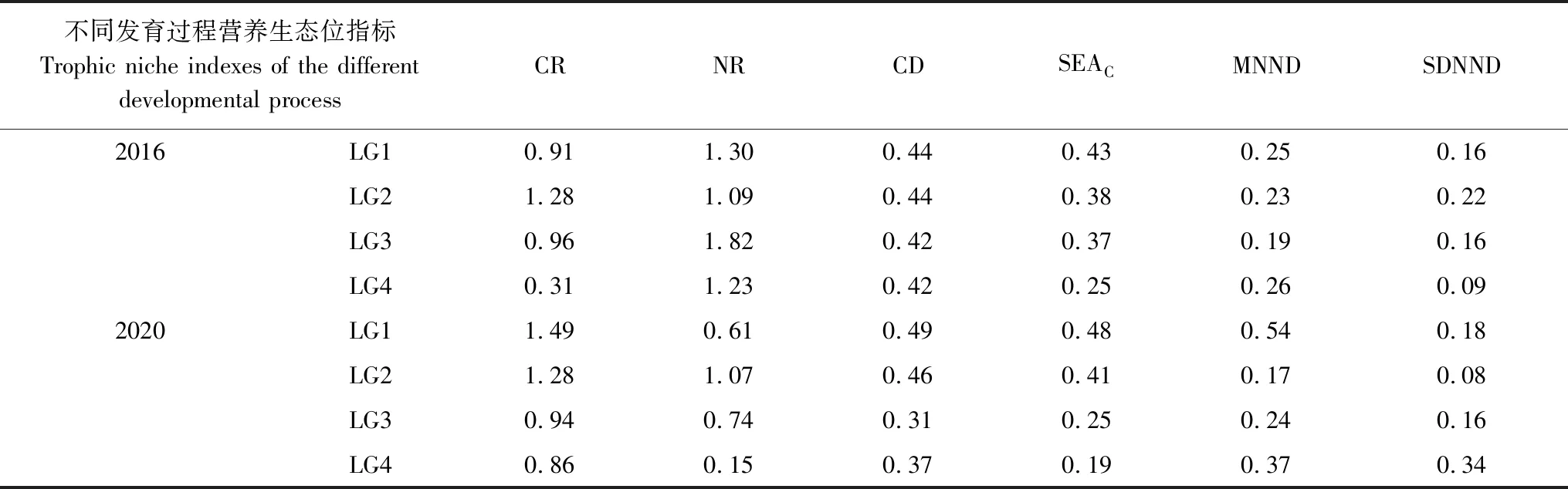
表2 浙江南部近海前肛鳗不同发育过程营养生态位指标
分析2020年4个肛长组的营养生态位量化指标,其中CR指标结果显示随着个体发育食源多样性呈下降趋势,大个体组的食源多样性要更低;NR指标结果表征大个体组的营养多元化程度整体要略低于小个体组;CD指标在LG1和LG2个体组中略高于另外两组;SEAc指标结果随个体发育呈下降趋势;营养冗余指标MNND和SDNND在各体长组间变动趋势不明显,仅见LG1和LG2中小个体组的SDNND要低于大个体组。
对比2016年和2020年营养生态位随个体发育的变化,从食源多样性来看,两年的CR值相似,且变动趋势也基本相似,大个体组的食源多样性要小于小个体组;从营养多元化程度来看,2020年前肛鳗的营养多元化程度要小于2016年,随个体发育的变动趋势相反;从生态位宽幅来看,两年的SEAc值相似,且均随个体发育呈降低趋势;从营养冗余来看,两年的冗余指标值无差异,且无明显随个体发育的变动趋势,单因素方差分析结果表明两年份除NR指标为显著性外(P<0.05),其余指标均无显著性差异(P>0.05)。
表3、图4为2016年和2020年前肛鳗不同发育过程的营养生态位重叠情况,2016年前肛鳗LG1和LG4个体组营养生态位重叠面积最小,仅0.04;其次为LG2和LG4个体组营养生态位重叠面积为0.09;LG2和LG3个体组营养生态位重叠面积最大,达到0.27。2020年前肛鳗LG3和LG4个体组营养生态位重叠面积最小,为0.04,其次为LG2和LG4个体组,营养生态位重叠面积为0.07;其次为LG2和LG3个体组营养生态位重叠面积为0.23;LG1和LG2个体组营养生态位重叠面积最大,为0.31。
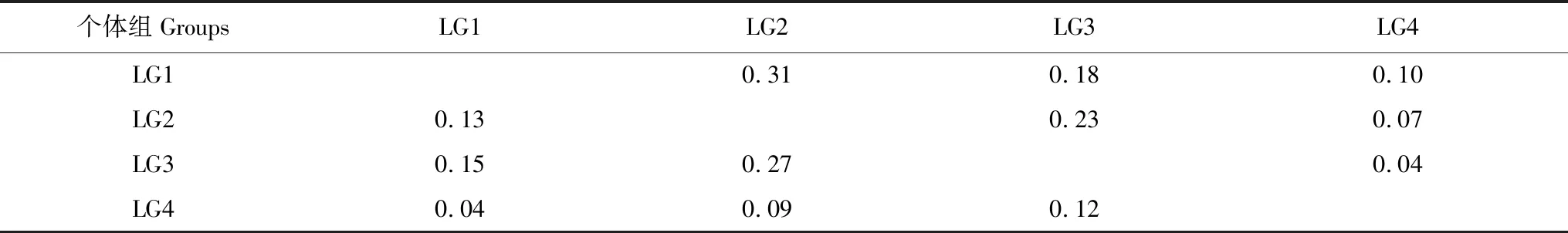
表3 浙江南部近海前肛鳗不同发育过程营养生态位重叠面积(OA)
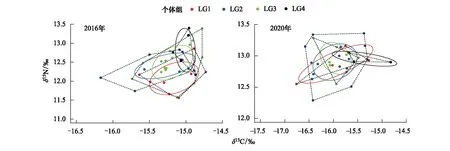
图4 2016年和2020年浙江南部近海前肛鳗不同发育过程的营养生态位变动Fig.4 Trophic niche changes of the different developmental process of Dysomma anguillare from the offshore waters of Southern Zhejiang in 2016 and 2020
2.3 营养生态位宽幅的时间变化
2.3.1 营养生态位宽幅的季节变化
分析2016年4个季节的营养生态位量化指标(表4),其中CR指标结果显示秋季的食源多样性要高于其他季节;NR指标结果表征夏季和冬季营养多元化程度相近,春季和秋季营养多元化程度相近,且夏、冬季营养多元化程度高于春、秋季;SEAc指标显示前肛鳗营养生态位宽幅随季节变化有升高趋势,其中冬季所占据的营养生态位空间最大,其次为秋季,春季最小;营养冗余指标MNND和SDNND在各季节组间并无明显差异。
分析2020年3个季节的营养生态位量化指标,其中CR指标结果显示秋季的食源多样性要高于其他季节;NR指标结果表征夏季和冬季营养多元化程度相近,但其营养多元化程度远低于秋季;SEAc指标显示2个季节中秋季营养生态位宽幅最大,夏季最小;营养冗余指标MNND和SDNND在各季节间无明显差异。
对比2016年和2020年前肛鳗营养生态位随季节的变化趋势,从食源多样性来看,秋季CR值普遍高于其他季节,两年的季节变动趋势基本相似;从营养多元化程度来看,2020年前肛鳗在各季节的营养多元化程度较2016年普遍下降;从营养生态位宽幅来看,2016年前肛鳗在秋、冬季拥有更高的生态空间利用率,2020年前肛鳗在秋季具有较高的SEAc,整体上 2020年前肛鳗的各季节营养生态位宽幅较2016年要更小;从营养冗余程度来看,这两年前肛鳗的营养冗余随季节变化无明显变动趋势,单因素方差分析结果表明各指标均无显著性差异(P>0.05)。

表4 浙江南部近海前肛鳗不同季节营养生态位指标
表5和图5为2016年和2020年前肛鳗不同季节的营养生态位重叠情况,2016年前肛鳗春-夏季不存在营养生态位重叠现象(OA=0),而夏-冬季营养生态位重叠面积最大,为0.16;春-冬季、夏-秋季营养生态位重叠面积均小于0.01。2020年夏-冬季营养生态位重叠面积最小,其重叠面积为0.003;秋-冬季营养生态位重叠面积最大,为0.08。

表5 浙江南部近海前肛鳗不同季节营养生态位重叠面积(OA)

图5 2016年和2020年浙江南部近海前肛鳗不同季节的营养生态位Fig.5 Trophic niche changes of the different seasons of Dysomma anguillare from the offshore waters of Southern Zhejiang in 2016 and 2020
2.3.2 营养生态位宽幅的年际变化
对比分析2016年和2020年整体营养生态位的变化情况(表4、图6),发现4个营养多样性指标中,除CR指标有所升高外,其余3个指标均有不同程度的降低;2个营养冗余指标显示2020年MNND和SDNND指标均略低于2016年。从营养生态位变化图可以看出,2020年前肛鳗在食物网中占据的营养位置要略高于2016年,两年的SEAc未出现重叠现象(OA=0)。
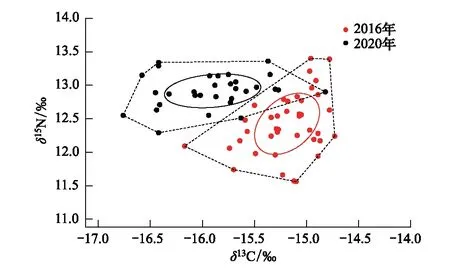
图6 浙江南部近海不同年际间前肛鳗的营养生态位 Fig.6 Trophic niche of Dysomma anguillare in different yaers from the offshore waters of Southern Zhejiang
3 讨论
3.1 营养生态位个体发育差异分析
鱼类在生长过程中因口器等摄食器官的发育,摄食饵料生物的种类和强度也会随之改变[25],这种食源变化则可能导致生物在其不同发育过程碳氮稳定同位素特征存在差异[26]。本研究利用碳氮稳定同位素技术分析前肛鳗不同发育过程δ13C、δ15N值,发现2016年和2020年前肛鳗δ13C值在个体发育间均无显著差异(P>0.05),表明前肛鳗在2016年和2020年的个体发育过程中均未伴随明显的食源转变现象。然而,研究发现2016年的前肛鳗不同体长组平均δ15N值随个体发育呈现增高趋势,单因素方差分析结果显示前肛鳗不同发育过程δ15N值存在显著差异(P<0.05),这表征前肛鳗有向高营养级饵料生物摄食的趋势。前肛鳗是底栖肉食性种类,出现这种现象与薛莹等[27]在南黄海皮氏叫姑鱼(Johniusbelengerii)中的研究发现一致,即随着个体发育,鱼类的营养来源未发生明显变化,但其小个体时食源以多种低营养级的底栖生物为主,随着体长的增加,成体摄食的优势饵料生物逐渐转变为大型底栖甲壳类,从而出现这种食源保持底栖不变但摄食种类占比出现变化的现象。这种摄食特性在2020年的前肛鳗中却并无明显表现,研究发现2020年前肛鳗LG1—LG3个体组平均δ15N值随个体呈现逐渐增高趋势,但是比值差异较小,不同发育过程前肛鳗的δ15N值差异不显著(P>0.05),造成这种结果差异的原因可能与2020年前肛鳗的食源多样性(CR=1.94)较高有关,在饵料充足的情况下,不同发育过程的捕食者都会偏向捕食利盈较大的种类,以通过提高自身营养级水平,避免与其他中、底营养级鱼类产生生态位竞争。
生态位宽幅是生物所能利用的各种资源的总和,是表征物种或种群适应环境和利用资源的实际幅度或潜在能力指标[28]。Layman等[11]提出的六项量化稳定同位素的指标,从不同角度度量种群营养生态位的大小,其中TA受样品量大小影响,常采用Jackson等[22]提出的小样品量校正后的标准椭圆面积(SEAC)代替TA指标表征物种营养生态位的总宽幅,SEAC对样品量大小的敏感度降低,代表物种占据的核心同位素生态位空间。目前有关物种营养生态位的较多研究多关注于评估物种种间差异[13, 29]、营养生态位的季节变化和营养群落营养结构[12, 30],较少考虑营养生态位随个体发育的变化情况。Heuvel等人[31]基于3种稳定同位素对伊利湖大眼狮鲈(Sandervitreus)等3种鱼类的摄食生态研究中发现鱼类营养生态位面积随个体大小的增加而显著减小。本研究同样发现前肛鳗随肛长增大其校正标准椭圆面积逐渐减小,小个体前肛鳗营养位置较低,营养生态位宽幅较大,且大个体组前肛鳗与其他肛长组的营养生态位重叠面积均小于小个体组,究其原因可能由于在鱼类生长发育过程常伴随饵料资源竞争,生物个体发育过程中为满足自身生长发育所需能量,趋于增强对饵料生物的捕食能力,营养生态位也会随之发生偏移以减小种间和种内竞争,小个体前肛鳗在相同资源环境条件下提高自身对饵料资源的利用能力,增大生态位宽幅以保证自身存活。
3.2 营养生态位的季节差异分析
碳氮稳定同位素特征可指示生物体与其饵料之间的捕食关系及营养富集趋势,δ13C范围(CR)用于表征食源的多样性,δ15N范围(NR)用于表征营养多元化程度。盖珊珊等[32]以小黑山岛人工鱼礁区许氏平鮋(Sebastesschlegelii)和大泷六线鱼(Hexagrammosotakii)为研究对象,利用碳、氮稳定同位素技术分析了上述两种鱼类的营养生态位宽幅变化及重叠情况,表明物种营养生态位存在季节变化。本研究通过ANOVA检验发现2016年和2020年不同季节前肛鳗δ13C值均存在显著差异(P<0.05),表明不同季节前肛鳗食源可能不同,食源分布受季节因素影响较大,王跃中等[33]对东海带鱼资源变动影响因子研究中表明水温对鱼类的天然饵料影响较大,水温的变化通过影响鱼类代谢强度、体温等进而直接影响鱼类摄食和生长;沙永翠等[34]对太湖贡湖湾高营养鱼类黄颡鱼(Pelteobagrusfulvidraco)营养生态位的研究中认为营养生态位主要受食物资源丰富度、可利用性及生物间相互作用的影响。本研究中不同季节的食源差异同样引起前肛鳗营养生态位的变动,计算营养生态位CR指标发现2016年和2020年春、夏两季δ13C范围普遍小于秋、冬两季,表明前肛鳗秋、冬两季食物来源较春、夏两季广泛,结合本研究SEAC指标结果来看,浙江南部近海前肛鳗春、夏两季SEAC指标结果小于秋、冬两季。究其原因,浙江南部近海春、夏季水温逐渐升高,可供捕食者摄食的饵料生物种类和数量增多,供给充足,捕食者将优先选择偏爱的饵料生物或处于相对高营养位置的饵料生物进行摄食活动,进而导致前肛鳗春、夏两季食源多样性下降,营养生态位总宽幅较小;而秋、冬两季近岸水域水温降低,饵料生物种类和数量下降,食源营养层次减少,鱼类通过扩大摄食范围以减少种内竞争,因此秋、冬两季δ13C值范围宽,食源多样性增加,营养生态位总宽幅较春、夏两季更高。此外,本研究中前肛鳗在各季节间营养生态位重叠面积较小,如2016年春季与其他季节仅存在极小重叠(OA≤0.06),营养生态位存在明显的时间分化,而个别季节间存在一定的生态位重叠现象,如2016年夏季与冬季的存在较高的营养生态位重叠,但至2020年发现夏季与冬季营养生态位重叠面积反而最小,这其实证明了前肛鳗的营养生态位随时间呈现动态变化,这也是物种通过自身调节方式降低与其他物种的种间竞争,维持种间稳定共存的一种机动性策略。
3.3 营养生态位的年际差异分析
生态位测度指标是影响物种种群结构及多样性的重要因素,表征物种在群落或生态系统中的功能位置,生态位面积的变化可反映出物种对资源利用的变化情况[31]、物种种群的稳定性及物种间潜在的竞争关系[7, 35]。本研究2016年前肛鳗δ13C值范围为-16.17‰—-14.73‰,δ15N值范围为11.56‰—13.40‰;2020年前肛鳗δ13C值范围为-16.76‰—-14.82‰,δ15N值范围为12.29‰—13.36‰,两个年份的变动范围接近,这表明2016年和2020年前肛鳗的食性在年际间可能并没有发生明显变化,而且它的δ13C值与浙江中南部底栖生物的δ13C值(-16.71‰—-14.58‰)一致,而δ15N值则高1—1.5个营养级左右(8.79‰—10.89‰)[30],表明前肛鳗主要以底栖动物为食,这个结果与张波等[36]基于胃含物分析方法得出的前肛鳗主要摄食对象为底栖虾蟹类的结论接近,也与李忠义等[18, 37]学者得出前肛鳗为完全底栖生物食性这一结论基本吻合。但是,我们也注意到,2016年δ13C值的主要集中范围和平均值均富于2020年,因此推测两年前肛鳗种群食性偏好发生一定的变化,平均δ15N值的年际差异为0.5‰,表明前肛鳗种群在浙江南部近海水域生态系统中营养级有所升高但所处的营养层次未出现明显的变化。
种群营养结构的指标CR、NR、SEAc、CD常用来表征种群营养结构的多样性,MNND、SDNND常用来量化种群冗余程度,种群中具有相似营养特征的个体占大多数时,MNND值越小,种群的营养冗余程度越高。本研究中,2020年与2016年相比较,除CR指标外,其余指标均出现不等程度的下降,从年际间营养生态位变化图(图6)可以看出,2020年浙江南部近海前肛鳗营养生态位总宽幅明显小于2016年,且未出现营养生态位重叠现象(OA=0),说明浙江南部近海前肛鳗营养生态位发生明显变化,种群营养结构多样性降低,对海域内环境的适应能力及对饵料资源的利用能力均有所减弱。结合冗余指标,2020年前肛鳗的种群结构冗余程度略低于2016年,种群内相似营养生态位个体及种群结构稳定性略低于2016年,出现此结果可能与浙江南部近海渔业资源的捕捞强度变动有关,根据《农业部部关于加快推进渔业转方式调结构的指导意见》(农渔发[2016]1号)文件[38],近海捕捞渔船数量已经迅速减少[39],海域内捕捞压力的变化会通过自上而下的级联效应对饵料生物营养群落结构产生影响。此外,王跃中等[33]表示气候变化不仅直接影响鱼类的生存环境,还可通过食物链的关系影响到鱼类的饵料生物量,本研究本底环境调查结果表明2016年浙江南部近海年平均水温为20.59℃,2020年浙江南部近海年平均水温较2016年升高1.30℃,樊伟等[40]对全球环境变化对渔业资源的影响研究中表明水温升高会使生物活动增强,鱼类生长所需要的饵料有机物增多,因此推断气候变动导致的海域内可供摄食的饵料生物变化也可能是造成前肛鳗营养生态位发生偏移的原因。
猜你喜欢
文萃报·周五版(2022年15期)2022-04-21
读者(2022年8期)2022-04-07
快乐作文(1.2年级)(2019年3期)2019-09-10
英语文摘(2019年5期)2019-07-13
幼儿画刊(2018年10期)2018-10-27
同位素(2018年1期)2018-01-18
扬子江诗刊(2016年2期)2016-11-25
小雪花·成长指南(2016年2期)2016-03-16
新少年(2015年5期)2015-07-10
同位素(2014年3期)2014-06-13
