
臭柏异形叶空间分异及光合生理特性研究
2022-12-24 05:54:38邹海冬韩阳媚欧阳嗣航刘叶凡陈海鹏李玉灵
林业与生态科学 2022年4期
邹海冬,韩阳媚,欧阳嗣航,刘叶凡,陈海鹏,李玉灵
(河北农业大学 林学院,河北 保定 071000)
光照可以通过光合作用影响植物的生长发育。然而,对于植物而言,光照既是必不可少的资源,又易对植物造成胁迫。如对于耐阴树种而言,植物间的遮阴可以促进其生长[1];而对于喜光树种,弱光条件下的光照作为有限的资源,可能引起植物间的竞争[2]。但是,过量的光照会引起包括光系统Ⅱ(PSⅡ)在内的植物细胞组织损伤,进而降低光合作用的效率,而植物也会通过多种措施减少光胁迫对植物产生的影响[3]。从形态结构上,植物可以通过叶形差异或叶片倾角不同,避免吸收过多的光照,进而提高对光照资源的合理利用,如胡杨垂直冠层自上而下分别为阔叶、卵圆形和披针形,及冠层中部出现少量的肾性叶和菱形叶等过渡叶,这种同一株植物上生长不同形状叶片的现象,称为异形叶性(Heterophylly)[4-8]。此外,植物也会通过生理活动减少光胁迫对植物带来的不利影响。当植物吸收过多的光照时,可以通过叶黄质循环等光保护机制,将光照以热量的形式耗散出去[9]。因此,探明光照在植物叶片异型分化中的作用及植物对光环境的响应情况,对更好的调控植物生长具有重要意义。
臭柏,学名叉子圆柏(Sabinavulgaris),柏科(Cupressaceae)刺柏属(Juniperus)常绿匍匐状小灌木,其冠层高度一般不超过1.5 m,因其生命力极强、适生范围广泛、生态功能良好,对于我国北方干旱和半干旱地区的植被恢复、防风固沙、园林绿化等均有重要作用[10-12]。但是,不同发育阶段臭柏对于光照条件的适应性有所差异。天然更新下的幼龄臭柏必须依靠乌柳等灌丛形成的遮阴环境才能正常生长,而成年臭柏则因完全适应强光环境而取代“保姆植物”,形成灌木纯林,其叶形也从以刺叶为主转变为以鳞叶为主,为典型异形叶性植物[11-13]。因此,基于本课题组前期研究结果,通过臭柏盆栽试验,研究10%和100%光照环境下1年生和3年生臭柏典型鳞叶和典型刺叶空间分异特征和光合生理特性,为异形叶分异及其对光环境适应性提供理论依据[12,14-15]。
1 材料与方法
试验地点为河北农业大学实验农场第三分厂(E 115°24′33″,N 38°48′20″),位于保定市南部地区,该地区属于暖温带大陆季风气候区。
1.1 试验材料
试验苗木为臭柏扦插苗,由河北农业大学清西陵教学林场臭柏苗木繁育圃提供。
1.2 试验设计
2021年4月29日,1年生臭柏和3年生臭柏扦插苗分别盆栽100株。本试验所用土壤为田间壤土。花盆容量为21 L,半埋入土中。臭柏株行距为50 cm×80 cm,露天栽植。
2021年5月29日,臭柏缓苗满1月后,开始进行遮光处理。基于前期研究结果[12,14-15],按照两因子完全随机区组试验设计,将1年生和3年生臭柏扦插苗雄株作为2个苗龄组,用黑色塑料遮阳网设置透光率10%光照(遮光)处理,100%光照(全光)处理,则不进行遮光处理。每个苗龄、每个处理50盆。
1.3 测定方法
1.3.1 臭柏异形叶空间分异 每个处理选择15株长势健康的臭柏,每株臭柏冠层分为上、中、下3个部位。自9月起,每7 天记录1次臭柏1级侧枝上典型刺叶小枝和典型鳞叶小枝的数量(后文均简称为鳞叶和刺叶,且不区分不同大小和叶片倾角的刺叶,统记为刺叶),每个部位记录5枝1级侧枝。
1.3.2 臭柏异形叶光合生理特性 2021年9月,选择晴朗无风的天气进行臭柏光合生理参数测定。每个处理下选取长势健康且具有代表性3株臭柏,使用Li-6800便携式光合仪(Li-Cor,Lincoln,NE,美国)对臭柏异形叶进行光合指标的测定。分别选取1年生臭柏冠层下部的鳞叶和上部的刺叶,3年生臭柏冠层上部的鳞叶和下部的刺叶进行测量。
测定光合特性包括净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs),测量时间为8:00-18:00,每2 h测定1次,重复数为3,并计算水分利用效率(WUE=Pn/Tr)和气孔限制值[Ls=(Ca-Ci)/Ca),Ca为大气CO2浓度][16]。测定光合特性同时记录部分叶绿素荧光参数,包括PSⅡ实际光化学效率[Fv′/Fm′=(Fm′-Fo′)/Fm′]、电子传递效率(ETR)、光化学淬灭系数(qP)、非光化学淬灭系数(NPQ)。光合生理测定结束后,用锡纸将臭柏叶包裹住,进行30 min暗适应,随后测定暗适应下最小初始荧光(Fo)和最大荧光(Fm),并计算暗适应下光系统Ⅱ(PSⅡ)最大光化学效率[Fv/Fm=(Fm-Fo)/Fm]和PSⅡ潜在活性[Fv/Fo=(Fm-Fo)/Fo][17-18]。
1.4 统计分析
使用Excel 2016进行数据整理。使用SPSS 24.0进行多因素方差分析,其中正态性检验采用Shapiro-Wilk方法,方差齐性检验采用Levin等同性检验,采用一般线性模型的单变量功能检验主效应和交互效应,对于主效应和交互效应显著(P<0.05),继续简单效应的检验;使用Origin 8.6软件进行绘图。
2 结果分析
2.1 空气温度和不同光照环境下光合有效辐射变化
试验期间空气温度和光合有效辐射日变化,见图1。
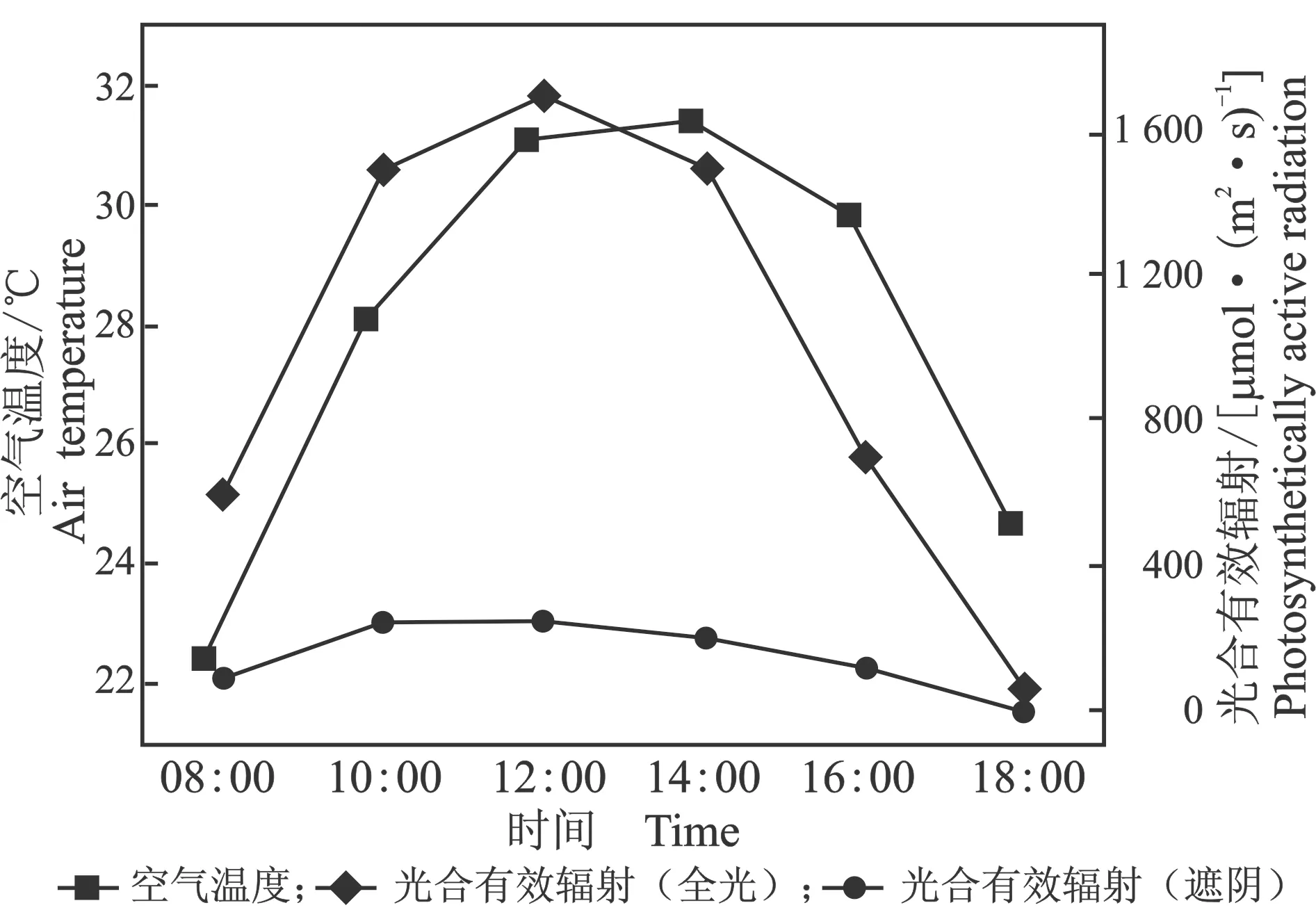
图1 空气温度和光合有效辐射日变化
由图1可知,光合有效辐射和空气温度日变化均为单峰曲线。全光环境下光合有效辐射峰值出现在12:00,达到1 700 μmol/(m2·s),遮阴环境的光合有效辐射为全光环境0~20%之间,遮阴处理符合预期效果;空气温度峰值出现在14:00,达到31.42 ℃,其峰值出现时间晚于光合有效辐射峰值出现的时间。
2.2 不同光照环境下不同苗龄臭柏异形叶空间分异特征
不同光照环境下不同苗龄臭柏异形叶空间分异特征,见表1。臭柏鳞刺比的主体间效应检验,见表2。
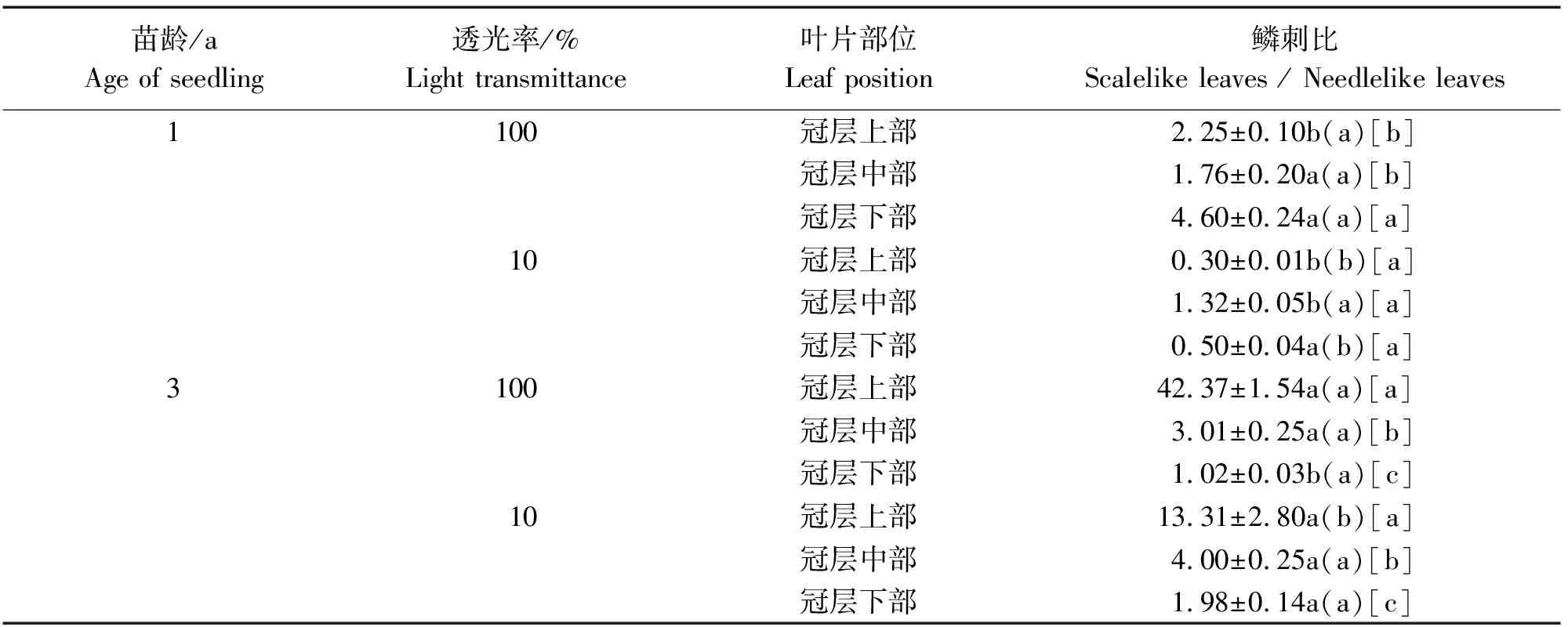
表1 不同光照环境下不同苗龄臭柏异形叶空间分异特征

表2 臭柏鳞刺比的主体间效应检验
由表1可知,1年生臭柏仅全光环境下冠层下部鳞刺比显著高于其冠层上、中部,而遮光处理显著降低了1年生臭柏冠层上、下部的鳞刺比,因此遮光处理对1年生臭柏异形叶分异的影响高于叶片部位对鳞刺比的影响。全光和遮光环境下,3年生臭柏冠层自上而下鳞刺比均显著减少,但仅遮光环境下3年生臭柏冠层上部显著低于全光环境下3年生臭柏冠层上部,因此叶片所处部位对3年生臭柏异形叶分异的影响高于遮光处理的影响。而除全光环境下3年生臭柏冠层下部鳞刺比显著低于全光环境下1年生臭柏冠层下部鳞刺比外,相同光照环境相同冠层下其余处理均3年生臭柏鳞刺比高于1年生。因此,1年生臭柏处于异形叶分异的关键时期,而3年生臭柏完成异形叶分异过程,以鳞叶为优势叶片,且叶片所处冠层位置成为主要影响因素。
由表2可知,苗龄、透光率和叶片部位主效应均对鳞刺比有显著影响,且苗龄、透光率和叶片部位两两之间的交互效应及三者间的交互效应均达到显著水平。因此本次试验中,苗龄、透光率和叶片部位的主效应及其交互效应均能影响臭柏异形叶空间分异。
2.3 不同光照环境下不同苗龄臭柏异形叶光合特性
不同光照环境下不同苗龄臭柏异形叶净光合速率和气孔限制值日变化,见图2。
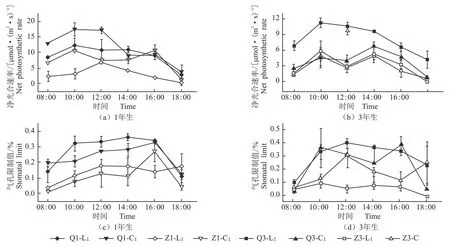
图2 不同光照条件下不同苗龄臭柏异形叶净光合速率和气孔限制值日进程曲线
由图2-a所知,100%光照环境下1年生臭柏鳞叶和刺叶和10%光照环境下1年生臭柏刺叶净光合速率(Pn)均为双峰曲线,即出现“午休”现象,而由图2-c可知,100%光照环境下,1年生臭柏鳞叶和刺叶“午休”时相应的气孔限制值(Ls)较高,因此,全光环境下1年生臭柏刺叶和鳞叶“午休”现象出现与气孔限制有关;而遮阴处理下,1年生臭柏刺叶“午休”时相应的Ls较低,因此,导致1年生臭柏刺叶出现“午休”现象的原因不是气孔限制。而100%光照环境下3年生臭柏刺叶Pn也出现了“午休”现象(图2-b),此时相应的Ls较高(图2-d),因此气孔导度降低引起了全光环境下3年生臭柏刺叶“午休”现象的出现;虽然10%光照环境下3年生臭柏鳞叶Pn出现“午休”现象(图2-b),但Ls日变化不显著(图2-d),因此,气孔导度不是10%光照环境下3年生臭柏鳞叶Pn出现“午休”现象的原因(图2-b)。此外,1年生鳞叶在全光环境下出现“午休”现象,在遮光环境下未出现“午休”现象(图2-a)。而3年生臭柏鳞叶在遮光环境下出现“午休”现象,在全光环境下未出现“午休”现象(图2-b)。因此,同为鳞叶,但是随着苗龄的改变,臭柏鳞叶对不同光环境的响应出现了差异;而仅遮光环境下3年生臭柏刺叶未出现“午休”现象,其余臭柏刺叶处理均出现“午休”现象(图2-a,2-b)。另外,10%光照环境下1年生臭柏鳞叶和刺叶的Pn均较低,而3年生臭柏鳞叶Pn因遮光处理而降低,3年生臭柏针叶在10%和100%光照环境下Pn仅在12:00有所差距(图2-a,2-b)。因此,臭柏刺叶也随着苗龄的增加而对光环境的响应有所差异。
不同光照环境下臭柏异形叶光合生理参数日均值,见表3。臭柏异形叶光合特性主体间效应检验,见表4。

表3 不同光照环境下不同苗龄臭柏异形叶光合生理特性

表4 臭柏异形叶光合特性主体间效应检验
由表3可知,在全光和遮光环境下,1年生臭柏刺叶净光合速率(Pn)显著高于鳞叶,此外,遮光环境下刺叶胞间CO2浓度(Ci)、水分利用效率(WUE)显著高于鳞叶(P<0.05)。而遮光处理显著降低了1年生臭柏鳞叶的Pn、WUE、气孔限制值(Ls),提高了其Ci;遮光处理还显著降低了1年生臭柏刺叶的Pn、蒸腾速率(Tr)。由此,遮光和叶形虽然对1年生臭柏叶片光合生理存在交互作用,但遮光对1年生臭柏叶片光合生理特性影响比叶形更显著,且遮光处理对1年生臭柏鳞叶光合生理特性影响大于刺叶。全光环境下,3年生臭柏鳞叶Pn和WUE显著高于刺叶,遮光环境下,3年生臭柏鳞叶Tr、Ci和Ls均有显著差异。而遮光处理仅显著增加了3年生臭柏刺叶的Ci,对其余3年生臭柏刺叶光合生理指标影响不显著,对除气孔导度(Gs)外其余3年生臭柏鳞叶光合生理指标影响均显著。因此,叶形对3年生臭柏叶片光合生理影响比遮光处理更显著。
由表4可知,本次试验叶形和苗龄单因素仅对Pn和WUE存在显著影响,透光率单因素除对Gs无显著影响外,对其余光合生理参数均有显著影响(P<0.05)。叶形和苗龄双因素交互作用对臭柏异形叶Pn、Ci、WUE有显著影响,叶形和透光率双因素交互作用对臭柏异形叶Pn、WUE、Ls有显著影响,而苗龄和透光率双因素交互作用仅对Pn有显著影响。叶形、苗龄、透光率三者间对除Gs外的光合生理指标均有显著交互作用。因此研究臭柏异形叶光合特性差异时,应兼顾光照环境主效应及其与苗龄、叶形的交互效应。
2.4 不同光照环境下不同苗龄臭柏异形叶叶绿素荧光参数
不同光照环境下不同苗龄臭柏异形叶叶绿素荧光参数日均值,见表5。臭柏异形叶叶绿素荧光参数主体间效应检验,见表6。
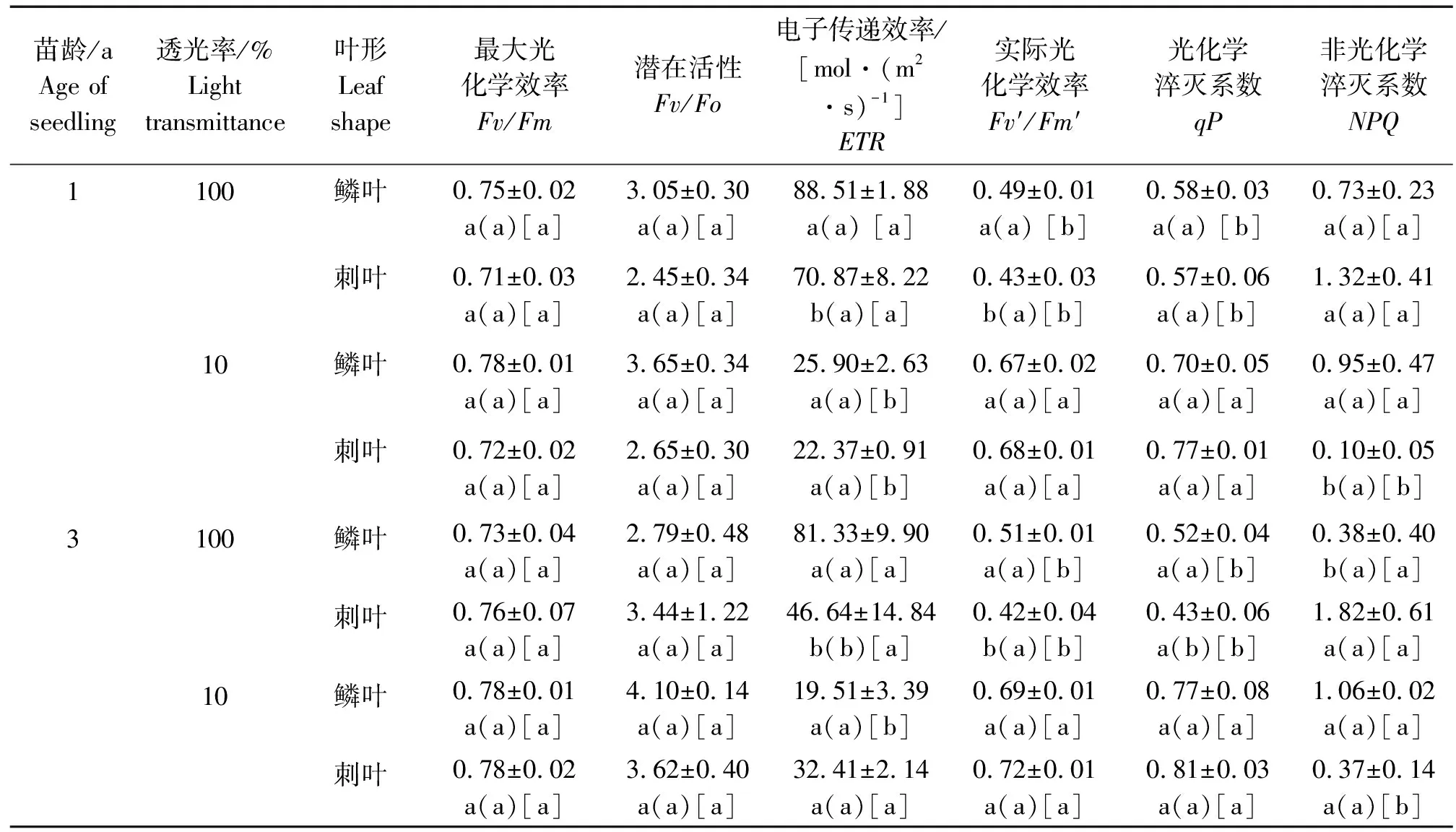
表5 不同光照环境下不同苗龄臭柏异形叶叶绿素荧光参数
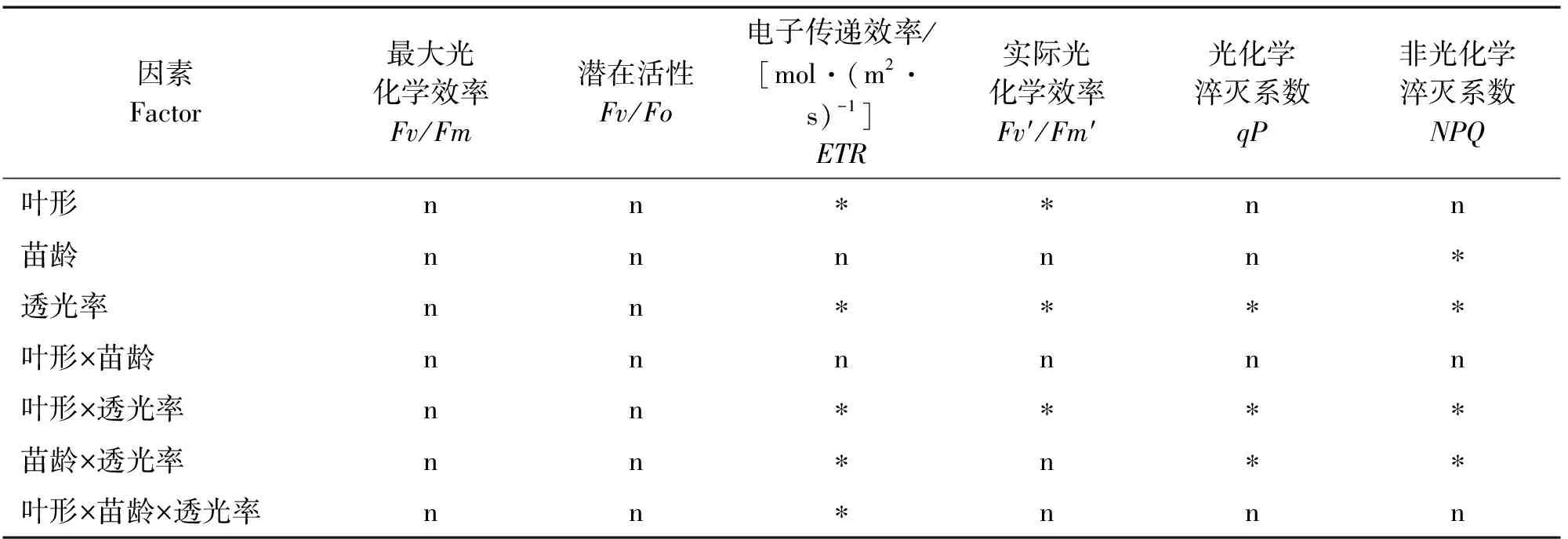
表6 臭柏异形叶叶绿素荧光参数主体间效应检验
由表5可知,全光环境下1年生臭柏鳞叶电子传递效率(ETR)和实际光化学效率(Fv′/Fm′)显著高于刺叶,遮光环境下1年生臭柏刺叶非光化学淬灭系数(NPQ)显著低于鳞叶。而遮光处理显著降低了1年生臭柏鳞叶ETR,显著增加了Fv′/Fm′和光化学淬灭系数(qP);遮光处理显著降低了1年生臭柏刺叶ETR和NPQ,显著增加了Fv′/Fm′和光化学淬灭系数(qP)。因此,1年生臭柏叶片叶绿素荧光参数受到遮光处理的影响大于叶形间差异带来的影响。全光环境下3年生臭柏鳞叶和刺叶ETR,Fv′/Fm′和NPQ均有显著差异;遮光环境下3年生臭柏鳞叶和刺叶叶绿素荧光参数无显著差异。而遮光处理显著降低了3年生臭柏鳞叶ETR,显著提高了Fv′/Fm′和qP;遮光处理显著提高了3年生臭柏刺叶Fv′/Fm′和qP,显著降低了NPQ。因此,遮光处理对3年生臭柏异形叶叶绿素荧光参数影响也高于叶形间差异。
由表6可知,叶形、苗龄和透光率对臭柏暗适应下Fv/Fm和Fv/Fo均无显著主效应和交互效应,而叶形、光照和苗龄对臭柏叶片的主效应及交互效应主要集中在光环境下叶绿素荧光参数。其中,叶形单因素主要影响臭柏异形叶ETR和Fv′/Fm′,苗龄单因素仅对NPQ存在显著影响,透光率对ETR,Fv′/Fm′,qP,NPQ均存在显著影响。叶形和苗龄双因素交互作用对臭柏异形叶叶绿素荧光参数均没有显著影响,叶形和透光率双因素交互作用对ETR,Fv′/Fm′,qP,NPQ均存在显著影响,苗龄和透光率双因素交互作用对ETR,qP,NPQ存在显著影响。叶形、苗龄和透光率三因素交互作用仅对ETR存在显著影响。因此,光照环境变化对臭柏异形叶叶绿素荧光参数产生主要影响。
3 讨论与结论
3.1 讨论
植物异形叶分异受到多种因素调控,如生长环境、生理活动和遗传机制的差异等[19-21]。本次试验通过研究光照、苗龄和叶片所处空间位置对臭柏异形叶鳞刺比影响,发现光照强度是影响1年生臭柏异形叶空间分异的主要因素,为生态异形叶性[8];而叶片所处空间位置是影响3年生臭柏空间分异的主要因素,为系统发育异形叶性[22]。这与胡杨异形叶性相异,其幼苗为条形叶,幼树兼有条形叶和披针形叶,成树自树冠下层至树冠上层分布披针形、卵圆形和阔叶等异形叶,由系统发育异形叶性转为生态异形叶性[5-6]。本次试验中,臭柏鳞叶和刺叶在1年和3年2个苗龄下对光环境适应性的差异也表明,臭柏异形叶在冠层上分布的差异与其叶片光合生理分异有关;相较于1年生臭柏刺叶,3年生臭柏刺叶在全光和遮光环境下净光合速率没有显著差异。因此,臭柏刺叶随着苗龄的增加由喜阳转变为有一定的耐阴性,而1年生和3年生臭柏鳞叶在全光环境下净光合速率均显著高于遮光环境。在本次试验中2种苗龄臭柏鳞叶均表现出喜阳性,2种叶片对光环境适应的分异也进一步导致1年生和3年生臭柏在冠层上分布的差异;1年生臭柏正处于叶片分异的关键时期,3年生臭柏完成叶片分异,冠层由上而下,鳞刺比逐渐降低。
3.2 结论
通过对10%和100%光照环境下1年生和3年生臭柏异形叶空间分异研究发现,苗龄、透光率和叶片部位单因素均对臭柏异形叶鳞刺比有显著影响,且双因素交互及三因素交互效应也对臭柏异形叶鳞刺比存在显著影响。1年生臭柏异形叶正处于分异的关键时期,空间分异受到光照环境的调控;3年生臭柏异形叶已完成空间分异,鳞刺比由冠层上部至冠层下部逐渐降低。
臭柏光合特性研究结果表明,透光率单因素对臭柏异形叶光合特性影响多于苗龄和叶形单因素的影响,双因素交互作用对臭柏异形叶光合特性影响较小,但三因素交互作用对臭柏光合特性存在影响。简单效应分析表明,遮光处理是影响1年生臭柏异形叶光合特性主要因素,而3年生臭柏异形叶光合特性影响因素则为叶形间的差异。
臭柏异形叶叶绿素荧光参数研究结果表明,透光率单因素对臭柏异形叶光合特性影响多于苗龄和叶形单因素的影响;双因素交互作用中,叶形和透光率、苗龄和透光率双因素交互作用对臭柏异形叶叶绿素荧光参数影响多于叶形和苗龄双因素的交互作用;三因素交互作用仅对电子传递效率存在显著作用;且苗龄、透光率、叶形三因素的主效应及交互效应对臭柏异形叶暗适应下叶绿素荧光参数均没有显著影响。简单效应分析结果表明,1年生和3年生臭柏异形叶叶绿素荧光参数主要受到透光率的影响,受到苗龄和叶形的影响较小。
综上,1年生臭柏正处于分异的关键时期,较易受到环境因素的影响;而3年生臭柏已完成叶形分异,其鳞叶在数量和生理上均存在优势,刺叶则表现出耐阴性。
猜你喜欢
花火B(2022年6期)2022-08-23 04:16:39
林业勘察设计(2021年3期)2022-01-04 03:29:06
农业科技通讯(2021年4期)2021-05-23 14:02:34
湖南林业科技(2020年2期)2020-09-18 05:36:44
石油化工建设(2018年1期)2018-07-10 09:49:50
汽车与安全(2017年7期)2017-08-03 17:20:12
汽车与安全(2017年6期)2017-07-25 09:57:08
西南农业学报(2017年5期)2017-06-23 08:14:29
汽车与安全(2017年3期)2017-04-26 01:25:27
北广人物(2017年2期)2017-03-25 04:52:40
