
闽西南红壤侵蚀区黑莎草功能性状特征及其与土壤因子的关系
2022-12-17 07:11:52郜鹏畅陈志彪陈志强牛玉娇区晓琳王海燕
热带亚热带植物学报 2022年6期
郜鹏畅,陈志彪,陈志强,牛玉娇,区晓琳,王海燕
(福建师范大学地理科学学院,湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
植物功能性状是与其生存发育有密切相关的核心属性,可以反映对环境变化的适应性,并且影响生态系统的功能[1–3]。植物在应对环境及生态系统变化时进行自我调节,从形态等方面表达其对环境的适应程度[4],功能性状的变化最大程度地减少了环境对自身不利的影响,反映出植物的最优“适应原则”[5],揭示植物群落对于环境变化的适应策略及在逆境条件下的生物多样性恢复机制[2]。植物功能性状之间是相互协同与权衡的关系,通过各性状间组合联动来实现生存与延续[6–7]。
叶片是植物进行光合作用和与大气能量交换的主要器官[8],目前对于影响叶片性状的因素较多且没有统一的认识,丁佳等[9]研究表明,土壤水分和氮含量影响森林生态系统幼龄植物叶片性状变异的主要因子,而在老龄林中影响叶片性状的主要因素有土壤有机质和磷含量。Jager 等[10]对新西兰热带雨林30 树种的研究表明,叶片氮磷含量、比叶面积、叶厚以及叶干物质含量均与土壤肥力存在相关关系。刘旻霞等[11]对甘南高寒草甸植物性状的研究表明,土壤pH 与土壤水分共同对植物性状产生影响。根系是植物吸收水分和养分的重要器官,根系功能性状是反映植物对水分和养分利用效率的重要指标[12],其研究是了解植物对资源利用效率高低的重要途径。根系对环境的适应能力是茎叶部分生长发育和增强植物自身环境适应性的重要因素[13],为应对生境变化,根系形成了自身特点,为适应极端干旱的环境植物会延长根深以抵御干旱[14],张帆等[15]研究表明,柠条人工林的细根根长密度受土壤温度等因子影响存在季节波动性。在植物生长发育的过程中,植物的功能性状与温度、水分和养分等有紧密的联系[16]。研究作为植物体重要的营养与表型器官的功能性状,有助于了解和掌握植物对生态系统中环境变化的适应程度。许多学者逐渐意识到可以通过对植物功能性状的研究辅助解决植物生态学问题,越来越多的研究开始侧重于在环境梯度的变化下所导致的植物功能性状的改变及对生态系统功能的改变等方面[17]。
福建省长汀县南方红壤侵蚀区是典型的生态系统严重退化区,植被恢复是恢复与重建退化生态系统的前提,寻找优良草本成为水土流失治理与退化生态系统恢复与重建的关键。在土壤贫瘠,干热化严重的退化生态系统中很多植被难以存活发育,但近年来我们在调查研究中发现,本土植物黑莎草(Gahnia tristis)能与代表性草本植物芒萁(Dicranopteris dichotoma)共生甚至生长更健壮,根系发达,地上与地下连接处保水作用十分明显。但经初步调查发现,黑莎草的分布存在明显的地域性,原因有待进一步探索。目前关于黑莎草的研究主要集中在植物群落多样性和生物量等方面[18–20],功能性状的相关研究鲜有报道。本研究以黑莎草为研究对象,分析黑莎草叶片和根系的功能性状特征,结合土壤因子的变化,明晰其功能性状与土壤环境因子的关系,探讨黑莎草在红壤侵蚀区的环境适应状况,以期为黑莎草投入侵蚀区水土流失治理工作提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于福建省西南部的长汀县(25°38′15″~25°42′55″ N,116°23′30″~116°30′30″ E),属于中亚热带湿润气候,多年平均气温18.3 ℃,无霜期年均260 d,1月平均气温8.1 ℃,7月平均气温27.2 ℃;年降水量为1 700~2 000 mm,且降水的季节变化较明显,其中3—8月为集中降水期,降水量占全年的70%以上。土壤以花岗岩风化淋溶形成的红壤和侵蚀红壤为主,抗侵蚀能力极低[21]。长期以来受自然和人为等因素的影响,植被遭到严重破坏,产生严重的土壤侵蚀现象,退化生态系统面积不断扩大。研究区内草本以黑莎草、芒萁和宽叶雀稗(Paspalum wettsteinii)为主。
1.2 样地选择、样品采集与处理
根据典型性和代表性原则并结合当地的植物调查,在长汀县来油坑和黄泥垄选取侵蚀样地3 处,分别命名为1、2、3 样地(表1,2),样地内的黑莎草植株均为自然生长。

表1 样地概况Table 1 Overview of plots
不同季节的光照强度、月均温和水分供应不同,植物以不同的形态和习性等特征来适应生长环境,另外不同温度和水分条件会影响土壤有机碳、总氮、总磷等含量和植物的生理功能及养分转移[22–23]。本研究选定采样时间为7月(夏季)和12月(冬季),分别代表一年中的2 个极端情况,通过测量冬夏季黑莎草各项功能性状指标和土壤理化性质,更有利于认识黑莎草对环境的适应性。分别于12月与次年7月采集样品,3 个样地自西向东布设20 m×20 m 的样方,再分别设置5 个5 m×5 m 的小样方,每个小样方内挑选1 株黑莎草植株。黑莎草地上部分采取收获法,随机选取20 片发育良好的完整叶片,用润湿滤纸相夹装入编号自封袋然后放入4 ℃冰盒保存,地下部分采取挖掘法,将伴有须根和细根的土壤一并放入编号自封袋,带回实验室。按照5 点法采集土样均匀混合成1 个样品后装入写好标签的自封袋中;同时采用环刀-铝盒采集各样点的原状土,用自封袋密封保存带回实验室。
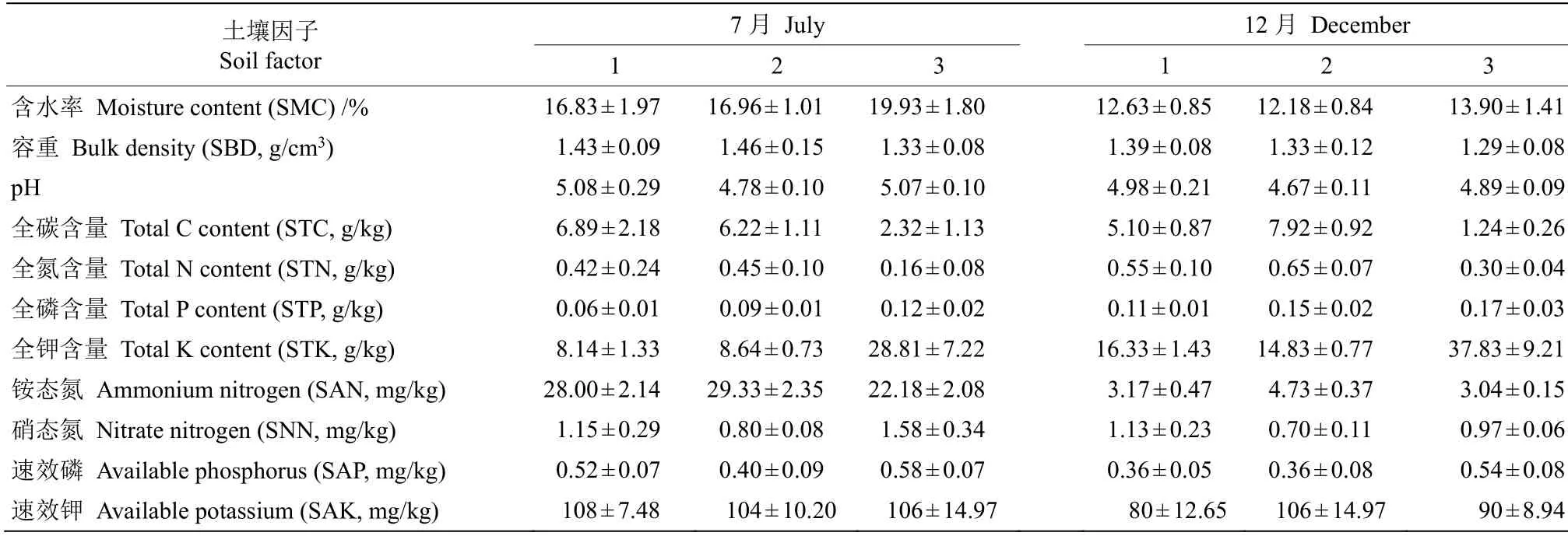
表2 土壤环境因子特征Table 2 Characteristics of soil environmental factors
在实验室中将采集回的植物样本用蒸馏水冲洗后阴干并将带有土壤的细根、须根用蒸馏水进行浸泡清洗,随后将黑莎草地上、地下部分样本分开装入牛皮纸信封袋中于75 ℃鼓风干燥箱中烘干后称量质量,计算地上部分(叶片)、地下部分(根系)的生物量。最后将黑莎草样本用粉碎机进行粉碎,再过100 目筛装入自封袋保存于干燥处,用于测定元素含量。除去土样中的石子、树根、枯草等杂质,放置在干燥通风的地方自然风干。风干后的土壤样本研磨过100 目筛,用于测定土壤养分等。
1.3 方法
叶片表型性状的测定挖取黑莎草植株前,随机选取3 片完整叶片,分别在叶基、叶中和叶尖部分用PJ-4N 便携式叶绿度测量仪(郑州明检农业科技有限公司)测定叶绿素相对含量(SPAD 值)。然后用游标卡尺(精度0.02 mm)按叶基、叶中和叶尖的顺序测量叶片宽度(mm)和厚度(mm)。清除叶片表面杂质,使用佳能2580S 扫描仪对叶片进行扫描,运用Image J 图像分析软件计算叶面积(cm2)、叶长(cm)。以上数据均取平均值。对叶片编号入档。采用精度为0.0001 g 的电子天平称量叶片鲜质量(g),然后置于4 ℃水中浸泡12 h,取出吸干表面水分,称量饱和叶片鲜质量(g)。叶片样品在烘箱内经105 ℃杀青30 min 后,在70 ℃下烘干,称量叶片干质量(g)。使用Excel 2010 计算比叶面积(specific leaf area,SLA )、叶干物质含量(leaf dry matter content,LDMC)、叶组织密度(leaf tissue density,LTD)和叶相对含水率(leaf relative water content,LRWC),比叶面积(cm²/g)=叶面积(cm²)/叶干质量(g); 叶干物质含量(g/g)=叶干质量(g)/叶饱和鲜质量(g); 叶组织密度(mg/mm3)=叶干质量(g)/[叶面积(cm²)×叶厚(mm)]; 叶相对含水率(%)=[叶鲜质量(g)-叶干质量(g)]/[叶饱和鲜质量(g)-叶干质量(g)]×100%。
根系表型性状的测定将植物根系上附着的土用水洗干净,并分离出细根(直径<2 mm)装入自封袋内并做好标签,放于4 ℃冰箱保存鲜样。用佳能2580S扫描仪对根系进行扫描,采用WinRHIZO 8.0软件对根系形态进行分析。测量根长(root length,RL)、根表面积(root superficial area,RSA)、根体积(root volume,RV)和平均根直径(root average diameter,RAD)等指标,根系经70 ℃烘干后,利用精度为0.000 1 g 的电子天平秤称量根干质量(g),计算比根长、根组织密度、比根面积。比根长(m/g)=根长(m)/根干质量(g);根组织密度(mg/cm3)=根干质量(g)/根体积(cm3);比根面积(cm²/mg)=根表面积(cm2)/根干质量(g)。
叶片和根系养分含量的测定将采集的叶片和根系于烘箱中105 ℃杀青30 min 后,70 ℃烘干,分别进行粉碎研磨、过100 目尼龙筛,装袋待测。叶片和根系全碳和全氮含量采用Elemantar Vario ELⅢ碳氮元素分析仪(德国)测定,全磷含量采用高氯酸消煮法,用Skalar san++连续流动分析仪(荷兰)测定;全钾采用氢氟酸-高氯酸消煮法,用FP 640 火焰光度仪(中国)测定。
土壤因子测定用五点法采集10 cm 厚土样,每样方设置3 组重复,相同样地不同样方的土壤混合均匀,同时,采用环刀-铝盒采集原状土。采用环刀法测定土壤容重;铝盒烘干法测定土壤含水率;以水∶土=1∶2.5 水浸-电位测定土壤pH 值。有机碳和全氮采用Elemantar Vario MAX 碳氮元素分析仪测定;全磷采用高氯酸消煮法、用Skalar san++连续流动分析仪测定;全钾采用氢氟酸-高氯酸消煮法,用FP 640 火焰光度仪测定; 速效氮(铵态氮、硝态氮)采用2 mol/L KCl 浸提法,用Skalar san++连续流动分析仪测定;速效磷采用Mehlich3 法浸提,用Skalar san++连续流动分析仪测定;速效钾采用1 mol/L NH4OAc 浸提,用FP 640 火焰光度仪测定。
1.4 数据处理和分析
采用Excel 2010 和SPSS 22 软件对数据进行统计分析,采用单因素方差分析(One-Way ANOVA)及LSD 多重比较进行显著性检验,用冗余分析(RDA)法对黑莎草叶片、根系表型性状与土壤理化性质之间的关系进行分析。相关图表在Excel 2010、Origin 2018 和Canoco 5 内完成。
2 结果和分析
2.1 黑莎草功能性状特征
2.1.1 表型性状特征
除叶面积外,其余叶片表型性状在冬夏季间均存在差异(图1)。叶长、叶宽、叶组织密度和叶绿素含量表现为夏季显著大于冬季(P<0.05),比叶面积则相反。3 个样地的叶面积在冬夏季间差异不显著。

图1 黑莎草叶片表型性状特征。柱上大写字母表示7 和12月间差异显著(P<0.05),小写字母表示不同样地间差异显著(P<0.05)。下同Fig.1 Leaf phenotypic characteristics of Gahnia tristis.Capital letters upon column indicate significant differences between July and December at 0.05 level,while lowercase letters indicate differences among plots at 0.05 level.The same below
根系表型性状同样表现出差异性(图2)。3 个样地的根直径在冬夏季的差异均不显著(P>0.05);样地1 和样地2 的比根长、根组织密度和比根面积在冬夏季均有显著差异(P<0.05);冬季的比根长和比根面积高于夏季,根组织密度则相反。样地2 夏季的比根长和比根面积均最小。

图2 黑莎草根系表型性状特征Fig.2 Root phenotypic characteristics of Gahnia tristis
2.1.2 养分含量特征
从图3可见,黑莎草叶片的碳、氮、磷和钾含量分别为403.65~433.45、4.17~5.82、0.14~0.24 和11.97~25.20 g/kg。样地3 的叶片碳含量最低,氮、磷、钾含量最高,冬夏季叶片碳、氮和钾含量分别仅在样地1、样地3和样地2存在显著差异(P<0.05),而叶片磷含量在样地1 和样地3 均差异显著(P<0.05)。

图3 黑莎草叶片养分含量特征Fig.3 Characteristics of nutrient content in leaf of Gahnia tristis
从图4可见,根系的碳、氮、磷和钾含量分别为258.85~428.22、1.49~2.76、0.09~0.21 和5.6~21.27 g/kg。样地3 的根系碳含量最低,钾含量最高,且根系碳含量在冬夏季的差异显著(P<0.05);冬季根系氮、磷含量高于夏季,且样地2 和样地3 的差异显著(P<0.05)。

图4 黑莎草根系养分含量特征Fig.4 Characteristics of nutrient content in root of Gahnia tristis
总体上,除碳含量和12月根系氮含量外,其余养分均以样地3 较高,冬季的根系养分含量均高于夏季,养分的分配上叶片高于根系。
2.2 相关分析
由表3和4 可见,叶组织密度与比叶面积呈显著负相关,而与叶绿素含量则呈显著正相关关系;叶绿素含量与叶片全碳和全氮含量相关;叶干物质含量与叶片氮、磷、钾含量均呈负相关关系。根组织密度与比根长、比根面积和根系全磷含量均呈显著负相关,叶片和根系氮、磷、钾含量之间均显著正相关。

表3 黑莎草叶片功能性状间的相关系数Table 3 Correlation coefficient of leaf functional traits of Gahnia tristis
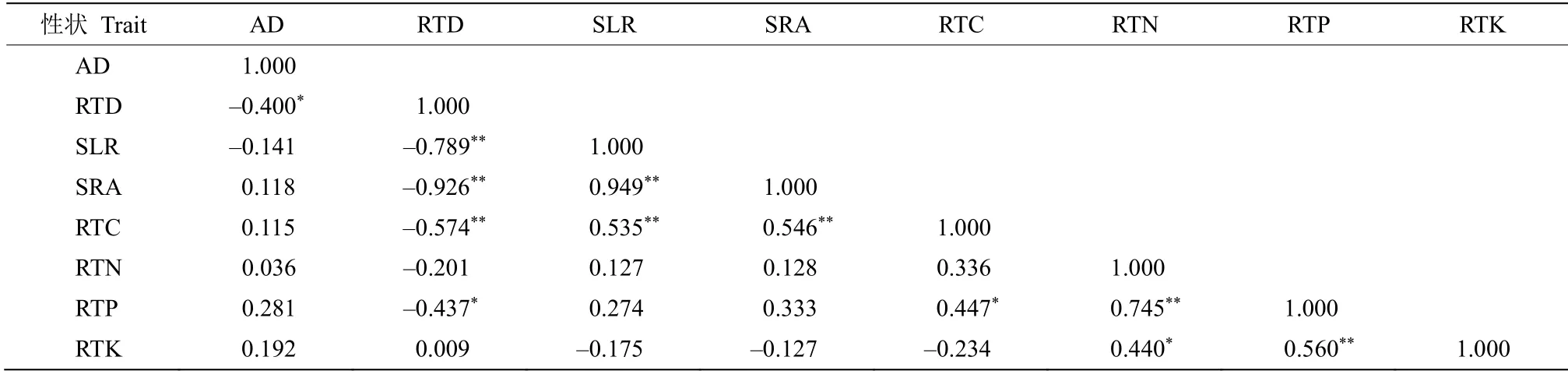
表4 黑莎草根系功能性状间的相关系数Table 4 Correlation coefficient of root functional traits of Gahnia tristis
2.3 功能性状对土壤因子的响应
为进一步探讨黑莎草叶片和根系功能性状与土壤因子间的关系,将7 和12月的功能性状指标分别与土壤含水率、pH、容重、土壤全量养分和速效养分等11 个指标进行冗余分析(RDA)。由表5可见,12月第1、2 排序轴的特征值分别为0.982 9和0.916 6,共解释了60.28%的叶功能性状变化和73.9%的叶功能性状与土壤因子关系;7月第1、2排序轴的特征值分别为0.944 1 和0.977 4,共解释了66.94%的叶功能性状变化和76.21%的叶功能性状与土壤因子关系。12月第1、2 排序轴的特征值分别为0.985 2 和0.996 0,共解释了72.31%的根系功能性状变化和74.92%的根系功能性状与土壤因子关系;7月第1、2 排序轴的特征值分别为0.991 1 和0.986 0,共解释了72.03%的根系功能性状变化和76.90%的根系功能性状与土壤因子关系。可见黑莎草叶片与根系功能性状和土壤因子有较好的相关关系,前两排序轴能较好地反映黑莎草功能性状和土壤因子的关系。由图5、6可见,12月黑莎草叶片功能性状主要受土壤全碳和铵态氮含量的影响,根系功能性状主要受全量碳、磷、钾含量的影响。7月黑莎草叶片功能性状主要受土壤全磷、全氮和硝态氮的影响,根系功能性状主要受土壤全氮、全磷和速效磷的影响。可见黑莎草叶片与根系功能性状和土壤因子有较好的相关关系,前2 排序轴能较好地反映黑莎草功能性状和土壤因子的关系。由图5、6 可见,12月黑莎草叶片功能性状主要受土壤全碳和铵态氮含量的影响,根系功能性状主要受全量碳、磷、钾含量的影响。7月黑莎草叶片功能性状主要受土壤全磷、全氮和硝态氮的影响,根系功能性状主要受土壤全氮、全磷和速效磷的影响。

表5 叶片功能性状(LFT)与土壤因子(SF)的RDA 分析Table 5 RDA analysis of leaf functional traits (LFT) and soil factors (SF)
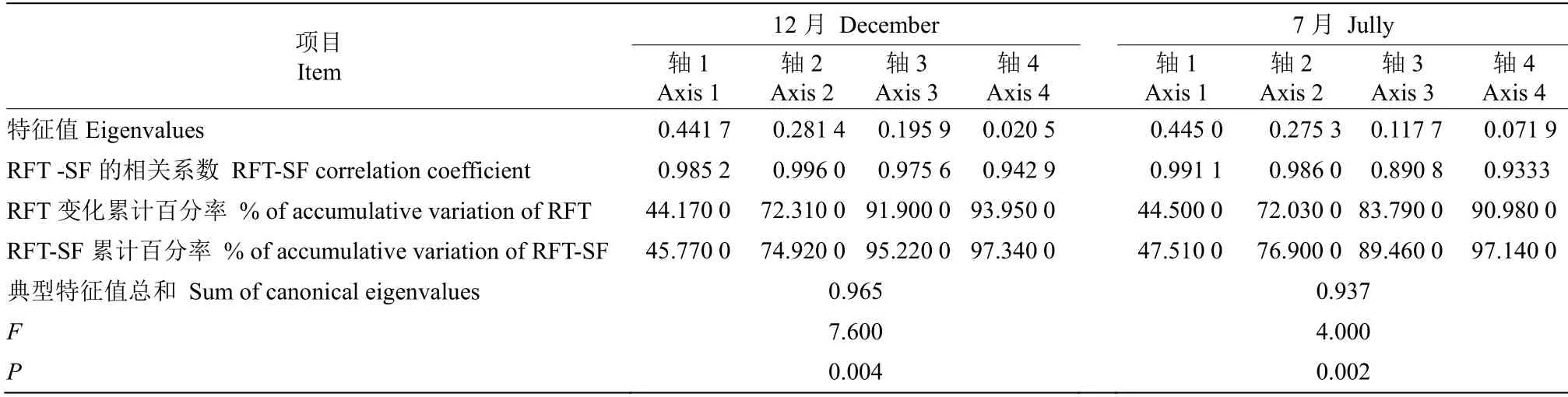
表6 根系功能性状(RFT)与土壤因子(SF)的RDA 分析Table 6 RDA analysis of root functional traits (RFT) and soil factors (SF)
3 结论和讨论
3.1 叶片和根系功能性状及其相关性
植物功能性状不是孤立发挥作用的,在长期适应生存环境过程中,黑莎草叶片和根系通过协同进化并最终形成最佳功能性状组合,响应外部环境的同时对生态系统产生一定影响[10]。植物通过比叶面积与叶组织密度共同作用来维持水分的平衡,具体表现为比叶面积减小和叶组织密度增大[24]。本研究中比叶面积和叶组织密度也呈显著负相关关系。目前有关叶面积与比叶面积的关系并未形成一致的认识[25–26],本研究中黑莎草的叶面积与比叶面积不存在显著相关关系,表现出不同植物对特定环境的适应性不完全一致。叶绿素是植物光合作用的基础[27],黑莎草叶绿素含量与叶片氮含量呈显著正相关关系,这与前人[28]的研究结果一致。比根长很大程度上反映了根系对水分与养分的吸收能力,根组织密度则反映根系的抗拉力和防御能力,黑莎草根系在12月表现出较高的比根长与较低的根组织密度来维持对养分的吸收,在7月保持较高的根组织密度以防止水分过度流失以及高温损伤,这与戚德辉等[29]对铁杆蒿(Artemisia sacrorum)对环境变化的适应研究结果一致,铁杆蒿在干旱贫瘠的环境中通过较高的比根长与较低的根组织密度维持对水分和养分的吸收,黑莎草的比根长与根组织密度也存在显著负相关关系。通常具有较大比根长或比根面积的植物根组织密度较小,可加快根系延伸与周转速率,提高对养分和水分的吸收能力,是植物吸收养分的策略[10]。黑莎草根组织密度与比根长、比根面积间呈显著负相关,反映其生长与防御之间的权衡关系[30]和对环境有较强适应能力[31]。本文对黑莎草根系表型性状的研究,仅选取了根直径、根组织密度、比根长和比根面积,指标相对叶片而言偏少,今后可增加如根尖数、根分支数等拓扑结构方面的形状指标,为根系性状研究提供更全面的参考。

图6 黑莎草根系功能性状与土壤因子的RDA 排序图Fig.6 RDA ordination diagram of root functional traits and soil factors of Gahnia tristis
叶片氮、磷平均含量均低于我国东部南北样带654 种陆生植物叶片的平均含量[32],根系碳、氮、磷平均含量均远低于中国植物细根的碳、氮、磷平均含量(分别为437.90、9.16 和0.95 g/kg)[33],说明黑莎草在侵蚀区养分含量低的环境中适应与耐受程度相对较高。植物叶片对光合、呼吸以及化合物的合成具有重要作用,黑莎草叶片养分含量总体上高于根系。7月属于黑莎草生长旺盛期,通过自身调节将更多的养分分配到地上部分从而导致7月根系养分含量低,而12月份为越冬及明年生长储存养分导致根系养分升高,黑莎草养分含量及其分配受所处环境和养分有效性的制约,同时也与自身的生长节律密切相关[34]。叶片和根系的氮、磷元素具有极显著的正相关关系,这与前人[35–37]的研究结果一致,作为陆地生态系统植物生长的主要限制元素,氮、磷两者的正相关关系符合全球尺度的普遍性[38]。
芒萁是南方红壤侵蚀区具有耐寒耐贫瘠特征的重要地带性植物[39],黑莎草的叶面积、比叶面积、根直径和根组织密度及叶片与根系的钾含量远大于芒萁[40],较大的比叶面积有利于黑莎草体内养分保持[41–42],较高的根组织密度则体现黑莎草在贫瘠环境中的适应性[43–44],黑莎草叶片和根系中较高的钾含量可促进糖类及蛋白质合成,提高纤维素和木质素含量,使得根茎膨大坚韧[45],显示出比芒萁更大生物量、良好水土保持潜力且籽粒富含油脂,具有较高经济价值。
3.2 功能性状对土壤因子的响应
植物性状是由遗传因素和环境条件共同决定的[46],土壤环境条件通过改变植物性状影响植物生存策略的调整[14]。从冗余分析结果来看,土壤养分是影响黑莎草功能性状的主要因素,黑莎草12月功能性状主要受土壤碳、氮和磷含量的影响,7月功能性状主要受土壤氮、磷含量的影响。土壤养分是植物氮、磷营养的基础,进而影响植物代谢与生长[47]。黑莎草叶片与根系养分含量与土壤养分之间具有很强的相关性。磷含量是限制植物生长的重要因子,低磷的环境使得植物不断调整自身形态来适应环境[48],黑莎草叶面积和根直径与土壤全磷显著正相关。比叶面积可反映植物碳的获取策略[49],在贫瘠土壤中的植物有较小比叶面积,养分充足的土壤环境中植物有较大的比叶面积[50–51],本研究中黑莎草比叶面积与土壤全氮和速效钾显著正相关。植物叶片与根系的组织密度则体现植物的防御能力,叶组织密度高的植物分配较多氮素提高细胞壁韧性从而防治叶片损伤或失水过多[52],本研究中叶组织密度与土壤氮含量存在密切相关性。
本研究主要侧重于黑莎草生长指标与土壤因子的相关性方面,在今后的研究中可考虑添加对其它环境因子的研究,如降水量、温度、湿度、日照强度以及日照时长等。
黑莎草叶片表型性状在冬夏季间存在显著差异。叶长、叶宽、叶组织密度和叶绿素表现为夏季显著大于冬季,根系表型性状与叶片相比季节差异较小,黑莎草根系表型性状比叶片更具稳定性,冬季的根系养分含量均高于夏季,养分的分配上叶片养分高于根系养分。叶组织密度与叶绿素显著正相关,与比叶面积显著负相关;根组织密度与比根长和比根面积均显著负相关,叶片和根系养分之间均显著正相关。土壤碳、氮、磷含量是影响黑莎草功能性状主要因子。黑莎草通过调节功能性状指标对环境变化表现出适应性策略,可作为地带性植物引入南方红壤侵蚀区的植被恢复和水土流失治理。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23 04:44:18
疯狂英语·读写版(2022年1期)2022-01-29 01:06:57
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
文史博览·文史(2021年1期)2021-04-01 14:04:00
落叶果树(2021年6期)2021-02-12 01:28:54
文苑(2020年12期)2020-04-13 00:54:10
新疆农业科学(2020年1期)2020-02-14 03:22:46
海峡姐妹(2019年1期)2019-03-23 02:42:46
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国麻业科学(2015年5期)2015-12-28 06:22:11
