
谷子不同核心种质的小米氨基酸组分积累特性分析及其评价
2022-12-14 08:31王啸旗郭宁馨李旭凯高建华王兴春侯思宇赵雄伟
山西农业科学 2022年12期
舒 军,王啸旗,郭宁馨,李旭凯,高建华,王兴春,侯思宇,赵雄伟
(1.山西农业大学 生命科学学院/杂粮种质创新与分子育种山西省重点实验室,山西 太原 030031;2.山西农业大学 农学院,山西 太谷 030801)
谷子(Setaria italica)是耐贫瘠抗旱作物,在亚洲半干旱地区(印度、中国和日本)以及南欧广泛种植,有着悠久的种植历史[1-3]。近年来,随着新一代谷子高质量全基因组序列的公布和谷子突变体“xiaomi”的出现,其作为C4模式植物也得到了越来越多研究人员的关注和认可[4-8]。谷子去壳后称为小米,主要包括维生素、碳水化合物、叶酸、类胡萝卜素和蛋白质等营养成分,并且具有较高的氨基酸含量[9-11]。有研究表明,亮氨酸、异亮氨酸和缬氨酸等人体必需氨基酸在蛋白质合成方面有明显的增强作用[12]。此外,小米中富含的甲硫氨酸在抗脂肪肝和促进人体胰岛素分泌方面有重要作用,并且可以促进血清素的分泌,还有助睡眠的功效[13-14]。因此,提升小米中的氨基酸含量是改善人们膳食结构多样性的有力保障。
前期研究者已对小米的氨基酸组分及其含量进行分析。冯耐红等[15]研究结果表明,供试10种山西主推品种小米富含17种氨基酸,其中必需氨基酸种类齐全,含量丰富,尤其谷氨酸和亮氨酸含量较高。与其他作物相比,小米中8种人体必需氨基酸含量丰富,占整个氨基酸总量的41.9%,比大米、小麦粉和玉米分别高出56.4%、80.6%、42.6%[14,16]。谷子中不同种质之间氨基酸含量差异较大,特别是赖氨酸的含量较低,不利于优质小米品种的推广。目前,主要通过培育高赖氨酸品种、氨基酸强化和饮食搭配等手段来弥补小米赖氨酸缺乏的这一特点。
虽然目前对于小米的营养成分已有研究报道,但其主要从蛋白质、碳水化合物、脂肪和维生素等方面进行[14,17],且谷子种质资源的氨基酸积累特性研究相对较少。因此,了解清楚来源不同的谷子种质之间氨基酸积累特性,以及对氨基酸积累的影响,对筛选和创制优质品种具有重要意义。本研究以来源于我国不同省份的154份谷子核心种质为研究材料,连续2 a分析小米中17种氨基酸含量,分析不同氨基酸在谷子小米中的积累特性,以及不同种质来源或亚群对氨基酸含量的影响,并筛选核心的优质种质,为选育谷子优良品种提供参考。
1 材料和方法
1.1 试验材料
基于中国农业科学院刁现民研究员团队[18]提供的重测序核心谷子种质,进一步以尽可能囊括不同地理来源、不同基因型品种多样性为原则,选择有代表性的我国154份谷子核心品种(图1),包括北京2份、甘 肃14份、河 北29份、河 南12份、黑 龙 江12份、吉林17份、江西3份、辽宁7份、内蒙古8份、宁夏6份、山东9份、山西25份、陕西3份、四川5份和新疆2份。此外,根据前期谷子不同种质遗传亚群研究[18],试验材料中58份种质来自于春谷亚群,96份种质来自于夏谷亚群。

图1 谷子154份核心种质来源分布Fig.1 Origin and distribution of 154 core germplasms of foxtail millet
1.2 试验方法
试验材料均于2019—2020年种植于山西农业大学杂粮育种基地中,行宽2 m,行距20 cm,单穴播种,株距15 cm,每份种质种植3行。待谷穗成熟后,剪取株型和穗大小一致的8穗,晾晒干后混合脱粒并使用小型垄谷机(TP-JLG-2018)去壳碾成小米,去杂过筛(直径1 mm圆孔筛)制得精小米,每个品种取10 g备用。
1.3 小米中17种氨基酸含量的测定
将1.2中精选的等量小米放于近红外光谱仪样品池中,保持自然状态(样品量高度、松紧度一致),利用已建立的小米氨基酸近红外光谱分析模型对小米中17种氨基酸含量进行测定,分别为谷氨酸(Glu)、精 氨酸(Arg)、脯 氨 酸(Pro)、天 冬 氨 酸(Asp)、苏氨酸(Thr)、甲硫氨酸(Met)、赖氨酸(Lys)、组 氨 酸(His)、丙 氨 酸(Ala)、苯 丙 氨 酸(Phe)、酪氨酸(Tyr)、丝氨酸(Ser)、甘氨酸(Gly)、半胱氨酸(Cys)、缬氨酸(Val)、异亮氨酸(Ile)、亮氨酸(Leu)。样品扫描前,仪器需预热0.5 h以上,以仪器内置参比作背景进行校正,所用的近红外光谱仪型号为Foss DS2500F。每个样品混合重复装样2次,测量结果取平均值。
1.4 数据处理
采用Microsoft Excel 2016和SPSS 21.0进 行变异系数和统计分析,并通过SPSS 21.0进行方差分析和主成分分析。使用R语言Hmisc包(Pearson系数)和Stats包的hclust函数(系统聚类法)进行相关性分析和聚类分析,并利用corplot包和ggplot包绘制相关性图、箱线图及聚类图。
利用综合主成分值模型,先计算出各主成分的载荷值(Ui),再利用载荷值和各氨基酸含量标准值得出主成分表达式(Yi),最后结合各主成分的权重,得出154份小米各氨基酸的综合主成分值(Y),进而综合评价小米氨基酸组分特点,筛选出谷子重点氨基酸种质资源。
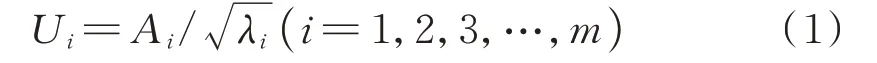
式中,Ui为主成分载荷矩阵,Ai为因子载荷矩阵,λi为主成分特征值,m为提取主成分的个数。
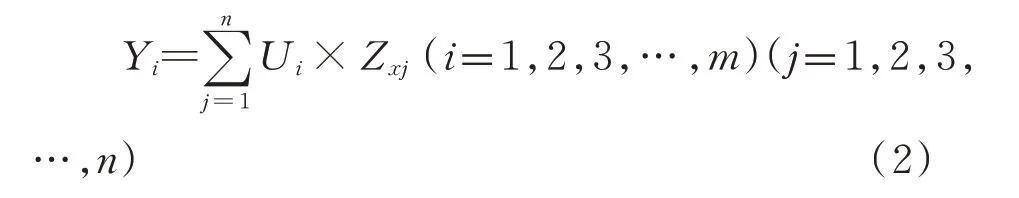
式中,Yi为第i主成分,Zxj为第j个氨基酸含量的标准值,n为氨基酸个数17。

2 结果与分析
2.1 小米17种氨基酸组分的遗传多样性分析
供试谷子种质在2019—2020年的氨基酸含量分析及其变异程度如表1所示。

表1 供试谷子种质在2019—2020年的氨基酸含量分析及其变异程度Tab.1 Amino acid content analysis and its variation in test germplasms of foxtail millet in 2019 and 2020
通过测定154份谷子种质小米的17种氨基酸含量(表1)发现,小米中总氨基酸含量平均值为11.93%。谷子小米中富含大量的Glu、Ala和Ile(大于1%),其中,Glu的平均含量最高,为2.78%;Pro、Asp含 量 次之,平均 含 量 为0.84%,而Thr、His、Lys、Cys含量较低(小于0.4%),其中Lys和Cys平均含量较低(0.19%)。对2019—2020年小米不同氨基酸含量进行方差分析发现,除His、Val无显著差异外,其余的15种氨基酸年份间均存在显著差异(P<0.05),其中Glu的F值高达838.56。氨基酸含量积累在不同谷子种质间也存在显著差异,变异范围因不同氨基酸差异较大,其中,Glu、Cys、Leu、Phe和His氨基酸变幅最大,呈现丰富的遗传多样性。
通过对氨基酸含量的变异系数分析发现,除Glu和Pro的变异系数在2 a相差大于18%外,其余氨基酸都小于4%,说明前者更容易受环境的影响。测定的17种氨基酸中有10种(如Glu、Pro、Cys等)的变异系数都高于10%,其中Clu变异系数最高,平均值为43.63%,表明这些氨基酸离散程度较大,有丰富的变异性,其余7种氨基酸的变异系数在3.81%~9.71%,暗示了其受环境和来源的影响更小,具有更高的稳定性。
2.2 不同来源及遗传亚群的小米氨基酸含量特征分析
为了研究不同地理来源的小米氨基酸变化特征,对2019—2020年连续2 a的各地区(品种数大于10)小米的氨基酸总含量进行分析。由图2可知,虽然2019年不同地区的氨基酸总含量普遍高于2020年,但不同地区的小米氨基酸总含量在2 a间趋势较为一致。在2019年,黑龙江和甘肃地区总氨基酸平均含量较高,分别为13.87%和13.82%,其次是吉林和山西地区(12.96%和12.90%),河南与河北地区最低。

图2 不同环境来源的小米总氨基酸含量分布Fig.2 The distribution of total amino acid content in fox⁃tail millet grains from different environmental origins
通过对谷子不同遗传亚群的氨基酸含量分析发现(图1),来源于甘肃和黑龙江地区的种质均属于春谷亚群,山西地区种质占据56%。河南地区的种质均属于夏谷亚群,河北和吉林地区种质的96.5%和88.2%属于夏谷。此外,通过对连续2 a不同遗传亚群的小米氨基酸含量特征分析发现(图3),春谷亚群小米的Ile、Leu等5种人体必需氨基酸及Ala、Arg、Asp等8种非必需氨基酸的含量显著高于夏谷亚群(P<0.05),Thr仅在2019年春谷亚群中极显著高于夏谷亚群(P<0.01)。以上结果表明,春谷亚群小米的品质明显高于夏谷品质,也暗示了甘肃、黑龙江和山西等北方地区更适宜优质谷子品种的种植。
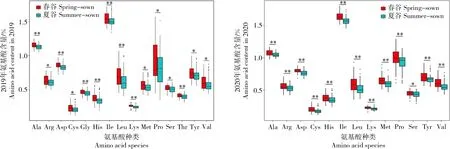
图3 2019、2020年春谷与夏谷亚群的氨基酸含量比较分析Fig.3 Comparative analysis of amino acid content between spring-sown millet group and summer-sown millet group in 2019 and 2020
2.3 小米17种氨基酸之间的相关性分析
为了研究小米中不同氨基酸含量之间的关系,本研究对2019—2020年连续2 a的17种氨基酸含量进行相关性分析(图4)。结果显示,除了Phe、His和Met外,Glu与其他13种氨基酸都呈现显著正相关关系,其中与Asp的相关性最高,相关系数为0.864;与Tyr的相关性最小(0.351)。Phe与Ser(0.781)、Thr、Gly呈极显著正相关,与His(-0.869)、Tyr等10种氨基酸呈极显著负相关,而与Glu、Asp、Lys不相关。Lys除了与Phe、Tyr、Val不相关外,与其余的13种氨基酸均呈现极显著正相关,其中与Asp的相关 系 数 最大(0.967)。Leu和Ile与Met(0.922和0.844)、Cys(0.820和0.943)等12种氨基酸呈极显著正相关,仅与Phe(-0.688和-0.451)呈 显 著 负 相 关,与Thr、Ser、Gly不 显 著 相 关。总之,小米中的17种氨基酸之间具有一定的相关性,大部分氨基酸之间呈现显著正相关,Phe、Ser、Thr与其他大量氨基酸存在显著负相关,暗示了不同氨基酸在积累过程中存在一定的协同或抑制关系,筛选氨基酸含量高的优质种质较为复杂。

图4 不同氨基酸之间的相关性分析Fig.4 Relationship analysis among different amino acids
2.4 小米17种氨基酸组分的主成分分析与综合主成分评价
为了筛选核心种质中的优质种质,本研究基于小米中不同氨基酸含量之间的相关性结果,进一步对2019—2020年的小米各氨基酸含量进行主成分分析,通过综合主成分值评价核心种质在氨基酸方面表现出的优质特性。结果显示,2019年第1、2主成分和2020年第1、2、3主成分的累积贡献率分别为87.215%和89.177%(表2),主成分组成的单元包含了17种氨基酸。此外,对其进行成分矩阵载荷分析发现(图5),Leu、Cys、Arg、Tyr、Met、His、Ile、Val在2019、2020年 的 主 成 分1中 载 荷 分 别 大于0.2和0.1,而Thr、Ser、Gly、Lys、Asp、Phe、Glu在主成分2中载荷大于0.28,其中Thr最大(0.43)。Met、Ile和Phe在2020年 的 主 成分3中有很高的载荷。因此,通过主成分的划分可以将小米17种氨基酸含量转化为3个独立的主成分指标,从而降低小米氨基酸分析的难度,会更加直观地展示氨基酸在种质之间的差异关系。
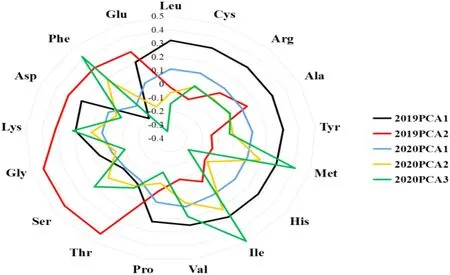
图5 2019、2020年不同氨基酸主成分矩阵的载荷雷达Fig.5 Load radar diagram for the principal component matrix of different amino acids in 2019 and 2020
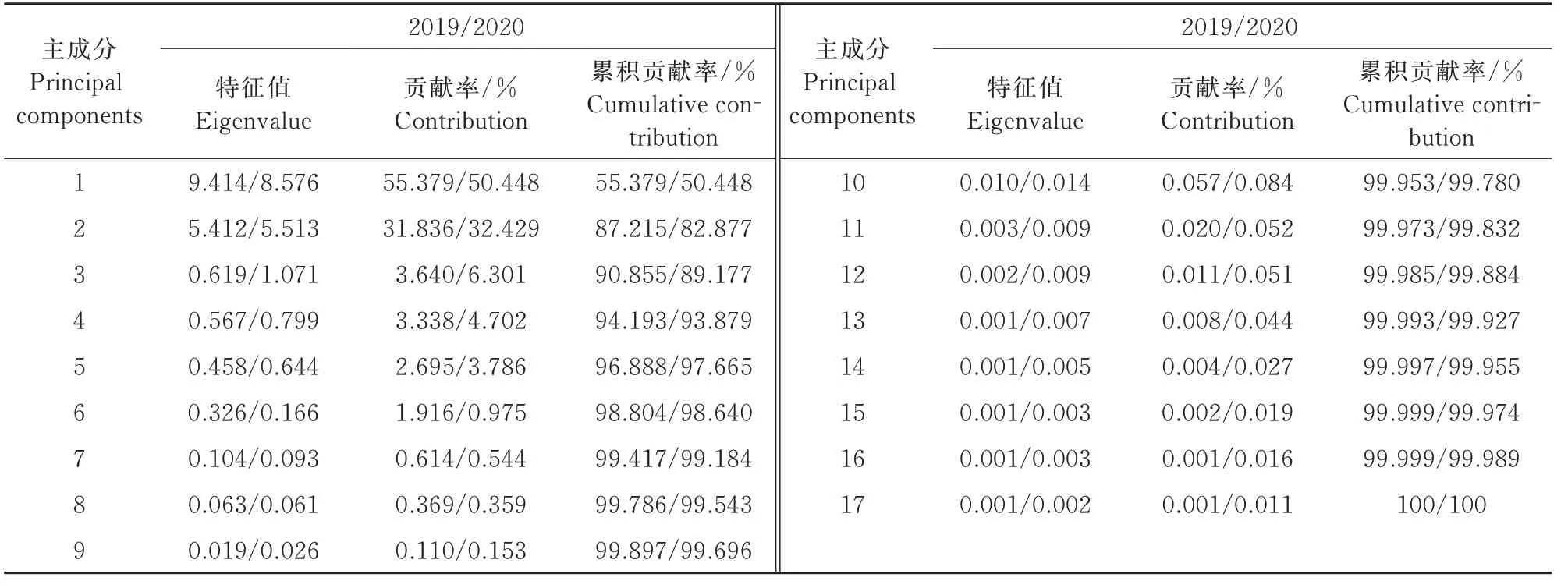
表2 谷子种质17种氨基酸含量的主成分分析Tab.2 Principal component analysis of contents of 17 amino acids in foxtail millet germplasms
对小米17种氨基酸含量进行标准化处理,以2 a各主成分所对应的特征值占所提取主成分总的特征值之和的比例作为权重,得到主成分综合模型,2019年综合主成分值Y=0.635×(0.20XGlu+0.31XArg+0.22XPro+0.22XAsp-0.01XThr+0.28XMet+0.19XLys+0.28XHis+0.31XAla-0.2XPhe+0.3XTyr-0.03XSer+0.06XGly+0.31XCys+0.25XVal+0.28XIle+0.32XLeu)+0.365×(0.28XGlu+0.05XArg-0.01XPro+0.31XAsp+0.43XThr-0.13XMet+0.32XLys-0.13XHis+0.13XAla+0.3XPhe-0.14XTyr+0.42XSer+0.42XGly-0.09XCys-0.09XVal-0.02XIle-0.03XLeu)。通过 上 述公式计算出每个谷子种质在2020年的综合主成分值,结果表明,Asp、Ala、Lys、Glu、Arg氨基酸对主成分综合值的作用最大(>0.2)。结合并比较2 a的综合主成分值,发现甘肃的陇谷10号、陇粟2号、陇谷5号,河北的毛毛谷、冀谷17,黑龙江的嫩选十四号、嫩选十一号,辽宁的大毛毛谷,内蒙古的白油沙、红苗齐头红,山西的白毛粱谷、猫蹄谷、大同黄以及新疆的二白谷等品种的综合主成分值均排前10%。
2.5 基于小米氨基酸组成的聚类分析
为了更全面并多角度评价核心种质和筛选出优异氨基酸种质的谷子资源,对154份2 a的小米氨基酸含量进行聚类分析(图6)。结果显示,可将154份谷子核心种质分为4个类群。其中,GroupⅠ以品种B029(山西白毛粱谷)、B077(新疆二白谷)、B110(甘肃陇谷5号)、B139(黑龙江嫩选十一号)和B112(甘肃陇粟2号)为代表,其平均总氨基酸含量最高,达15.37%,Lys平均含量最高,为0.27%,且除Phe、Met、Thr外,其余14种氨基酸含量都要显著高于其他三大类群,其中Glu平均含量高1.18%,Lys平均含量高0.04%。而GroupⅡ以B032(甘肃茄谷)、B023(山东阴天旱)、B108(甘肃陇谷3号)为代表,其Glu、Arg、Asp、Ala、Gly、Phe、Tyr平均含量与GroupⅢ和GroupⅣ氨基酸含量相比,分别提高了33.7%和67.5%。GroupⅢ以B005(辽宁黑黏谷)为代表,各氨基酸平均含量较GroupⅠ和GroupⅡ降低了59.8%,但较GroupⅣ提高34.6%。GroupⅣ以B089(河北冀谷15)、B021(山东黄黏谷)、B144(山西晋谷23号)等品种为代表,所有氨基酸含量普遍偏低。基于此,不同谷子核心种质的氨基酸含量存在较大差异,通过聚类分析将供试154份谷子种质划分为4个类群,其中GroupⅠ中具有高氨基酸且赖氨酸含量较高的种质,如甘肃陇谷10号和山西向阳谷。

图6 2019(A)、2020年(B)154份谷子种质聚类及各类群氨基酸占比Fig.6 Dendrograms and proportion of amino acids of different groups in 154 foxtail millet germplasms in 2019(A)and 2020(B)
3 结论与讨论
谷子脱粒后即为小米,是日常生活中主要的杂粮之一,具有很高的营养价值,其中氨基酸含量和种类更是丰富多样[9]。作物的种质资源与基因型多样性既是种质创新的物质基础,也是遗传特性研究的重要基石[19],因此,基于人们对膳食营养的追求和谷子优异氨基酸种质资源挖掘的需要,开展关于小米氨基酸组分及其积累特性的研究,筛选优质种质资源及培育新品种就显得尤为重要。
作物遗传变异特性可以从多个角度进行阐释,变异系数就是其中一个重要参数。有研究表明[20],变异系数大于10%即可表示个体间具有较大差异。本研究所测定的不同材料小米氨基酸含量与冯耐红等[15]基本相似,并发现除Asp、Thr、Ala、Ile外,其余13种氨基酸的变异系数均大于10%,表明供试小米种质具有丰富的遗传差异,可为谷子种质资源的遗传基础深入研究提供理论基础。基于2 a的小米各氨基酸含量值,方差分析发现,除His和Val外,其余15种氨基酸都呈极显著差异,其中2019年黑龙江地区的总氨基酸含量最高,达13.87%,表明不同年份产生的生态环境差异对氨基酸的积累有较大影响。有研究表明,气候因素、品种因素、地域因素、播种期等都会显著影响小米氨基酸的积累[21-24]。对不同来源的小米氨基酸含量分析发现,黑龙江、甘肃、山西等地区的氨基酸平均总含量较高,说明这些地区环境选育的品种或品系在遗传特性上更加有利于小米氨基酸的积累。不同夏谷和春谷亚群小米各氨基酸含量有明显差异,本研究中春谷有13种氨基酸含量连续2 a都显著高于夏谷,其中还包含了Lys、Ile等5种必需氨基酸,说明春谷亚群是具有优异品质的种质资源。主要集中在甘肃、山西和黑龙江地区的春谷亚群也表明在一定生境下小米的氨基酸能得到更加有效的积累。
本研究相关性分析结果表明,小米不同氨基酸间均有较高的相关性。Glu与Asp、Gly等13种氨基酸呈极显著正相关,表明其在氨基酸代谢中可能促进其他氨基酸的积累;Phe则表现出与其他氨基酸间的复杂相关性,既有正相关,也有负相关,甚至无相关关系,表明Phe可能参与了各个氨基酸复杂的代谢调控;Lys和Asp间的相关性系数高达0.97,说明Lys和Asp在氨基酸代谢积累中可能存在协同作用,此结果与古世禄等[25]结果相似。
主成分分析能够综合不同氨基酸含量的特征,增加其在优质品种选育中的准确性和可靠性。胡晓航等[26]利用主成分分析对甜菜块根氨基酸进行分析综合评价,鉴定筛选出具有优异营养品质的甜菜品种。本研究通过主成分分析将2019、2020年的小米17种氨基酸分别转化为2个主成分和3个主成分,并根据王倩朝等[27]主成分因子分析筛选标准和莫昭展等[28]综合主成分值模型,得出小米氨基酸综合主成分值,并以此筛选出陇谷10号、陇粟2号、冀谷17、嫩选十四号、白油沙、白毛粱谷、大同黄等氨基酸含量较高的谷子品种或种质。Lys是禾谷类作物中第一限制性必需氨基酸,育种家们也是一直致力于提高禾谷类作物中Lys的水平[29]。在谷子中,任瑞玉等[30]利用多元聚优杂交和水旱穿梭选育相结合的方法选育出高蛋白谷子品种陇谷10号。崔海英等[31]采用有性杂交方法,水旱平行与系谱法相结合连续定向选择筛选出高赖氨酸谷子品种衡谷11号。本研究最终通过聚类分析将154份供试谷子核心种质分为4个类群,并结合主成分分析,从GroupⅠ中筛选出陇粟2号、陇谷5号、陇谷10号、嫩选十一号、大同黄、白毛粱谷、二白谷等高氨基酸品种和陇谷10号、向阳谷等高赖氨酸品种,为谷子优质氨基酸种质筛选和新品种培育提供参考。
小米不同种质间17种氨基酸积累特性存在较大的差异,不同生态环境差异对Glu、Arg等15种氨基酸的积累有较大影响,且春谷小米氨基酸含量显著高于夏谷。各氨基酸之间的积累相互影响,其中Glu仅可正向促进其他氨基酸的积累,而Phe则表现出正负调控,Lys和Asp的积累存在协同作用。结合主成分和聚类分析结果,筛选出了山西白毛粱谷、新疆二白谷等6个高氨基酸品种和高赖氨酸品种。本研究的小米氨基酸组分积累特性分析与评价,可为谷子优质品种的鉴定、筛选以及品质改良提供理论参考。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
中国医药科学(2022年5期)2022-05-05
当代陕西(2021年21期)2022-01-19
四川蚕业(2020年4期)2020-02-10
延河(2018年6期)2018-06-11
中国麻业科学(2018年6期)2018-04-09
中国卫生标准管理(2015年3期)2016-01-14
中国药业(2014年24期)2014-05-26
中医研究(2014年2期)2014-03-11
